蛻膜CXCR6+/-單核-巨噬細胞在人早孕期母胎界面的表型差異
2018-05-09 05:51:41劉倩倩黃煜顏莉莉張鵬張清宇杜美蓉
生殖醫學雜志 2018年5期
關鍵詞:界面
劉倩倩,黃煜*,顏莉莉,張鵬,張清宇,杜美蓉
(1.青島大學附屬青島婦女兒童醫院,青島 266000;2.復旦大學婦產科研究所,上海 200000)
人早孕期母-胎界面主要發生三個生理事件:子宮內膜蛻膜化、滋養細胞發育與胎盤形成、母-胎之間的復雜對話。母胎界面形成中,子宮蛻膜聚攏了大量的免疫細胞,參與母胎免疫耐受的形成,其中蛻膜NK細胞(dNK)含量最多,約占50%~70%,蛻膜T細胞(dT)約占10%~15%,蛻膜單核巨噬細胞(dMΦ)約占10%~15%,幾乎不含有B細胞[1-2]。與dNK細胞不同的是,dMΦ在整個妊娠過程中數量并不會發生明顯的變化[3]。在不同微環境的誘導下單核-巨噬細胞可形成不同的表型,發揮不同的功能。根據表型和功能不同,單核巨噬細胞主要分為兩型即經典M1型巨噬細胞,主要表達共刺激分子CD80和CD86,可分泌TNF-a、IL-8等大量的促炎性細胞因子,影響母胎界面滋養細胞的侵襲,不利于胎盤的植入和妊娠的維持[4-5]。在正常母胎界面偏向于分化為非經典的M2型巨噬細胞,主要表達表面分子CD206,可釋放大量的抗炎細胞因子如IL-10,抑制母體的免疫反應,參與免疫耐受,有利于妊娠的維持[6-8]。
我們課題組前期研究發現人早孕滋養細胞可以分泌高水平CXCL16,通過與其唯一受體CXCR6相互作用選擇性的招募外周單核細胞聚集到蛻膜局部并且保持穩定,從而促進自身的生長和侵襲;然而這些被募集而來的CXCR6+巨噬細胞在蛻膜局部的表型與功能如何,目前仍不明確[9]。因此在前期工作的基礎上,我們采用流式細胞術對正常早孕組和自然流產組婦女CXCR6+/-dMΦ的表型進行分析,揭示其在母胎免疫耐受中可能的作用機制。
材料和方法
一、材料
1.實驗標本: 于上海復旦大學附屬婦產科醫院門診手術室選擇正常早孕期(6~10周)行人工流產的患者40例及早孕期無明確原因自然流產患者20例。
正常早孕組的收集標準:20~35歲計劃外妊娠;術前無感染、發熱等其他基礎疾病。不明原因發生自然流產患者的收集標準:20~35歲,排除染色體異常、生殖道畸形、發熱等其他疾病。
本研究經過倫理委員會批準,取得患者知情同意后,收集蛻膜組織于DMEM/F12培養液中,1 h內帶回實驗室進行細胞分離。
2.實驗試劑:DMEM/F12培養液、PBS、RPMI 1640培養液(Gibco,美國);Percoll分離液,Collagenase Ⅳ(Sigma,美國);DNase I(Applichem,德國);流式抗體(Biolend,美國):APC anti-human CXCR6、FITC anti-human CD14、BV421 anti-human CD45、PE anti-human CXCR6、PE/CY7 anti-human CD80、CD206。
二、實驗方法
1.分離蛻膜免疫細胞群(DIC):將收集到的蛻膜組織置于100 mm的培養皿中,PBS充分洗滌,去除血塊,將組織充分剪碎(約1 mm×1 mm×1 mm),吸入50 ml離心管中,加入0.1 % DNase I及0.1% Collagenase Ⅳ,放入37℃搖床震蕩消化30~40 min。將消化后的組織依次經過100目、300目、400目的濾網過濾,收集濾液,1 500 rpm離心8 min,去上清液,將細胞重懸于2 ml RPMI 1640細胞培養液中,然后平鋪于20%、40%、60% 3個梯度的Percoll分離液上,2 500 rpm離心20 min。40%~60%密度區域的細胞即為分離的蛻膜免疫細胞群,將細胞洗滌并重懸于DMEM/F12培養液中備用。
2.流式細胞術(FCM)檢測dMΦ表面分子的表達:將分離的蛻膜免疫細胞用PBS充分洗滌后,根據樣本的細胞數和抗體推薦用量加入CXCR6、CD45、CD14及表面分子CD80、CD206流式抗體,4℃避光孵育30 min,充分洗滌后加入適量PBS重懸,采用Beckman流式儀(型號CyAn ADP)檢測,Flowjo 7.6.1對結果進行處理分析。
三、統計學處理
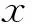
結 果
一、雙色熒光標記分析dMΦ兩組在DIC中的占比
采用CD45、CD14標記dMΦ,首先對蛻膜免疫細胞群“開窗”于CD45+細胞群(圖1A),分析其CD14的標記情況(圖1B)。流式結果分析顯示,不明原因自然流產組CD14+dMΦ在DIC中所占比例為(9.9±1.63)%,正常早孕期組CD14+dMΦ在DIC中所占比例為(8.9±0.88)%,兩組差異并無顯著性(P>0.05)(圖1C)。
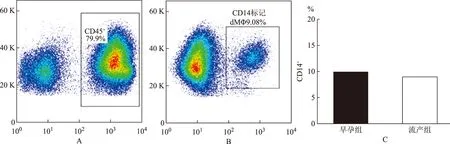
A: CD45標記dMΦ的流式細胞分析散點圖;B: CD14標記dMΦ的流式細胞分析散點圖;C:兩組dMΦ在DIC中占比統計圖圖1 正常早孕組和不明原因自然流產組婦女蛻膜單核-巨噬細胞在蛻膜免疫細胞群中的占比
二、三色熒光標記分析正常早孕組和不明原因自然流產組CXCR6+在dMΦ中的表達情況
圖2是流式細胞分析散點圖。其中雙陽細胞為CD14+CXCR6+的dMΦ,在正常早孕組及不明原因自然流產組中均有表達。正常早孕期組中,CD14+CXCR6+在dMΦ的表達率為(40.0±1.90)%;不明原因自然流產組中,CD14+CXCR6+在dMΦ的表達率為(31.22±2.38)%,兩組比較差異顯著(P<0.05)。
三、多色熒光流式細胞術分析比較兩組蛻膜組織中CD14+CXCR6+/-dMΦ的表型差異
結果顯示:正常早孕組和不明原因自然流產組中,M2型巨噬細胞表面分子CD206在CD14+CXCR6+dMΦ中表達率較CD14+CXCR6-dMΦ均顯著增高(P<0.05),且CD206在正常早孕組CD14+CXCR6+dMΦ和CD14+CXCR6-dMΦ中的表達均顯著高于不明原因自然流產組(P<0.05);正常早孕組中,M1型巨噬細胞表面分子CD80在CD14+CXCR6+dMΦ中表達率顯著高于CD14+CXCR6-dMΦ(P<0.01),而自然流產組中,CD80在CD14+CXCR6+dMΦ和CXCR6-dMΦ的表達率并無顯著性差異(P>0.05)。正常早孕組和不明原因自然流產組相比,CD80在兩組CD14+CXCR6+dMΦ的表達無顯著性差異(P>0.05),在正常早孕組CD14+CXCR6-dMΦ中的表達顯著低于不明原因自然流產組(P<0.01)(圖3)。
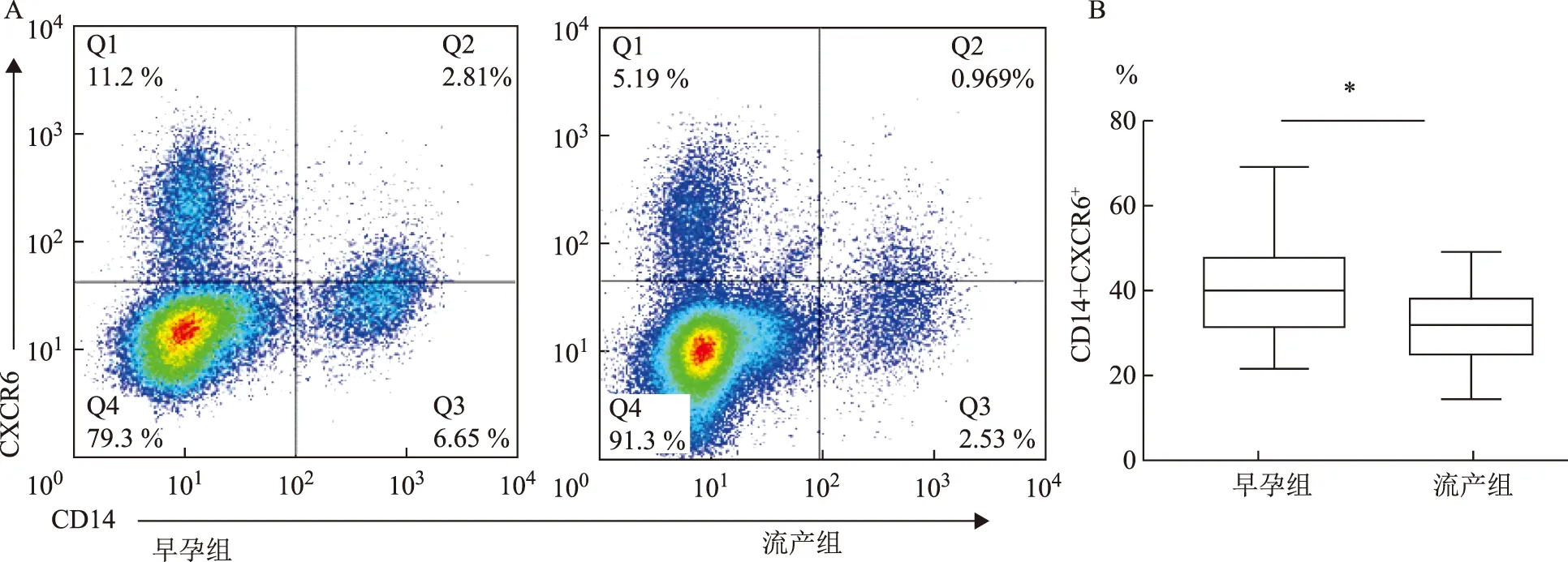
A: 兩組CXCR6+在dMΦ中均有表達流式細胞分析散點圖;B:CD14+CXCR6+在dMΦ的表達率統計結果圖;兩組比較,*P<0.05圖2 流式三色熒光標記分析正常早孕組和不明原因自然流產組CD14+CXCR6+在dMΦ的表達情況

A:正常早孕組和不明原因自然流產組中CD206 在CD14+CXCR6+dMΦ表達率;B:正常早孕組中CD80在CD14+CXCR6+dMΦ表達率;兩組比較,*P<0.05,**P<0.01圖3 正常早孕期與不明原因自然流產組CD14+CXCR6+/-dMΦ的表型比較
討 論
正常妊娠的過程中,母胎界面的細胞免疫即Th1型免疫反應下調,體液免疫即Th2免疫反應占優勢,形成了特殊的免疫耐受環境,對于成功妊娠具有重要作用[10]。單核細胞可從外周遷移到蛻膜局部,分化為巨噬細胞在蛻膜局部大量聚集,通過與滋養細胞和調節性T細胞相互作用,釋放大量的IL-10、IL-8、PGE2等細胞因子參與母胎免疫耐受的建立,調節子宮內膜蛻膜化、胚胎的植入和胎盤發育的全過程[11]。本實驗發現,對比正常早孕組和不明原因自然流產組,dMΦ的數量并沒有明顯的變化,可能由于單核-巨噬細胞免疫功能發生改變,呈現高度易激活狀態,使母體對胎兒產生強烈的排斥反應,最終導致流產的發生。
近年來,人們發現多種趨化因子及其受體是調節免疫應答的重要分子,對 Th1/Th2 型細胞因子的分化、極化及Th1/Th2 型細胞因子的免疫平衡的調節具有重要作用[12]。CXCR6是目前已知趨化因子CXCL16的唯一受體,在本實驗中,相對于正常早孕期組,不明原因自然流產組蛻膜組織中CXCR6+dMΦ在蛻膜的局部的聚集明顯減少,提示CXCR6+dMΦ可能在維持早孕期的母胎界面正常妊娠中發揮重要作用。
普遍認為,蛻膜免疫細胞可通過獨特的表型和分泌大量的細胞因子參與滋養細胞的增殖、侵襲、和分化,從而在母胎界面形成了獨特的免疫耐受微環境[13]。在外周血單核細胞中,80%分化為具有較強抗原提呈功能的經典M1型單核-巨噬細胞,而在正常早孕期母胎界面蛻膜免疫細胞中單核-巨噬細胞在可溶性HLA-G5及細胞因子IL-4、IL-13、IL-10和M-CSF的誘導下更趨向于分化為具有免疫抑制作用的非經典M2型巨噬細胞[3]。有研究發現,在某些腫瘤微缺氧環境中,CXCR6的表達水平與組織局部M2型單核細胞的表達呈正相關,CXCL16/CXCR6可介導單核-巨噬細胞在腫瘤組織中的活性,并且使其更趨向于分化為具有免疫抑制作用的M2型巨噬細胞,參與腫瘤的血管生成和轉移[14]。在正常早孕期母胎界面也存在著類似于腫瘤的缺氧微環境,影響著滋養細胞的侵襲和子宮螺旋動脈的生成[15]。我們通過實驗發現 M2型巨噬細胞表面標志分子CD206在正常早孕期和不明原因自然流產婦女CXCR6+dMΦ中的表達都要顯著高于CXCR6-dMΦ,這提示在早孕期母胎界面CXCR6可介導單核-巨噬細胞趨向于分化為M2型巨噬細胞。同時實驗還發現M1型巨噬細胞表面分子CD80在正常早孕期婦女CXCR6+dMΦ中的表達高于CXCR6-dMΦ,而在不明原因自然流產婦女CXCR6+/-dMΦ的表達水平無明顯差異,提示CXCR6與母胎界面促炎性M1型巨噬細胞在不明原因自然流產中的高度聚集相關。
由此可見,CXCR6不僅參與募集和吸引母體外周血免疫細胞到達并停留于妊娠子宮,還可能對局部的免疫細胞產生誘導效應,維持M1/M2型巨噬細胞達到一定比例的動態平衡。我們實驗還發現,在不明原因自然流產婦女CXCR6+dMΦ中,M2型巨噬細胞比例降低,M1型巨噬細胞在CXCR6-dMΦ中的比例升高,更驗證了CXCR6可能參與了早孕期M2型巨噬細胞在蛻膜組織中的形成和分化及蛻膜局部TH2型免疫偏倚的形成,當不明原因自然流產反復發生時,CXCR6的表型和功能特性也會發生改變,從而改變母胎界面的免疫微環境,打破免疫耐受的平衡狀態。
總之,在不明原因自然流產中,蛻膜單核-巨噬細胞數量比例并未發生明顯變化,CXCR6可能通過介導單核-巨噬細胞的分化,影響母胎界面免疫耐受,此發現為預防和治療復發性流產提供了新的思路,但其相關功能特性,影響機制和通路還需進一步的實驗研究。
【參考文獻】
[1] Xu YY,Wang SC,Li DJ,et al. Co-signaling molecules in maternal-fetal immunity[J]. Trends Mol Med,2017,23:46-58.
[2] 黃煜,李大金,朱影.三色熒光標記鑒定早孕蛻膜及外周免疫細胞組成[J].生殖醫學雜志,2006,15: 328-332.
[3] Tsai YC,Tseng JT,Wang CY,et al. Medroxyprogesterone acetate drives M2 macrophage differentiation toward a phenotype of decidual macrophage[J]. Mol Cell Endocrinol,2017,452:74-83.
[4] Ning F,Liu H,Lash GE. The role of decidual macrophages during normal and pathological pregnancy[J]. Am J Reprod Immunol,2016,75:298-309.
[5] 唐茂興,廖愛華.單核細胞亞群與妊娠[J].中國免疫學雜志,2014,30:717-721.
[6] Houser BL,Tilburgs T,Hill J,et al. Two unique human decidual macrophage populations[J]. J Immunol,2011,186:2633-2642.
[7] Kalish SV,Lyamina SV,Usanova EA,et al. Macrophages reprogrammed in vitro towards the M1 phenotype and activated with LPS extend lifespan of mice with ehrlich ascites carcinoma[J]. Med Sci Monit Basic Res,2015,21:226-234.
[8] Yamaguchi T,Fushida S,Yamamoto Y,et al. Tumor-associated macrophages of the M2 phenotype contribute to progression in gastric cancer with peritoneal dissemination[J]. Gastric Cancer,2016,19:1052-1065.
[9] Huang Y,Zhu XY,Du MR,et al. Human trophoblasts recruited T lymphocytes and monocytes into decidua by secretion of chemokine CXCL16 and interaction with CXCR6 in the first-trimester pregnancy[J]. J Immunol,2008,180:2367-2375.
[10] Piao HL,Tao Y,Zhu R,et al. The CXCL12/CXCR4 axis is involved in the maintenance of Th2 bias at the maternal/fetal interface in early human pregnancy[J]. Cell Mol Immunol,2012,9:423-430.
[11] Svensson-Arvelund J,Mehta RB,Lindau R,et al. The human fetal placenta promotes tolerance against the semiallogeneic fetus by inducing regulatory T cells and homeostatic M2 macrophages[J]. J Immunol,2015,194:1534-1544.
[12] 于潔,黃煜,崔竹梅,等.趨化因子CXCL16對人早孕蛻膜免疫細胞的遷移及分泌細胞因子的調節作用[J].現代免疫學,2009,3:235-239.
[13] 朱曉勇,李大金.母胎免疫調節機理的研究進展[J].生殖醫學雜志,2000,9: 56-60.
[14] Cho SW,Kim YA,Sun HJ,et al. CXCL16 signaling mediated macrophage effects on tumor invasion of papillary thyroid carcinoma[J]. Endocr Relat Cancer,2016,23:113-124.
[15] 羅健英,喬福元.缺氧對細胞滋養細胞MMP9/TIMP1基因表達及細胞侵襲力的影響[J].華中科技大學學報(醫學版),2010,39:216-220.
猜你喜歡
艦船科學技術(2022年16期)2022-09-22 02:15:00
北京航空航天大學學報(2021年6期)2021-07-20 07:23:54
當代陜西(2020年13期)2020-08-24 08:22:02
制造技術與機床(2017年5期)2018-01-19 02:49:17
制造技術與機床(2017年11期)2017-12-18 06:47:29
金秋(2017年4期)2017-06-07 08:22:16
蘇州科技大學學報(自然科學版)(2017年1期)2017-03-20 15:25:18
中國材料進展(2016年10期)2016-12-26 06:50:20
濰坊學院學報(2016年2期)2016-12-01 13:00:11
新聞傳播(2015年11期)2015-07-18 11:15:04
