茶樹CsLhcb4基因的電子克隆與生物信息學(xué)分析
2018-05-30 04:52:50梅菊芬徐德良湯茶琴周靜峰邵元海
江蘇農(nóng)業(yè)科學(xué) 2018年9期
關(guān)鍵詞:結(jié)構(gòu)分析
梅菊芬, 徐德良, 湯茶琴, 周靜峰, 邵元海
(無錫市茶葉品種研究所/江蘇省茶樹種質(zhì)改良與推廣工程技術(shù)中心,江蘇無錫 214125)
光合作用產(chǎn)物是人類賴以生存和發(fā)展的基礎(chǔ),光捕獲是植物光合作用的重要過程。在高等植物中,捕光葉綠素a/b結(jié)合蛋白(light harvesting chlorophyll a/b binding protein,LHC)是光捕獲中的重要功能蛋白,其與葉綠素a/b形成復(fù)合體,將光能迅速傳到光系統(tǒng)Ⅰ(PSⅠ)和光系統(tǒng)Ⅱ(PSⅡ)的反應(yīng)中心,使光能轉(zhuǎn)化為化學(xué)能,促進(jìn)光合反應(yīng)的進(jìn)行[1]。PSⅠ和PSⅡ都含有各自的LHC,即LHCⅠ和LHCⅡ。其中,LHCⅡ在類囊體膜上的含量最為豐富,它所結(jié)合的葉綠素約占類囊體膜上色素量的50%,目前研究較多,也是結(jié)構(gòu)較清楚的一類膜蛋白[1-3]。LHCⅡ被認(rèn)為是一類結(jié)構(gòu)相似、進(jìn)化相關(guān)、由核基因家族(Lhcb)編碼的蛋白,與色素所形成的色素蛋白復(fù)合體家族,含有保守的葉綠素結(jié)合(chlorophyllbinding,CB)結(jié)構(gòu)域,它們除進(jìn)行光能的捕獲與傳遞外,還廣泛參與了激發(fā)能在PSⅠ和PSⅡ之間的調(diào)節(jié)與分配、類囊體膜結(jié)構(gòu)的維持、光保護(hù)以及對各種環(huán)境的應(yīng)答等[4]。LHCⅡ主要由6種色素蛋白復(fù)合體組成,分別由Lhcb1、Lhcb2、Lhcb3、Lhcb4、Lhcb5和Lhcb6編碼,其組成了4種蛋白亞復(fù)合物,LHCⅡa、LHCⅡb、LHCⅡc、LHCⅡd。其中,LHCⅡb為主體LHCⅡ,包括LHCB1、LHCB2、LHCB3;其他3種合稱為微量LHCⅡ,分別為LHCⅡa(LHCB4)、LHCⅡc(LHCB5)、LHCⅡd(LHCB6)[1]。
LHCB4別稱CP29,是PSⅡ內(nèi)部天線葉綠素a/b結(jié)合亞復(fù)合體,其為次要捕光色素蛋白,較其他蛋白復(fù)合體更接近PSⅡ核心復(fù)合物,其編碼基因?yàn)長hcb4[1]。目前已從擬南芥、秈稻、楊樹、菠菜、銀杏等多種高等植物中克隆了Lhcb4基因,并研究了其在各種生態(tài)環(huán)境下的表達(dá)情況[5-10]。
近年,茶樹育種目標(biāo)傾向葉色多樣化,白化或紫化等特異葉色品種的選育成為茶樹新品種選育的熱點(diǎn)。研究表明,光是誘導(dǎo)茶樹花青素形成的主要原因[11],光合作用與花青素的合成之間是否存在一定關(guān)系有待于研究。本研究從紫芽中篩選到下調(diào)表達(dá)基因片段,Blastx比對為Lhcb4的部分基因片段。以該基因片段為探針,在NCBI中Blast檢索茶樹中與探針序列相似性較高的ESTs,然后利用DNAstar進(jìn)行序列拼接,得到茶樹葉綠素a/b結(jié)合蛋白CP29基因(CsLhcb4),利用生物信息學(xué)技術(shù)預(yù)測該基因編碼蛋白的理化性質(zhì)和結(jié)構(gòu)分析,為后續(xù)研究CsLhcb4與花青素代謝的關(guān)系提供參考。
1 材料與方法
1.1 材料
筆者所在研究組從紫芽中篩選的差異表達(dá)基因片段(D-ZY-5),經(jīng)Blastx比對為Lhcb4的部分基因片段。
1.2 方法
1.2.1 茶樹CsLhcb4基因的電子克隆 以差異篩選的EST基因片段為查詢探針,利用NCBI中的Blastn工具,搜索茶樹EST數(shù)據(jù)庫(taxid:4442),得到具有高度同源性(選擇相似性大于等于90%,覆蓋率大于100 bp)的EST序列,再利用DNAStar進(jìn)行重疊區(qū)域拼接和組裝,構(gòu)建重疊序列群。以重疊序列群為信息標(biāo)簽,進(jìn)一步檢索比對,搜索高度同源序列,并拼接組裝,直至沒有發(fā)現(xiàn)高度同源序列。在非冗余數(shù)據(jù)庫中進(jìn)行搜索,確認(rèn)新基因序列,獲得茶樹CsLhcb4的cDNA序列。
1.2.2 茶樹CsLhcb4基因及蛋白質(zhì)序列的生物信息學(xué)鑒定 以NCBI的ORFfinder和Conserved Domain Database功能對擴(kuò)增的基因序列進(jìn)行蛋白質(zhì)翻譯及其保守結(jié)構(gòu)域預(yù)測。以Expasy的在線分析軟件對蛋白序列進(jìn)行理化性質(zhì)(ProtParam tool)、疏水性(ProtScale)、二級結(jié)構(gòu)(SOPMA)和三維結(jié)構(gòu)分析預(yù)測(SWISS-MODEL),同時(shí)利用在線分析工具分析無序區(qū)域(foldindex)、亞細(xì)胞定位(TargetP 1.1 Server)及磷酸化位點(diǎn)預(yù)測(NetPhos 3.1 Server)。利用NCBI的PSI-BLAST進(jìn)行蛋白序列同源比對,利用DNAman軟件進(jìn)行同源蛋白的氨基酸多序列比對并用Mega構(gòu)建系統(tǒng)進(jìn)化樹。
2 結(jié)果與分析
2.1 茶樹CsLhcb4基因克隆及序列分析
以篩選的EST基因片段為查詢探針,利用Blastn對茶樹EST數(shù)據(jù)庫進(jìn)行搜索,得到2個(gè)高度同源的EST序列(FS954792.1和JK993206.1),利用DNAStar對序列進(jìn)行重疊區(qū)域拼接和組裝,在以拼接的重疊序列群進(jìn)行檢索,得到高度同源的EST序列(FS960161.1),重復(fù)拼接得到茶樹CsLhcb4基因cDNA序列,長度為1 085 bp。經(jīng)ORFfinder分析,該基因包含1個(gè)858 bp的ORF,編碼1個(gè)含285個(gè)氨基酸的蛋白質(zhì)。
2.2 茶樹CP29蛋白保守結(jié)構(gòu)域預(yù)測
利用NCBI在線分析軟件Conserved Domain Database(CDD)對蛋白進(jìn)行結(jié)構(gòu)域分析,結(jié)果(圖1)顯示,在茶樹葉綠素a/b連接蛋白CP29的氨基酸序列第13個(gè)和第285個(gè)之間存在光系統(tǒng)Ⅱ光捕獲復(fù)合體結(jié)構(gòu)域(PLN00187),編碼蛋白屬于葉綠素a/b連接蛋白超家族成員。

2.3 茶樹CP29蛋白同源比對及序列進(jìn)化分析
在NCBI中的Blastp中采用PSI-Blast方法在refseq數(shù)據(jù)庫中進(jìn)行氨基酸序列同源比對,結(jié)果顯示,有478個(gè)蛋白與茶樹CP29蛋白同源,一致性最大為89%,最小為32%,這些蛋白包括假定未知的蛋白、預(yù)測的蛋白、功能已鑒定的蛋白,它們都有葉綠素a/b結(jié)合蛋白超家族結(jié)構(gòu)域(PLN00187),相對保守。在功能已鑒定的蛋白中,茶樹CP29蛋白與陸地棉CP29.1(NP001314598.1)、歐洲大葉楊CP29(XP002323575.1)、大豆CP29(NP001304343.1)、擬南芥(NP187506.1)、深山南芥(XP002873005.1)、玉米(NP001105502.1)、衣藻(XP001697193.1)、綠藻(XP005646225.1)一致性分別是87%、87%、87%、83%、83%、82%、51%、47%,這表明植物葉綠素a/b連接蛋白CP29序列在不同植物之間具有一定程度的序列分化。利用DNAman軟件將茶樹與這8個(gè)植物的CP29蛋白進(jìn)行多重比較分析,序列一致性為74.19%(圖2)。Mega構(gòu)建系統(tǒng)發(fā)育樹表明,高等植物的CP29區(qū)別于藻類,在進(jìn)化距離上較近;茶樹與大豆、歐洲大葉楊和陸地棉進(jìn)化距離最近,聚為一類(圖3)。

2.4 茶樹CP29蛋白理化性質(zhì)分析
CsLhcb4編碼的蛋白質(zhì)分子式為C1 427H2 180N366O407S4,分子量為31.1 ku,理論等電位點(diǎn)5.79。亮氨酸(Leu)和丙氨酸(Ala)占比最大,分別為11.2%和10.9%,芳香族氨基酸[苯丙氨酸(Phe)、酪氨酸(Tyr)、色氨酸(Trp)]33個(gè),占比11.6%。負(fù)電荷殘基[天冬氨酸(Asp)+谷氨酸(Glu)]數(shù)為31個(gè),正電荷殘基[精氨酸(Arg)+賴氨酸(Lys)]數(shù)為27個(gè),脂溶系數(shù)為84,總平均疏水性為-0.099。不穩(wěn)定系數(shù)31.69,該蛋白為穩(wěn)定蛋白。無序區(qū)域預(yù)測結(jié)果表明,其存在1個(gè)無序區(qū)域1~50位氨基酸處,不可折疊性0.194,對蛋白折疊、表達(dá)水平干擾較小。

2.5 茶樹CP29蛋白疏水性分析
用ProtScale軟件對蛋白質(zhì)進(jìn)行疏水性/親水性分析(圖4),正值越大表示越疏水,負(fù)值越大表示越親水,介于+0.5~-0.5之間的主要為兩性氨基酸。結(jié)果表明,Glu191疏水性最強(qiáng),疏水性參數(shù)為2.789;Leu206和Asp207親水性最強(qiáng),疏水性參數(shù)為-2.333。蛋白中約在183~197區(qū)域疏水性最強(qiáng),其次是245~256、144~157區(qū)域具有較強(qiáng)的疏水性;在199~219區(qū)域親水性最強(qiáng),其次在38~54、227~232區(qū)域具有較強(qiáng)的親水性。該蛋白的親水區(qū)域大于疏水區(qū)域,預(yù)測該蛋白為親水蛋白,與ProtParam分析結(jié)果一致。

2.6 茶樹CP29蛋白亞細(xì)胞定位預(yù)測
結(jié)合TargetP亞細(xì)胞定位預(yù)測(表1),蛋白定位于葉綠體。CP29為植物PSⅡ3種次要捕光色素蛋白之一,蛋白亞細(xì)胞定位結(jié)果符合其功能預(yù)測。

表1 茶樹CP29蛋白亞細(xì)胞定位
2.7 茶樹CP29蛋白磷酸化修飾位點(diǎn)及分析
磷酸化是蛋白翻譯前修飾的最常見的形式之一,在調(diào)節(jié)從基因表達(dá)到信號和代謝調(diào)控等所有細(xì)胞功能中都起作用[4]。CP29的磷酸化可能與植物的抗逆有關(guān)[12-13],其磷酸化可能是調(diào)節(jié)光能的捕獲及激發(fā)能向反應(yīng)中心傳遞的一種方式,也是一種新的光保護(hù)機(jī)制。利用NetPhos 3.1預(yù)測茶樹CP29蛋白磷酸化位點(diǎn),結(jié)果(表2)顯示,CP29蛋白磷酸化位點(diǎn)有23個(gè),其中Thr 9個(gè),絲氨酸(Ser)10個(gè),Tyr 4個(gè)。
2.8 茶樹CP29蛋白二級結(jié)構(gòu)預(yù)測和分析
利用SOPMA在線預(yù)測蛋白的二級結(jié)構(gòu)(圖5),α-螺旋和無規(guī)則卷曲是其主要二級結(jié)構(gòu),其中115個(gè)氨基酸可能形成無規(guī)則卷曲,占40.35%,97個(gè)氨基酸可能形成α-螺旋,占34.04%,49個(gè)氨基酸可能形成延伸鏈,占17.19%,24個(gè)氨基酸可能形成β-轉(zhuǎn)角,占8.42%,不含其他二級結(jié)構(gòu)。
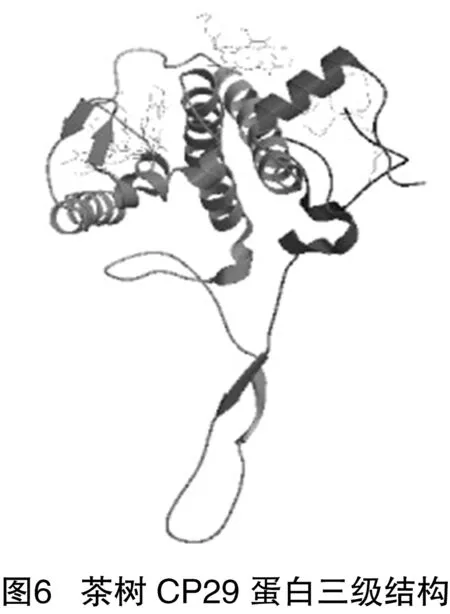
2.9 茶樹CP29蛋白三級結(jié)構(gòu)建模及分析
利用swiss-model在線對其進(jìn)行建模,從PDB(蛋白質(zhì)結(jié)構(gòu)數(shù)據(jù)庫)中獲得63個(gè)同源蛋白序列,選擇其中的3jcu.1菠菜PSⅡ-LHCⅡ超分子復(fù)合物,與靶標(biāo)序列比對結(jié)果作模板,其序列一致性為88.51%,獲得其三級結(jié)構(gòu)(圖6)。三級結(jié)構(gòu)從N端開始形成8端螺旋結(jié)構(gòu),至C端形成較長的延伸鏈。建模結(jié)果估值(OMQE)為0.75,介于0~1之間,接近1,建模結(jié)果處于接受水平。但是QMEAN4值為-3.95,其理想值為1,因此建模結(jié)果并不十分理想。
3 討論與結(jié)論
LHC蛋白是高等植物光合作用過程中的重要功能蛋白,本研究以在紫芽中下調(diào)表達(dá)的基因片段(D-ZY-5)為查詢探針,通過電子克隆的方式得到編碼茶樹CP29蛋白基因的cDNA序列,命名為CsLhcb4,長度為1 085 bp,ORF為 858 bp,編碼285個(gè)氨基酸,分子量為31.1 ku。蛋白保守結(jié)構(gòu)域分析表明其編碼氨基酸序列第13個(gè)和第285個(gè)之間存在光系統(tǒng)Ⅱ光捕獲復(fù)合體結(jié)構(gòu)域(編號PLN00187),屬于葉綠素a/b連接蛋白超家族成員。亞細(xì)胞定位于葉綠體上,與蛋白預(yù)測的功能相吻合。
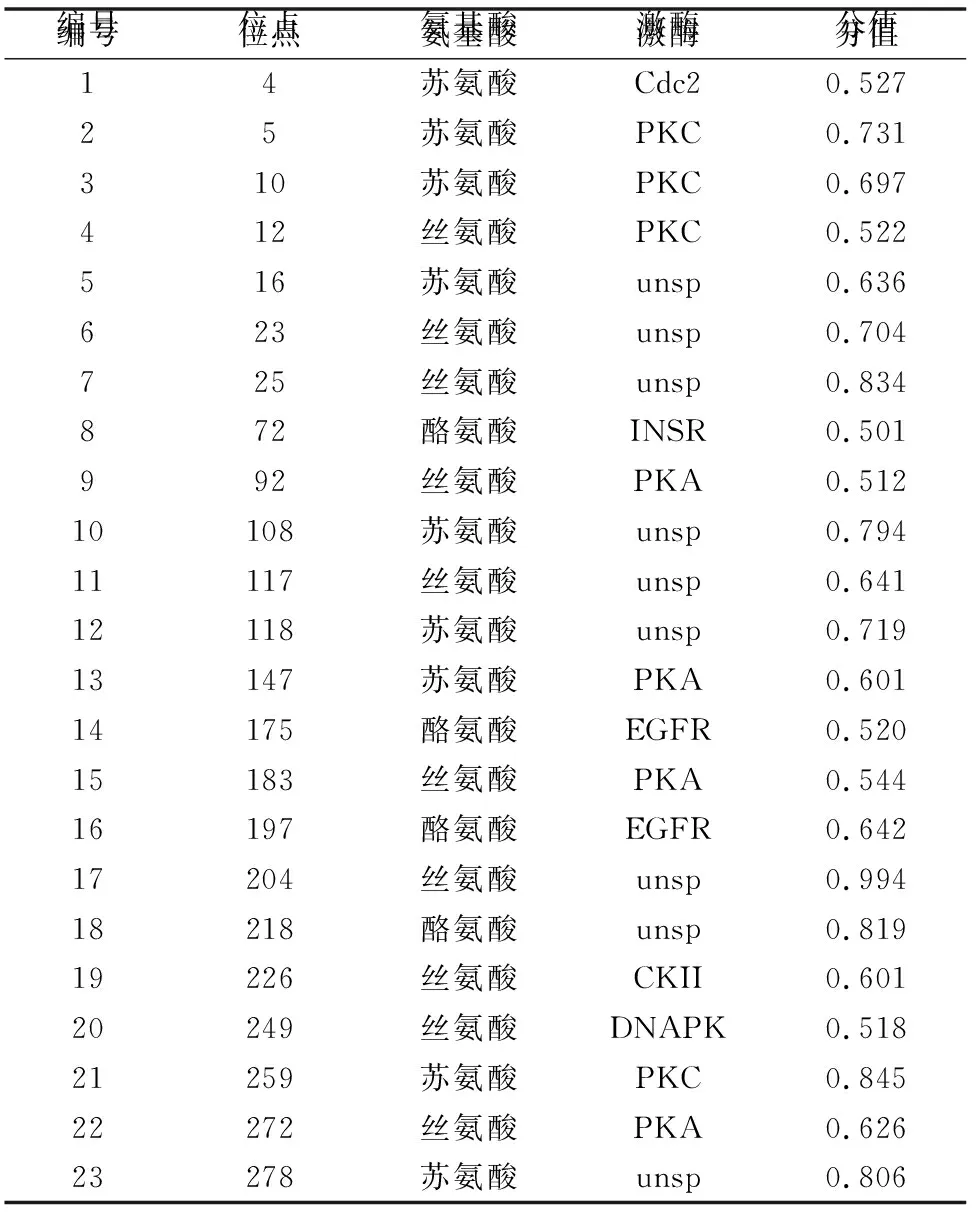
表2 茶樹CP29蛋白磷酸化修飾位點(diǎn)預(yù)測

氨基酸同源序列分析表明,克隆的茶樹CsLhcb4編碼氨基酸與這些序列相似性為32%~89%,總體上相對保守,屬于葉綠素a/b結(jié)合蛋白CP29。構(gòu)建進(jìn)化樹表明,茶樹與高等植物陸地棉、綠豆和煙草進(jìn)化距離最近,聚為一類。
二級結(jié)構(gòu)分析表明,α-螺旋和無規(guī)則卷曲是茶樹CP29的主要二級結(jié)構(gòu),α-螺旋是一種穩(wěn)定結(jié)構(gòu),能維持蛋白的結(jié)構(gòu)穩(wěn)定性。三維結(jié)構(gòu)分析表明,基于茶樹CP29與菠菜PSⅡ-LHCⅡ超分子復(fù)合物(3jcu.1)序列同源性為88.51%,利用Swiss-Model中的同源建模法構(gòu)建茶樹CP29的三維結(jié)構(gòu),構(gòu)建模型為單體形式。
氨基酸序列親水性/疏水性分析結(jié)果顯示,該蛋白的親水區(qū)域大于疏水區(qū)域,預(yù)測該蛋白為親水蛋白。不穩(wěn)定系數(shù)為31.69,該蛋白為穩(wěn)定蛋白,能穩(wěn)定保持其與葉綠素a/b形成的復(fù)合體,傳輸光能,促進(jìn)光合作用。
在高等植物中,CP29在LHCⅡ中分子量最大,結(jié)構(gòu)最簡單,光譜值相對簡單,結(jié)合6個(gè)葉綠素a、2個(gè)葉綠素b和2個(gè)類胡蘿卜素分子,其結(jié)構(gòu)及功能研究相對明確[14-15]。研究表明,CP29是植物光系統(tǒng)Ⅱ天線蛋白不可或缺的結(jié)構(gòu)組成,在非光化學(xué)淬滅(NPQ)作用中至關(guān)重要,其缺失會造成光系統(tǒng)Ⅱ結(jié)構(gòu)變化,并影響光保護(hù)作用[16-18]。CP29的磷酸化可能與植物的抗逆性有一定關(guān)系,低溫、強(qiáng)光等逆境能誘導(dǎo)CP29的磷酸化,其磷酸化可在多個(gè)位點(diǎn)發(fā)生,并通過可逆磷酸化調(diào)節(jié)和決定LHCⅡ在2個(gè)光系統(tǒng)之間的親和性和LHCⅡ的狀態(tài)遷移[19]。
本研究通過電子克隆方式得到在高花青素茶樹中下調(diào)表達(dá)的葉綠素a/b結(jié)合蛋白CP29基因,通過生物信息學(xué)分析預(yù)測其理化性質(zhì)、二級結(jié)構(gòu)和三維結(jié)構(gòu),為進(jìn)一步研究CP29在高花青素茶樹代謝中的作用奠定了基礎(chǔ)。CP29是否通過光保護(hù)作用或磷酸化等來影響花青素的代謝有待于進(jìn)一步研究。
捕光色素蛋白有14類,最初序列上的相似性卻表明這些色素蛋白都起始于1個(gè)相同的原始基因,原始基因能夠編碼1個(gè)含跨膜螺旋的蛋白[4]。高等植物的PSⅡ外周光捕獲蛋白都在非光化學(xué)淬滅(NPQ)下發(fā)揮其作用,起到光保護(hù)作用[17]。因此,在CP29的研究基礎(chǔ)上繼續(xù)對高花青素茶樹捕光色素蛋白的表達(dá)及功能進(jìn)行研究具有一定的意義。
:
[1]孫欽秒,冷 靜,李良璧,等. 高等植物光系統(tǒng)Ⅱ捕光色素蛋白復(fù)合體結(jié)構(gòu)與功能研究的新進(jìn)展[J]. 植物學(xué)通報(bào),2000,17(4):289.
[2]Labate M T,Ko K,Ko Z W,et al. Constitutive expression of peaLhcb1-2 in tobacco affects plant development,morphology and photosynthetic capacity[J]. Plant Molecular Biology,2004,55(5):701-714.
[3]李曉鵬. 杜林方和梁厚果.菠菜光系統(tǒng)Ⅱ天線組分CP29的分離及其性質(zhì)[J]. 生物化學(xué)與生物物理學(xué)報(bào),1999,31(6):631-636.
[4]羅 玲. 捕光色素蛋白復(fù)合物的研究進(jìn)展[J]. 現(xiàn)代農(nóng)業(yè)科技,2008(22):270-273,276.
[5]Xu Y H,Liu R,Yan L,et al. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid inArabidopsis[J]. Journal of Experimental Botany,2012,63(3):1095-1106.
[6]袁定陽,余 東,譚炎寧,等. RT-PCR克隆秈稻葉綠素a/b結(jié)合蛋白基因全長cDNA及序列的in silico分析[J]. 基因組學(xué)與應(yīng)用生物學(xué),2012,31(2):173-177.
[7]王歡利,劉新亮,郁萬文,等. 銀杏葉綠素a/b結(jié)合蛋白基因(GbLhcb4)及其啟動(dòng)子克隆[J]. 中南林業(yè)科技大學(xué)學(xué)報(bào),2015,35(5):114-121.
[8]Klimmek F,Sj?din A,Noutsos C,et al. Abundantly and rarely expressed Lhc protein genes exhibit distinct regulation patterns in plants[J]. Plant Physiology,2006,140(3):793-804.
[9]Pan X,Li M,Wan T,et al. Structural insights into energy regulation of light-harvesting complex CP29 from spinach[J]. Nature Structural & Molecular Biology,2011,18(3):309-315.
[10]鄒 智,安 鋒,楊禮富,等. 大戟科Lhcb基因家族的全基因組鑒定、分類與進(jìn)化分析[J]. 中南林業(yè)科技大學(xué)學(xué)報(bào),2013,33(12):46-52.
[11]張澤岑,王能彬. 光質(zhì)對茶樹花青素含量的影響[J]. 四川農(nóng)業(yè)大學(xué)學(xué)報(bào),2002,20(4):337-339.
[12]劉文娟,袁 澎,林宏輝. 高等植物的光系統(tǒng)Ⅱ蛋白磷酸化機(jī)制及其對環(huán)境脅迫的響應(yīng)[J]. 植物生理學(xué)通訊,2007,43(6):995-1001.
[13]Liu W J,Chen Y E,Tian W J,et al. Dephosphorylation of photosystem Ⅱ proteins and phosphorylation of CP29 in barley photosynthetic membranes as a response to water stress[J]. Biochimica et Biophysica acta,2009,1787(10):1238-1245.
[14]Pascal A,Peterman E,Gradinaru C,et al. Structure and interactions of the chlorophyll a molecules in the higher plantLhcb4 antenna protein[J]. Journal of Physical Chemistry B,2000,104(39):9317-9321.
[15]Gastaldelli M,Canino G,Croce R,et al. Xanthophyll binding sites of the CP29 (Lhcb4) subunit of higher plant photosystem Ⅱ investigated by domain swapping and mutation analysis[J]. Journal of Biological Chemistry,2003,278(21):19190-19198.
[16]de Bianchi S,Betterle N,Kouril R,et al.Arabidopsismutants deleted in the light-harvesting proteinLhcb4 have a disrupted photosystem Ⅱ macrostructure and are defective in photoprotection[J]. The Plant Cell,2011,23(7):2659-2679.
[17]Mozzo M,Passarini F,Bassi R,et al. Photoprotection in higher plants:the putative quenching site is conserved in all outer light-harvesting complexes of photosystem Ⅱ[J]. Biochimica et Biophysica Acta,2008,1777(10):1263-1267.
[18]Miloslavina Y,de Bianchi S,Dall’osto L,et al. Quenching inArabidopsisthalianamutants lacking monomeric antenna proteins of photosystem Ⅱ[J]. The Journal of Biological Chemistry,2011,286(42):36830-36840.
[19]Kargul J,Turkina M V,Nield J,et al. Light-harvesting complex Ⅱ protein CP29 binds to photosystem Ⅰ ofChlamydomonasreinhardtiiunder state 2 conditions[J]. FEBS Journal,2005,272(18):4797-4806.
猜你喜歡
哲學(xué)評論(2021年2期)2021-08-22 01:53:34
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- 基于因子分析法的農(nóng)村電子商務(wù)發(fā)展水平實(shí)證分析
- 我國農(nóng)村人力資源開發(fā)研究的文獻(xiàn)計(jì)量分析
- 我國區(qū)域財(cái)政農(nóng)業(yè)支出非均衡性與優(yōu)化
- 中國與絲綢之路經(jīng)濟(jì)帶主要國家農(nóng)產(chǎn)品貿(mào)易增長分解及影響因素
——基于CMS模型與拓展引力模型的實(shí)證分析 - PSR框架下生態(tài)文明建設(shè)區(qū)域差異分析
——以江蘇省為例 - 農(nóng)業(yè)產(chǎn)業(yè)鏈融資與農(nóng)戶收入