KLF5在脫氧膽酸誘導胃黏膜腸上皮化生中的作用*
2018-07-13 14:12:58孫杰付立芳
中國現代醫學雜志 2018年20期
關鍵詞:差異
孫杰,付立芳
(1.山東醫學高等專科學校 中醫學教研室 山東 臨沂 276000;2.山東省臨沂市中醫醫院 呼吸內科,山東 臨沂 276002)
胃癌的發生、發展是一個漸進過程,從正常胃黏膜、腸上皮化生、異型增生向胃癌的發生、發展進程中有多種基因的參與[1]。含有膽汁酸的胃十二指腸反流引起的胃黏膜慢性炎癥,進而發生胃黏膜腸上皮化生[2]。目前有研究表明,Krüppel樣因子5(Krüppellike factor 5,KLF5)在慢性胃炎和腸上皮化生的表達與正常對照組相比,呈上升趨勢[3-4]。因此,本文通過脫氧膽酸(deoxycholic acid,DCA)誘導胃黏膜上皮細胞,探究KLF5是否參與DCA誘導的腸上皮化生的發生及其可能的作用機制。
1 材料與方法
1.1 材料與試劑
胃黏膜上皮細胞GES-1(中國科學院上海細胞庫)。選取山東省臨沂市中醫醫院2012~2015年的56例胃黏膜標本,其中30例正常胃黏膜,26例腸上皮化生。本研究經本院醫學倫理委員會批準,并與患者簽署知情同意書。鼠抗KLF5、TFF2、VIL1、CDX2、Wnt3a、β-catenin、多克隆抗體、β-actin鼠抗單克隆抗體、羊抗鼠二抗(HRP)購自英國Abcam公司,MTT、二甲基亞砜、二喹啉甲酸(bicinchoninic acid,BCA)試劑盒、Annexin V-FITC/PI凋亡試劑盒購自上海碧云天生物技術有限公司,流式細胞儀(美國BD公司)。
1.2 方法
1.2.1正常胃黏膜和腸上皮化生組織的選取正常胃黏膜30例,其中,男性13例,女性17例;腸上皮化生26例,其中,男性16例,女性10例。以上組織均置于-80℃中保存,用于提取蛋白和mRNA。
1.2.2胃黏膜上皮細胞的培養和轉染GES-1細胞在含10%胎牛血清、100μ/ml青霉素、100μg/ml鏈霉素的RPMI 1640培養基中培養,置于37℃、5%二氧化碳CO2孵箱中生長。將細胞分為兩組,應用1 mol/L氯化氫調定培養基pH值至7.2,采用不同濃度(100、200和500μmol/L)的DCA處理細胞12 h,置于37℃、5% CO2孵箱中孵育。空白對照組為不含DCA的RPMI 1640培養液,繼續培養12 h。
1.2.3細胞轉染將GES-1細胞分為空白對照組、DCA組、陰性對照組及KLF5沉默組。細胞長至50%~60%密度時,分別將KLF5 siRNA和陰性序列對照按50 nmol/L濃度轉染GES-1細胞,24 h后更換為不含雙抗的RPMI 1640培養液,并加入500μmol/L DCA或者溶劑對照(Veh)處理細胞,置于37℃、5%CO2培養箱中繼續培養24 h,觀察細胞感染情況。用Western blot檢測驗證KLF5沉默的效率。
1.2.4MTT法MTT法檢測細胞增殖。按分組收集轉染48 h后的GES-1細胞,磷酸鹽緩沖溶液(phosphate buffer saline,PBS)清洗后加入MTT,37℃溫育4 h,加入二甲基亞砜150μl/孔,將培養板置于微孔板振蕩器上振蕩10 min,使結晶物溶解,用酶標儀在570 nm處測吸光度值。
1.2.5流式細胞術按分組收集轉染48 h后的GES-1細胞,PBS清洗,參照Annexin V-FITC/PI凋亡檢測試劑盒說明書,在流式細胞儀上檢測細胞凋亡。
1.2.6Western blot檢測分別取正常胃黏膜、腸上皮化生組織,以及DCA處理GES-1細胞樣品,提取總蛋白。采用BCA法測定蛋白濃度,十二烷基磺酸鈉-聚丙烯酰胺凝膠電泳、轉膜,5%脫脂奶粉封閉,4℃條件下用抗KLF5、Wnt3a和β-actin(均為1∶300)的一抗孵育過夜,PBST洗膜3次,二抗室溫孵育1 h,PBST洗膜3次,最后用增強化學發光法發光液在暗室曝光并壓片。將GES-1細胞分為空白對照組、DCA組、陰性對照組及KLF5沉默組,按分組收集轉染48 h后的GES-1細胞,按照上述方法檢測TFF2、VIL1和CDX2蛋白的表達。將GES-1細胞分為空白對照組、DCA組、陰性對照組、KLF5沉默組、氯化鋰LiCl(Wnt通路激活劑)組及LiCl+KLF5沉默組,按分組收集轉染48 h后的GES-1細胞,按照上述方法檢測Wnt3a、β-catenin和CDX2蛋白的表達。
1.2.7實時熒光定量聚合酶鏈反應(quantitative real-time polymerase chain reaction,qRT-PCR)分別取正常胃黏膜、腸上皮化生組織,以及DCA處理GES-1細胞樣品。采用Trizol法分別提取總RNA,并檢測其純度及完整性。按照逆轉錄試劑盒說明書生成cDNA,擴增cDNA,基因擴增成功后,采用2-△△Ct法,對KLF5和Wnt3a的含量進行分析。將GES-1細胞分為空白對照組、DCA組、陰性對照組及KLF5沉默組,按分組收集轉染48 h后的GES-1細胞,按照上述方法檢測TFF2、VIL1和CDX2 mRNA的表達。將GES-1細胞分為空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組,按分組收集轉染48 h后的GES-1細胞,按照上述方法檢測Wnt3a、β-catenin和CDX2 mRNA的表達。
1.3 統計學方法
數據分析采用SPSS 17.0統計軟件,計量資料以均數±標準差(±s)表示,比較用t檢驗或單因素方差分析,兩兩比較用SNK-q檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 KLF5和Wnt3a在腸上皮化生組織中異常表達
2.1.1KLF5和Wnt3a蛋白腸上皮化生組織中KLF5和Wnt3a蛋白相對表達量分別為(2.37±0.06)和(2.32±0.07);正常胃黏膜中KLF5和Wnt3a蛋白相對表達量分別為(1.02±0.08)和(1.00±0.07)。兩組KLF5和Wnt3a蛋白表達水平比較,經t檢驗,差異有統計學意義(t=80.350和98.002,均P=0.000),腸上皮化生組織中KLF5和Wnt3a蛋白表達升高。見圖1。

圖1 正常胃黏膜和腸上皮化生組織中KLF5、Wnt3a蛋白的表達(±s)
2.1.2KLF5和Wnt3a mRNA腸上皮化生組織中KLF5和Wnt3a mRNA相對表達量分別為(2.14±0.06)和(2.22±0.07);正常胃黏膜中KLF5和Wnt3a mRNA相對表達量分別為(1.00±0.06)和(1.00±0.08)。兩組KLF5和Wnt3a mRNA表達水平比較,經t檢驗,差異有統計學意義(t=70.910和70.817,均P=0.000),腸上皮化生組織中KLF5和Wnt3a mRNA表達升高。見圖2。
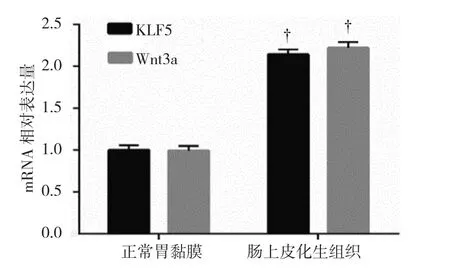
圖2 正常胃黏膜和腸上皮化生組織中KLF5和Wnt3a mRNA表達水平比較(±s)
2.2 DCA誘導GES-1細胞中KLF5和Wnt3a的表達
2.2.1KLF5和Wnt3a蛋白100、200和500μmol/L DCA組,以及空白對照組的KLF5蛋白相對表達量分別 為(1.26±0.07)、(1.63±0.06)、(2.47±0.06) 和(1.00±0.06);100、200和 500μmol/L DCA 組,以及空白對照組的Wnt3a蛋白相對表達量分別為(1.20±0.07)、(1.59±0.09)、(2.34±0.08)和(1.00±0.07)。各組細胞中KLF5和Wnt3a蛋白表達水平比較,經方差分析,差異有統計學意義(F=268.043和143.273,均P=0.000),隨著DCA濃度增加,GES-1細胞中KLF5和Wnt3a的表達水平逐漸升高。見圖3。
2.2.2KLF5和 Wnt3a mRNA100、200和 500μmol/L DCA組,以及空白對照組的KLF5 mRNA相對表達量分別為(1.24±0.08)、(1.74±0.06)、(2.21±0.06)和(1.00±0.05);100、200 和 500μmol/L DCA 組,以及空白對照組的Wnt3a mRNA相對表達量分別為(1.20±0.05)、(1.73±0.08)、(2.28±0.07)和(1.00±0.06)。各組細胞中KLF5和Wnt3a mRNA表達水平比較,經方差分析,差異有統計學意義(F=179.614和209.881,均P=0.000),KLF5和Wnt3a mRNA表達水平呈劑量依賴效應。KLF5和Wnt通路可能在胃黏膜腸上皮化生中發揮重要作用。見圖4。
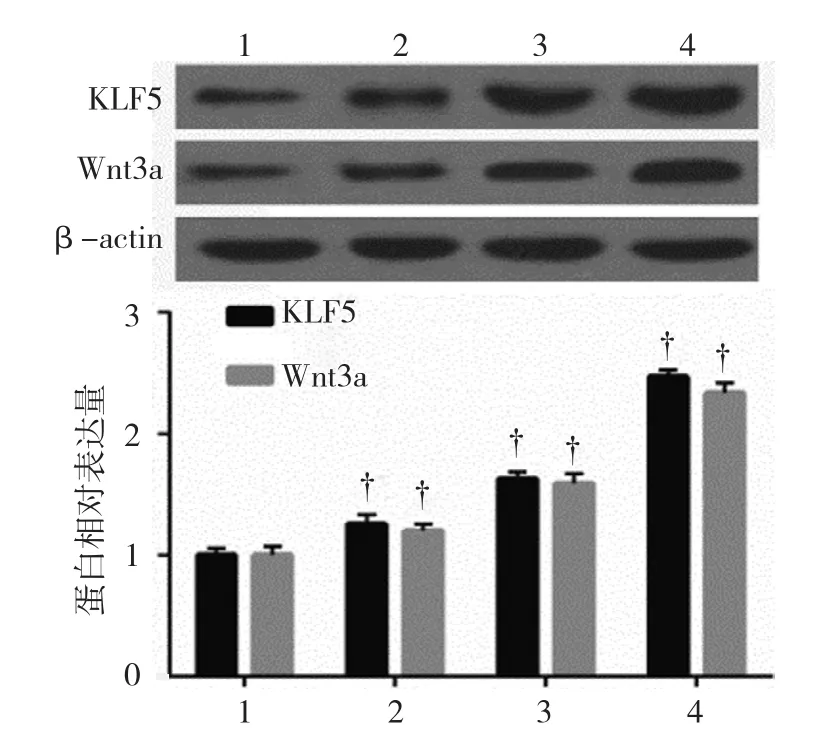
圖3 不同濃度DCA對GES-1細胞中KLF5和Wnt3a蛋白表達的影響(±s)
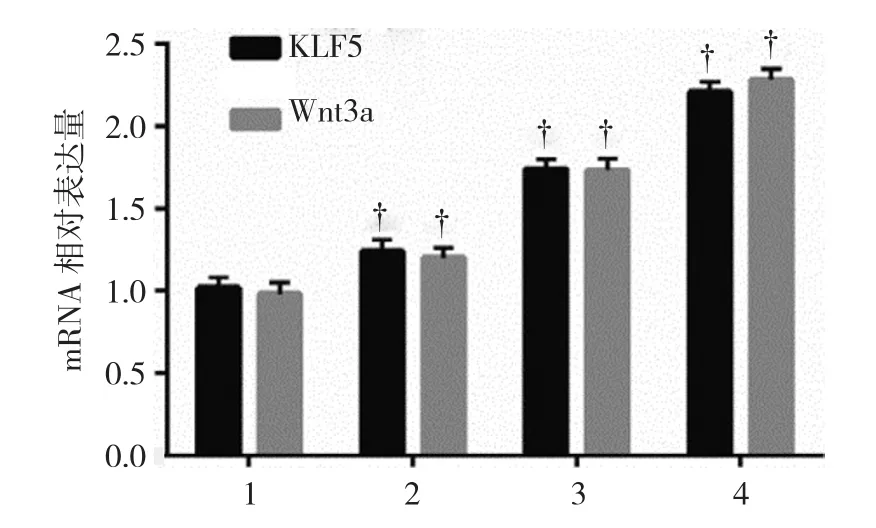
圖4 不同濃度DCA對GES-1細胞中KLF5和Wnt3a mRNA 表達的影響(±s)
2.3 KLF5沉默抑制DCA誘導的GES-1細胞增殖并促進凋亡
2.3.1KLF5蛋白空白對照組、DCA組、陰性對照組、KLF5沉默組的KLF5蛋白相對表達量分別為(1.00±0.06)、(2.45±0.08)、(2.36±0.06)和(0.11±0.06),經方差分析,差異有統計學意義(F=822.422,P=0.000),si-KLF5轉染后,細胞中KLF5蛋白表達降低。見圖5。
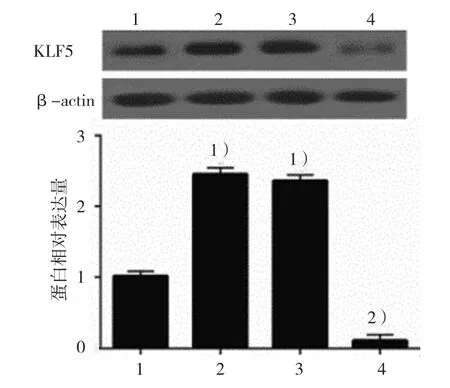
圖5 各組GES-1細胞中KLF5蛋白的表達(±s)
2.3.2增殖率空白對照組、DCA組、陰性對照組、KLF5沉默組GES-1細胞增殖率分別為(100.00±3.46)%、(125.00±3.60)%、(123.00±3.60)% 和(102.00±4.58)%,經方差分析,差異有統計學意義(F=36.136,P=0.000),DCA促進GES-1細胞增殖,而將KLF5沉默后,DCA的促增殖作用被抑制。見圖6。
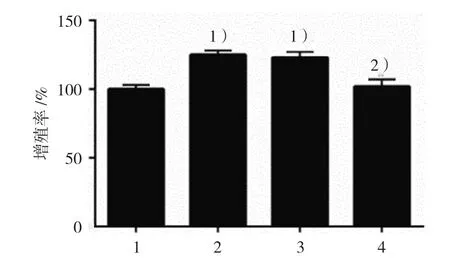
圖6 各組GES-1細胞增殖率比較(±s)
2.3.3凋亡率空白對照組、DCA組、陰性對照組、KLF5沉默組GES-1細胞凋亡率分別為(18.70±0.72)%、(10.30±0.70)%、(9.80±0.79)%和(17.80±0.79)%,經方差分析,差異有統計學意義(F=119.419,P=0.000),DCA 抑制GES-1細胞凋亡,而KLF5沉默后,DCA的抑制凋亡作用被抑制。DCA可能通過KLF5影響GES-1細胞的增殖和凋亡。見圖 7、8。

圖7 各組GES-1細胞凋亡情況
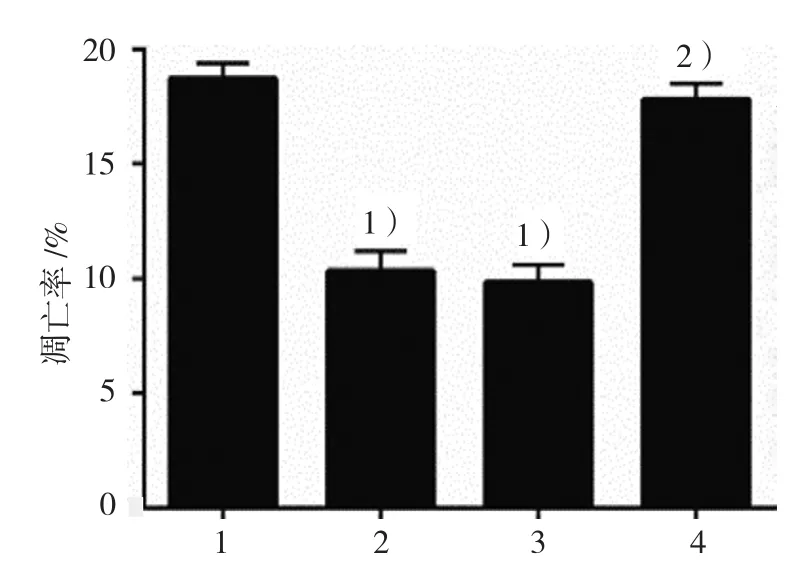
圖8 各組GES-1細胞凋亡率比較(±s)
2.4 KLF5沉默抑制DCA誘導的GES-1細胞腸上皮化生
2.4.1TFF2、VIL1和CDX2蛋白空白對照組、DCA組、陰性對照組、KLF5沉默組的TFF2蛋白相對表達量分別為(1.00±0.05)、(2.46±0.08)、(2.38±0.09)和(1.09±0.08),經方差分析,差異有統計學意義(F=301.250,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組的VIL1蛋白相對表達量分別為(1.00±0.05)、(2.39±0.09)、(2.37±0.07)和(1.11±0.05),經方差分析,差異有統計學意義(F=385.109,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組的CDX2蛋白相對表達量分別為(1.00±0.06)、(2.52±0.08)、(2.51±0.09)和(1.02±0.05),經方差分析,差異有統計學意義(F=413.707,P=0.000)。DCA升高TFF2、VIL1和CDX2蛋白表達水平,而KLF5沉默后DCA的作用被抑制。見圖9。
2.4.2TFF2、VIL1和CDX2 mRNA空白對照組、DCA組、陰性對照組、KLF5沉默組的TFF2 mRNA相對表達量分別為(1.00±0.05)、(2.35±0.05)、(2.37±0.06)和(1.11±0.08),經方差分析,差異有統計學意義(F=418.568,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組的VIL1 mRNA相對表達量分別為(1.02±0.03)、(2.32±0.05)、(2.29±0.05)和(1.10±0.07),經方差分析,差異有統計學意義(531.171,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組的CDX2 mRNA相對表達量分別為(1.00±0.04)、(2.41±0.08)、(2.43±0.05)和(1.05±0.08),經方差分析,差異有統計學意義(F=452.901,P=0.000)。DCA 升 高 TFF2、VIL1和CDX2 mRNA表達水平,而KLF5沉默后DCA的作用被抑制。DCA可能通過KLF5誘導胃黏膜腸上皮化生。見圖10。
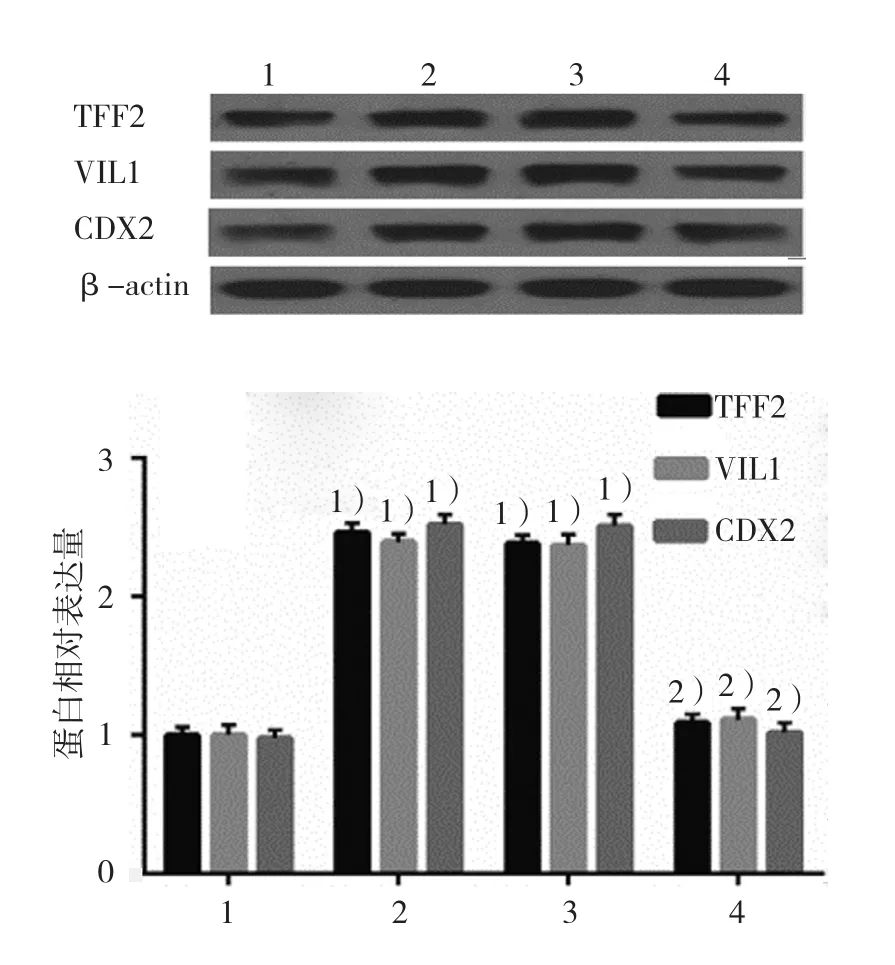
圖9 各組GES-1細胞中TFF2、VIL1和CDX2 蛋白的表達(±s)
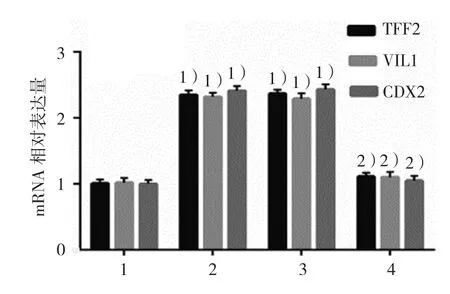
圖10 各組GES-1細胞中TFF2、VIL1和CDX2 mRNA表達水平比較(±s)
2.5 KLF5促進DCA誘導的GES-1細胞腸上皮化生
2.5.1Wnt3a、β-catenin和CDX2蛋白空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的Wnt3a蛋白相對表達量分別為(1.01±0.06)、(2.38±0.06)、(2.26±0.05)、(1.02±0.05)、(2.36±0.08) 和(2.32±0.08), 經 方差分析,差異有統計學意義(F=320.400,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的β-catenin蛋白相對表達量分別為(0.98±0.06)、(2.32±0.07)、(2.31±0.07)、(1.08±0.07)、(2.33±0.08) 和(2.40±0.07), 經 方差分析,差異有統計學意義(F=266.022,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的CDX2蛋白相對表達量分 別 為(1.02±0.06)、(2.48±0.06)、(2.34±0.08)、(1.02±0.08)、(2.37±0.05) 和(2.38±0.06), 經 方差分析,差異有統計學意義(F=312.093,P=0.000)。DCA激活Wnt通路Wnt3a、β-catenin,以及腸上皮化生標志蛋白CDX2蛋白的表達,將KLF5沉默后,Wnt通路蛋白和腸上皮化生標志蛋白表達下降,而Wnt激活劑則使抵消了si-KLF5的作用,激活Wnt通路并升高CDX2的表達。見圖11。
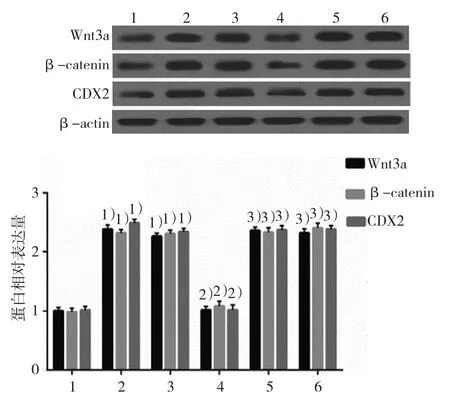
圖11 Wnt激動劑對si-KLF5調節GES-1細胞中Wnt3a、β-catenin和CDX2蛋白表達的影響(±s)
2.5.2Wnt3a、β-catenin 和 CDX2 mRNA空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的Wnt3a mRNA相對表達量分別為(1.00±0.06)、(2.32±0.07)、(2.28±0.06)、(1.09±0.06)、(2.26±0.06)和(2.23±0.06),經方差分析,差異有統計學意義(F=310.259,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的β-catenin mRNA相對表達量分別為(1.00±0.05)、(2.29±0.06)、(2.25±0.08)、(1.11±0.09)、(2.21±0.08)和(2.20±0.08),經方差分析,差異有統計學意義(F=190.732,P=0.000);空白對照組、DCA組、陰性對照組、KLF5沉默組、LiCl組及LiCl+KLF5沉默組的CDX2 mRNA相對表達量分別為(1.00±0.03)、(2.31±0.06)、(2.26±0.06)、(1.05±0.04)、(2.27±0.06)和(2.25±0.06),經方差分析,差異有統計學意義(F=400.107,P=0.000)。DCA激活Wnt通路Wnt3a、β-catenin,以及CDX2 mRNA的表達,將KLF5沉默后,Wnt3a、β-catenin和CDX2 mRNA表達下降,而Wnt激活劑則使抵消了si-KLF5的作用,激活Wnt通路并升高CDX2 mRNA的表達。KLF5可能通過激活Wnt/β-catenin通路促進DCA誘導的胃黏膜腸上皮化生。見圖12。
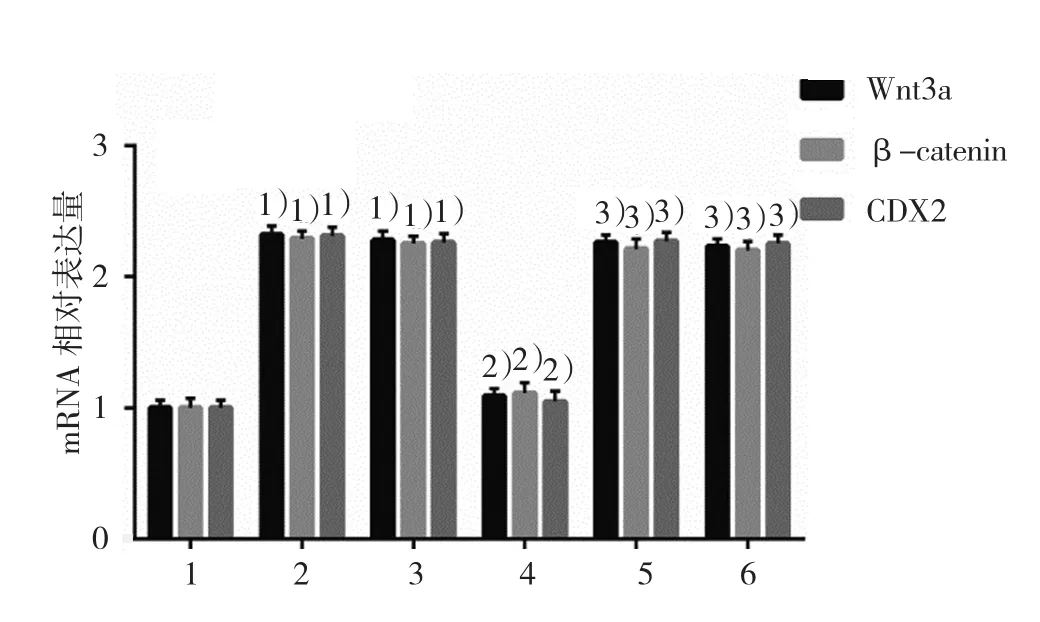
圖12 Wnt激動劑對si-KLF5調節GES-1細胞中Wnt3a、β-catenin和CDX2 mRNA表達的影響(±s)
3 討論
我國是胃癌高發國家,每年新發病例占全世界發病人數的絕大多數。決定胃癌患者預后的關鍵在于早期發現、診斷和治療。慢性萎縮性胃炎伴腸上皮化生及異型增生是重要的胃黏膜癌前病變[5-6]。因此,胃黏膜腸上皮化生發病機制的研究,可為胃黏膜異型增生甚至癌變的治療提供新思路。
Krüppel樣基因家族(Krüppel like factor,KLFs)為哺乳類動物的轉求調節因子,屬于鋅指蛋白轉錄因子家族。其中KLF5又稱為BTEB2和IKLF,屬于KLF家族[7]。KLF5在不同組織不同層次中有廣泛的表達。作為一個基本的轉錄因子,KLF5在細胞周期調控、細胞凋亡、遷移和分化中發揮必不可少的作用。另外,KLF家族成員KLF2、KLF4、KLF5及KLF8均證實作為致癌因子,促進各種癌癥的發生、發展[8-10]。有研究證明,KLF5協同其他轉錄因子促進胃癌的發展[11],而其在癌前病變中的作用并無相關報道。有研究表明,DCA可以誘導食管癌的癌前病變[2]。因此,本文用DCA誘導胃黏膜細胞,觀察KLF5沉默對胃黏膜腸上皮化生的作用。本實驗結果證明,與正常胃黏膜相比,KLF5在腸上皮化生中的表達升高,提示KLF5可能在胃黏膜腸上皮化生中發揮重要作用。進一步實驗結果表明,與正常胃黏膜上皮細胞相比,DCA誘導胃黏膜腸上皮化生細胞模型中KLF5的表達升高。
細胞增殖和凋亡是保持機體平衡和修復損傷的重要因素。胃黏膜上皮細胞更新非常迅速,研究認為腸上皮化生的發生是細胞異常增生逐漸積聚的結果[12]。有研究表明,抑制KLF5能夠減輕口腔黏膜癌前病變[13]。KLF5能夠促進乳腺癌細胞的增殖、遷移和侵襲,從而促進癌癥的發展[14]。本實驗結果表明,DCA促進GES-1細胞的增殖,并抑制凋亡,而KLF5沉默后抑制DCA對細胞增殖、凋亡的影響。
同源異形盒基因CDX2是一種腸上皮特異性轉錄因子,可控制腸上皮細胞的分化和成熟,在腸上皮化生黏膜中呈高表達[15]。三葉因子家族(trefoil factor family,TFF)是胃腸道黏液細胞分泌的具有≥1個三葉結構域的小分子蛋白質多肽,主要功能是保護黏膜、促進損傷黏膜修復。研究發現,在TFF2缺陷小鼠模型實驗中,小鼠的胃黏膜增生降低,提示TFF2可能有刺激胃黏膜增殖的作用,增加胃癌發生的危險性[16]。絨毛蛋白(Villin,VIL)是一種重要的細胞支架蛋白,在刷狀緣上皮細胞分化過程中發揮重要作用。近年來研究表明,VIL1是腸上皮細胞分化和轉移性腸腺癌的標志物,可以輔助明確轉移性腫瘤的來源[17]。因此,TFF2、VIL1和CDX2作為腸上皮化生的標志蛋白,在腸上皮化生過程中發揮重要作用。越來越多的研究表明,Wnt信號通路除參與調控發育過程外,在癌癥的發展中也發揮重要作用。Wnt途徑激活后,相關的細胞周期和凋亡相關基因、生長因子和黏附分子等開始轉錄,參與癌癥的發生、發展[18]。有研究表明,KLF5敲除的小鼠中Wnt通路被抑制,腸上皮穩態被破壞[19]。本實驗結果表明,KLF5沉默抑制DCA對TFF2、VIL1和CDX2的上調作用;另外,在Wnt通路干預后,si-KLF5的作用又被抑制,表明KLF5可能通過Wnt/β-catenin通路促進DCA誘導的腸上皮化生。
綜上所述,本文通過DCA誘導胃黏膜上皮細胞,發現KLF5沉默明顯抑制DCA誘導的胃黏膜上皮細胞腸上皮化生,表明KLF5可能促進DCA誘導的腸上皮化生,其機制可能是通過激活Wnt/β-catenin通路來實現的。本研究指出KLF5參與DCA誘導的胃黏膜腸上皮化生的發生,為預防胃癌提供新的靶點。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
