轉Bt基因抗蟲玉米HGK60的農藝性狀分析
2018-08-18 01:44:18梁海生李夢桃李圣彥汪海張杰郎志宏
生物技術通報 2018年7期
關鍵詞:差異
梁海生 李夢桃 李圣彥 汪海 張杰 郎志宏
(1. 西南科技大學生命科學與工程學院,綿陽 621010;2. 中國農業科學院生物技術研究所,北京100081;3. 中國農業科學院植物保護研究所/植物病蟲害生物學國家重點實驗室,北京 100193;4. 東北農業大學生命科學學院,哈爾濱 150030)
玉米是我國三大農作物之一,種植面積和產量居三大作物之首[1]。2016年玉米播種面積為3.677×107hm2,年產量約為2.2億噸。我國雖然能實現主糧的基本自給,但農產品缺口較大,2013年玉米籽粒進口326.42萬噸,2014年玉米籽粒進口259.90萬噸。病蟲害是造成玉米減產的重要因素之一,玉米每年受螟蟲危害的面積超過2.335×107hm2,約占種植總面積的70%,2013年造成減產639萬噸、2014年造成減產544.78萬噸。害蟲危害不僅直接造成減產,還常引發玉米穗腐病,進而使籽粒中真菌毒素含量增高,品質下降[2]。因此,迫切需要安全有效的措施控制蟲害,減少農業生產投入,增加農民收入。
傳統防治蟲害是在生長過程中噴施化學殺蟲劑和生物防治。但前者不僅能殺死害蟲,也能殺死害蟲的天敵,農藥殘留問題還會造成部分農民中毒傷亡,害蟲的耐藥性增加,也導致農藥用量增加,破壞生態平衡、污染環境;后者釋放赤眼蜂等方法受氣候條件影響較大,防治效果不理想[3]。而將外源抗蟲基因轉入玉米中為培育抗蟲玉米新品種提供了新的思路。1995年瑞士先正達公司通過基因槍法將來源于蘇云金芽胞桿菌的cry1Ab轉入玉米中獲得的BT176轉化事件,以bar作為篩選標記,這是世界第一例商業化種植轉基因抗蟲玉米。1996年美國孟山都公司將改良后的cry1Ab基因利用基因槍法轉入玉米中,獲得抗玉米螟的MON810轉基因玉米[4]。在轉基因作物應用的20多年間,全球農藥使用量大幅減少,為可持續發展作出了重要貢獻,1996-2015年,節約了6.2 億kg的農藥用量,同比減少8.1%,使環境影響商數(EIQ)降低了19%。同時轉基因作物的種植為全球糧食產量帶來大幅增長,1996-2015年,作物產量增加5.74億t,產值增加1678億美元[5]。而在歐洲玉米螟(European Corn Borer,ECB)危害嚴重的情況下,轉Bt基因的雜交玉米品種比非轉基因雜交品種增產12%-23%[6];在自然ECB發生情況下,轉Bt基因雜交種比非轉基因雜交種增產 5.5%[7]。Elisa等[8]通過分析 21年的同行文獻得出轉基因玉米雜交種的玉米產量比其近等基因系要高5.6%-24.5%。Wang等[9]對比了轉基因雜交玉米12-5×IE034和2個親本轉基因玉米以及
6個常規玉米品種中的轉錄譜和代謝譜的差異,發現轉基因玉米育種基因疊加所帶來的基因表達和代謝物的變化數量介于常規玉米品種間的變異范圍之內。到目前為止,市場中的轉基因抗蟲玉米品種全部為國外公司所擁有。近年來,我國雖然在轉基因工作中取得重大進展,但尚未有一例商業化抗蟲轉基因玉米品種上市。本研究使用的cry1Ah是從國內篩選的BT8菌株中鑒定克隆的新型Bt基因,具有自主知識產權[10],其編碼的殺蟲蛋白的殺蟲活性優于目前使用的cry1Ab和cry1Ac[11]。前期的研究結果表明轉Bt cry1Ah玉米在溫室和田間具有良好的抗蟲效果,證明cry1Ah可作為培育商業化抗蟲玉米的候選基因[12-13]。標記基因選用2 m G2-epsps,該基因來源于熒光假單胞菌G2,編碼5-烯醇式丙酮酰莽草酸-3-磷酸合酶(EPSPS),具有草甘膦耐受性[14]。為了獲得滿足商業化需求的轉基因抗蟲玉米品種,前期通過大規模遺傳轉化進行群體篩選,獲得一個Bt蛋白表達量高且對亞洲玉米螟有顯著抗性的轉化事件HGK60[15]。本研究對HGK60及其雜交種進行了兩地連續三代的分子檢測及農藝性狀分析,證明HGK60轉基因抗蟲玉米中外源基因能夠穩定的遺傳和表達,對亞洲玉米螟具有很好的抗性,農藝性狀與對照材料相比無顯著差異。這將推動HGK60轉基因抗蟲玉米的產業化應用。
1 材料與方法
1.1 材料
1.1.1 植物材料 玉米自交系鄭58(輪回親本)、轉Bt cry1Ah基因抗蟲玉米材料HGK60的BC5F2(與鄭58回交5代,自交2代,下同,2016年海南)、BC6F2(2017年廊坊)、BC7F2(2017年海南)及HGK60的BC5F2(2016年海南)、BC6F2(2017年廊坊)、BC7F2(2017年海南)分別與昌7-2的雜交種鄭單958H,雜交種鄭單958,由中國農業科學院生物技術研究所郎志宏課題組保存。
1.1.2 供試蟲源 亞洲玉米螟由北京美延農業科技有限公司購買。
1.2 方法
1.2.1 PCR檢測轉基因植株 取長至3葉期的玉米幼苗葉片采用CTAB法提取基因組DNA,選用cry1Ah特異性引物和篩選標記基因2mG2-epsps特異性引物對其進行PCR檢測,留存陽性植株。cry1Ah特異性引物序列為F1:5'-ATGGGCAAGAACTCCATCAA-3'和 R1:5'-TCAGTCGATGTGGTAGTCGGTAAC-3',擴增產物長度為2007 bp。反應條件為94℃5 min ;95℃ 20 s,55℃ 30 s,72℃ 1 min,32 個循環;72℃ 5 min。2mG2-epsps特異性引物序列為 F2:5'-ATGGCGTGCCTCCCTGACGA-3'和 R2:5'-TCATCAGTCGTTGAGGTGAA-3',擴增產物長度為1338 bp,反應條件與上述一致。
1.2.2 實時熒光定量PCR檢測轉基因植株 為了比較不同生長發育階段陽性植株在轉錄水平的差異,對PCR陽性植株進行實時熒光定量PCR(quantitative RT-PCR)檢測。取玉米不同生長時期(抽穗期、灌漿期)的展開葉、雄穗、穗軸、花絲、苞葉和籽粒組織,提取總RNA,利用反轉錄試劑盒(Thermo公司,美國)合成cDNA,采用SYBR GreenⅠ定量PCR試劑盒(Roche公司,瑞士)。反應體系為2×FastStart Universal SYBR Green Master(Rox)10 μL、上游引物(10 μmol/L)0.4 μL、下游引物(10 μmol/L)0.4 μL、cDNA 模板 2 μL,加水補至 20 μL。反應條件為 95℃ 10 min;95℃ 15 s,60℃ 60 s(采集熒光信號),共40個循環。將SYBR GreenⅠ的擴增產物從60℃緩慢而均勻地以0.3℃/s的速率升溫到95℃,記錄熒光信號的變化,繪制出擴增產物的溶解曲線。內參基因為玉米內參actin,引物 為 F3:5'-CGAATGCCCAGCAATGTA-3'和 R3:5'-TTAGGTGGTCGGTGAGGT-3',擴增產物長度為188 bp。
1.2.3 轉基因玉米Cry1Ah蛋白表達含量分析 取玉米不同生長時期(六葉期、大喇叭口期、抽穗期、灌漿期)的心葉、展開葉、莖、雄穗、花絲、穗軸、苞葉、根和籽粒組織,提取可溶性總蛋白,利用酶聯免疫吸附測定(Enzyme-linked immunosorbent assay,ELISA)轉基因植株的Cry1Ah蛋白表達量,以純化的Cry1Ah蛋白制作標準曲線。蛋白提取及ELISA檢測方法均按試劑盒說明書進行,ELISA檢測試劑盒購自美國Envirologix公司。
1.2.4 轉基因玉米高代自交系和雜交組合的主要農藝性狀考察 2016年12月-2018年3月,對轉基因抗蟲玉米進行了兩地連續三代田間試驗,地點為河北省中國農業科學院國際農業高新技術產業園生物技術示范基地和海南省中國農業科學院南繁基地。試驗地分小區種植,設3次重復,重復內各材料隨機排列。按照鄭58、HGK60、鄭單958及鄭單958H分4個小區,每個小區種植10 行,行長5 m,行寬0.60 m,每行種植20株。試驗地不進行任何蟲害的治理,其他(如水、肥以及除草等)完全與常規大田管理一樣。生長期間調查出苗期、吐絲期等生育期。在每個品種內隨機選取100株,吐絲后測量株高、穗位高,收獲后考種,于室內測量穗長、穗粗、禿尖長、穗行數、行粒數等農藝指標,果穗干燥后脫粒稱重,并求千粒重。
1.2.5 玉米種子發芽率檢測 對收獲的玉米種子進行發芽率檢測,按GB/T 3543.3規定的方法進行。采用紙床紙間發芽方法,先將玉米種子表面消毒(2%的次氯酸鈉溶液浸泡10 min),然后將濾紙放在培養皿內,用水將濾紙濕潤,將消毒后的種子均勻放置在濕潤的濾紙上,再用另一張同樣大小的濾紙覆蓋在種子上,蓋上培養皿,放入25℃光照培養箱內。每個培養皿擺放13粒種子,每個品種3皿為一組(統計單位),重復3次,每個品種共計9皿,117粒種子。發芽過程中,逐日統計正常發芽種子的粒數,4 d統計發芽勢,7 d統計發芽率。
1.2.6 玉米螟室內外生測
1.2.6.1 玉米螟田間生測 按照農業部953號公告-10-2007在大田進行隨機區組設計。對小喇叭口期的鄭58、HGK60、鄭單958及鄭單958H,以每株40-60頭玉米螟初孵幼蟲的用量接于心葉中,3 d后重復接蟲一次,吐絲期除接蟲部位為玉米花絲外其他同上。在接蟲14-21 d后,按照公告標準進行等級判定。
1.2.6.2 玉米螟室內生測 按照宋苗等[15]方法,采集生長至小喇叭口期玉米心葉,用細毛筆輕輕將亞洲玉米螟初孵幼蟲接入放有玉米心葉的24 孔生測皿中,每24 h調查幼蟲存活數,每個試驗材料重復3次。
1.2.7 數據分析 利用Excel軟件進行數據整理,用SPSS數據處理軟件對試驗數據進行分析,對不同處理間平均數的差異顯著性比較均采用t檢驗。
2 結果
2.1 轉基因玉米外源基因cry1Ah和標記基因2mG2-epsps的PCR檢測
對每一代的轉Bt cry1Ah抗蟲玉米材料HGK60自交系中的cry1Ah和2mG2-epsps進行PCR檢測,留取陽性植株與昌7-2雜交,收獲的鄭單958H種子種植至六葉期,取葉片檢測陽性率。以cry1Ah和2mG2-epsps特異性引物分別檢測目的基因和標記基因,以表達載體為模板作為陽性對照,以鄭58玉米基因組DNA為模板作為陰性對照。結果顯示,含目的基因cry1Ah的轉基因HGK60和鄭單958H擴增片段為2007 bp,與陽性對照片段大小一致,陰性對照鄭58未擴增出目的條帶,2017廊坊樣品HGK60均有陽性條帶,鄭單958H也都有陽性條帶(圖1);圖2顯示,含目的基因2mG2-epsps的轉基因HGK60擴增出片段為1338 bp,與陽性對照片段大小一致,陰性對照鄭58未擴增出目的條帶,2017海南樣品HGK60均有陽性條帶,與昌7-2的雜交種均有陽性條帶。初步判斷轉Bt cry1Ah抗蟲玉米中cry1Ah和2mG2-epsps均能穩定遺傳給子代個體。
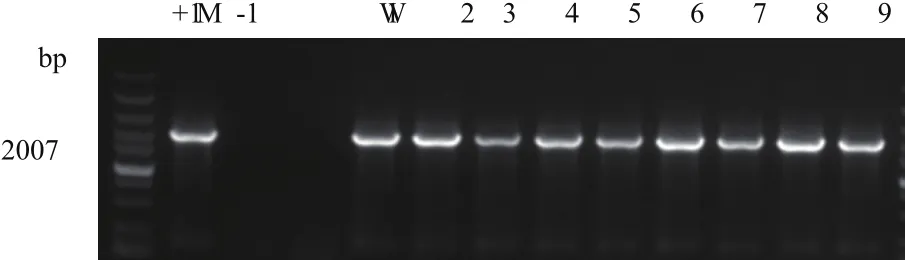
圖1 以cry1Ah為引物擴增部分2017廊坊HGK60和鄭單958H材料PCR電泳圖
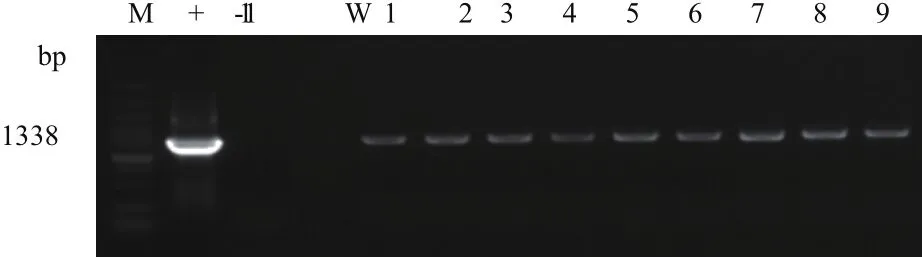
圖2 以2mG2-epsps為引物擴增部分2017海南HGK60和鄭單958H材料PCR電泳圖
2.2 實時熒光定量PCR檢測
分別取2017年海南生長至抽雄期、灌漿期的HGK60陽性植株及其雜交種鄭單958H各組織進行熒光定量PCR檢測(圖3),以灌漿期HGK60籽粒中的cry1Ah轉錄水平為參照,HGK60和鄭單958H均為灌漿期的展開葉(穗上葉)中cry1Ah轉錄水平最高,而穗軸和籽粒的轉錄水平較低;抽穗期鄭單958H中cry1Ah轉錄水平最高的部位是雄穗,而HGK60中的轉錄水平最高部位為展開葉。將灌漿期與抽穗期相同組織中cry1Ah轉錄水平進行比較,結果顯示,HGK60的展開葉與苞葉在這兩個時期中cry1Ah轉錄水平差異顯著;而鄭單958H僅有展開葉中cry1Ah轉錄水平差異顯著。
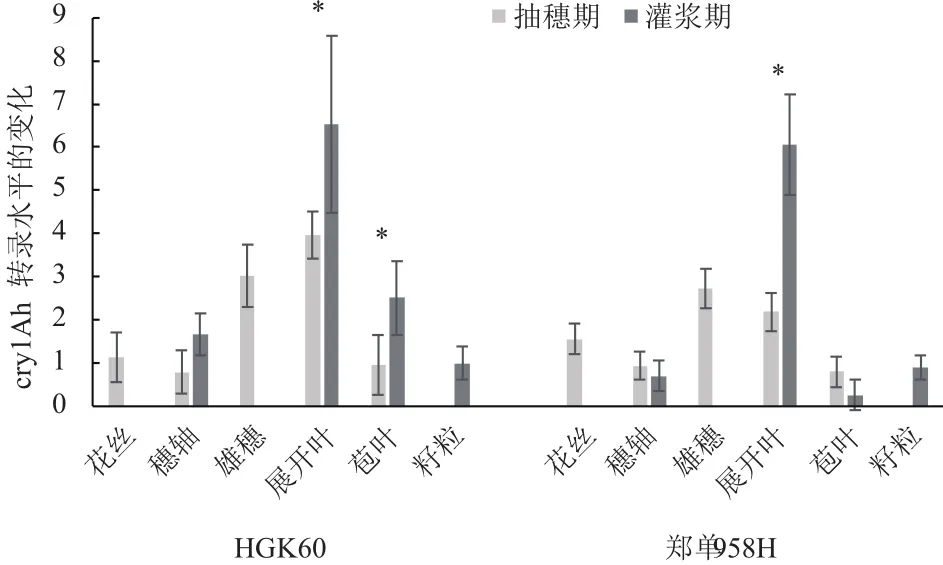
圖32017海南HGK60與鄭單958H不同時期不同組織cry1Ah的轉錄水平
2.3 ELISA檢測Cry1Ah表達量結果
利用ELISA方法檢測兩地連續三代HGK60及其雜交種不同時期不同部位的Cry1Ah蛋白表達量。結果(圖4)顯示,在2016年海南、2017年廊坊、2017年海南HGK60及其雜交種同一生長期同一部位的Cry1Ah蛋白表達量趨于穩定,不同生長期不同部位的Cry1Ah蛋白表達量存在差異。在抽穗期葉片中Cry1Ah蛋白表達比六葉期增加30.71%,在灌漿期雄穗中Cry1Ah蛋白表達量最高,為4017 ng/g(鮮重,下同)。在灌漿期中苞葉及花絲的Cry1Ah蛋白表達量比抽雄期分別增加41.62%和77.05%。灌漿期籽粒所含Cry1Ah蛋白表達量為1467 ng/g。表1所示,2017海南HGK60抽穗期中雄穗Cry1Ah表達量最高,為5720 ng/g,灌漿期中籽粒和穗軸表達量分別為2630和1106 ng/g;雜交種鄭單958H與自交種HGK60都是在抽穗期雄穗表達量處于最高水平,而穗軸表達量在灌漿期顯示下降的趨勢。雜交種的Cry1Ah表達量要比自交種HGK60低。
ELISA與實時熒光定量PCR結果相比較,在抽穗期HGK60的雄穗Cry1Ah蛋白表達量最高,而抽穗期的展開葉的轉錄水平最高,其雜交種中的cry1Ah雄穗轉錄水平最高。這種差異可能與環境差異有關,Miluse Trtikova通過比較2種的轉MON810品種發現由于遺傳背景和環境差異能造成轉基因表達量和Bt蛋白含量的顯著差異[16]。而苞葉和花絲中的蛋白表達水平和轉錄水平均偏低。灌漿期中展開葉的Bt蛋白表達水平與轉錄水平均比抽穗期要高,自交系和雜交種均如此,而籽粒中的蛋白表達水平和轉錄水平均偏低。

圖42017廊坊HGK60不同時期不同組織Cry1Ah蛋白表達

表12017海南部分HGK60和鄭單958H不同時期不同組織Cry1Ah蛋白表達
2.4 轉基因玉米高代自交雜交系的主要農藝性狀結果
2.4.1 轉基因對玉米生育期的影響 從表2可以看出,轉基因抗蟲玉米鄭單958H與鄭單958在同一世代出苗期一致,抽雄期、吐絲期及生育期略有差異,但只有1-2 d,差異不大。播種到抽雄時間2017年海南、2017年廊坊和2016年海南分別為47、65-66和42-43 d。生育期各代分別是95、125-126和92-93 d。各世代從播種到抽雄時間以及生育期不同,這主要是因為河北廊坊及海南三亞在不同時期的有效積溫及土壤類型不同造成的。轉基因抗蟲玉米HGK60與鄭58亦如此。
從3個世代的物候期調查結果可以得出轉入Bt cry1Ah對玉米生育期的影響不大。
2.4.2 轉基因對玉米株高、穗部及產量性狀的影響 對2個世代的轉cry1Ah抗蟲玉米的農藝性狀進行了田間跟蹤調查及實驗室考種工作。由表3可知,鄭單958H與鄭單958在不接玉米螟時,在同一世代株高、穗位高、穗長、穗粗、穗行數、禿尖長、行粒數、千粒重雖有差異,但差異不大,經檢驗差異不顯著;但鄭單958H的產量與鄭單958的差異比較大,除了2017廊坊的產量差異不顯著,在2017廊坊和2017海南鄭單958H比鄭單958分別增產7.91%和9.1%。在小喇叭口期和吐絲期接玉米螟后,鄭單958H與鄭單958同一世代穗長、穗行數、禿尖長、行粒數、千粒重雖有差異,但差異不大,經檢驗差異不顯著;但鄭單958H的株高在2個世代比鄭單958均差異較大,達極顯著水平;在2017海南鄭單958H的產量比鄭單958增產10.15%,經檢驗差異顯著。
不同世代間同一性狀的差異主要是由于試驗地點的土壤氣候條件不同造成的,總體趨勢為,在海南種植的植株株高和穗位高度比在廊坊種植的低,接蟲后的產量要比不接蟲的產量低,接蟲后的轉基因植株產量要比非轉基因產量要高,且差異顯著。隨著育種代數的增高,田間單株間已經變化不大,即各群體的農藝性狀隨著世代的增加更加趨于穩定。為轉Bt基因玉米自交系能夠直接用于育種提供了有力證據。
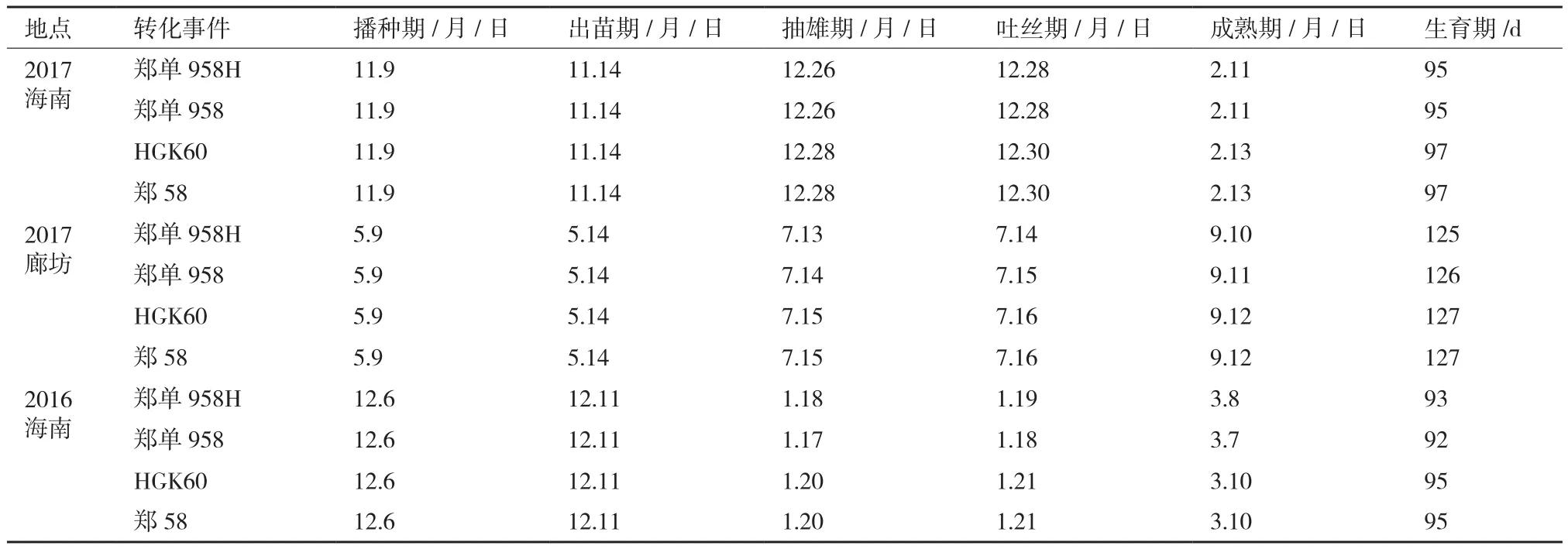
表2 不同年份不同材料的物候期
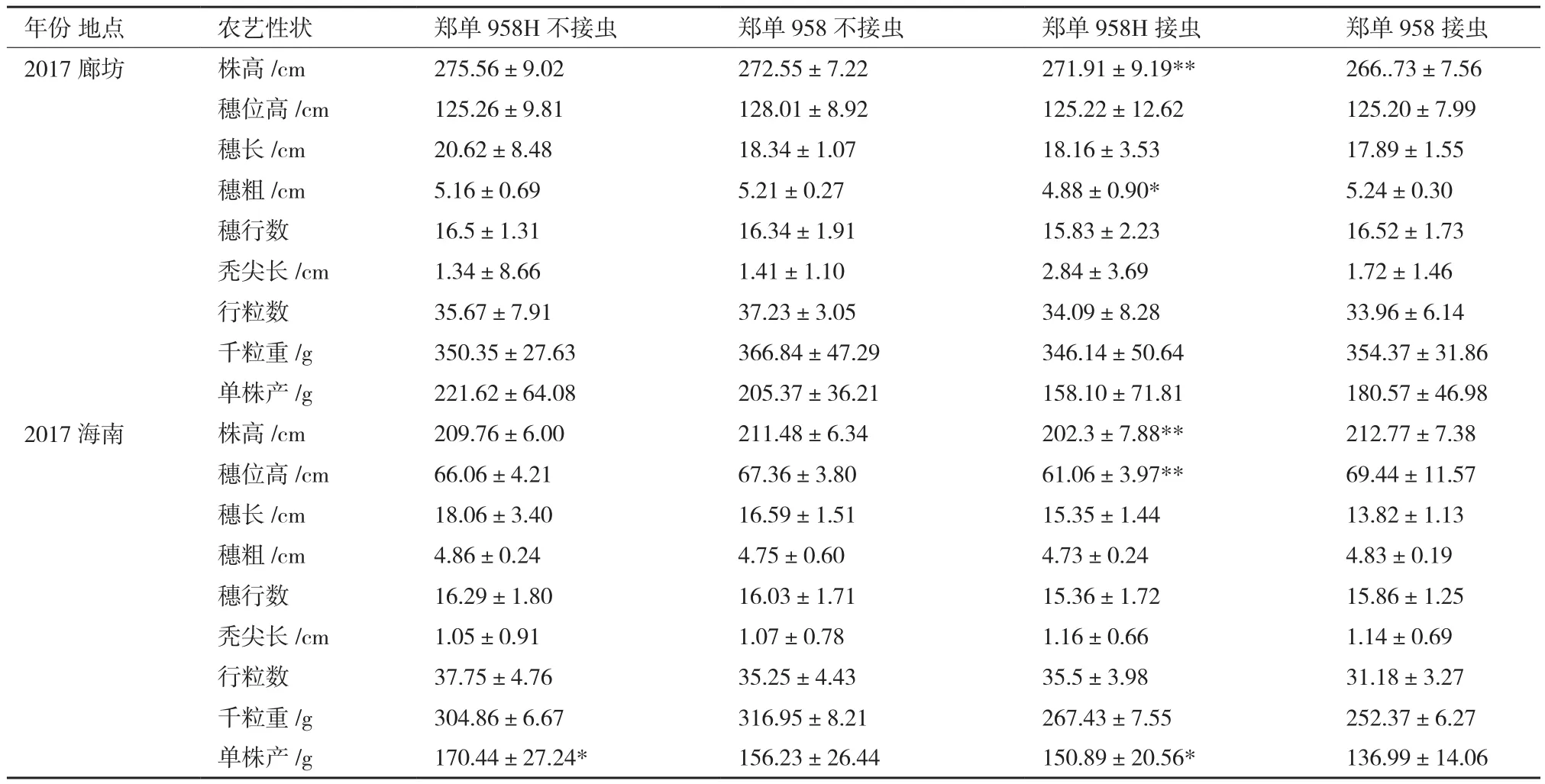
表3 不同年份的鄭單958H和鄭單958農藝性狀
2.5 轉基因對玉米種子發芽率的影響
由圖5及表4可以得出,轉基因玉米鄭單958H與非轉基因玉米鄭單958進行比較,在發芽勢、發芽率和發芽指數上存在一定的差異,經t檢驗,進行差異顯著性分析,P值分別為0.3302、0.2174和0.3958均大于0.05,因此差異不顯著。轉基因玉米HGK60與非轉基因玉米鄭58也在發芽勢、發芽率和發芽指數上存在一定的差異,經t檢驗,進行差異顯著性分析,P值分別為0.3143、0.4293和0.3518均大于0.05,因此差異不顯著。說明轉基因玉米鄭單958H與非轉基因玉米鄭單958及HGK60與鄭58在發芽勢、發芽率和發芽指數上沒有顯著差異,外源基因Bt cry1Ah的導入對玉米發芽活力方面沒有影響。
圖5 種子發芽4 d試驗
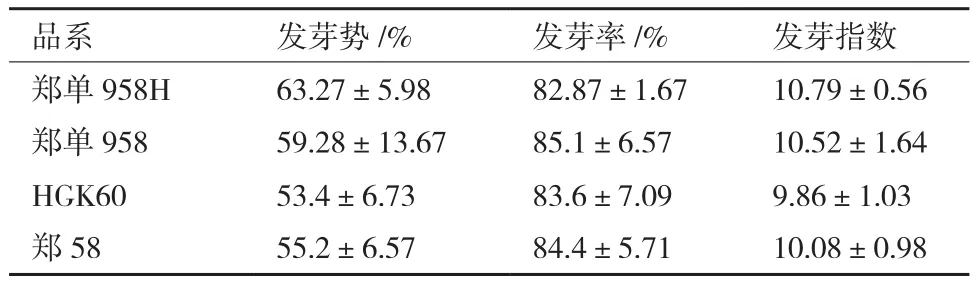
表4 玉米種子發芽率
2.6 玉米螟室內和室外生物活性檢測結果
2.6.1 玉米螟室內生測 室內接蟲3 d后(圖6),轉基因玉米中玉米螟幼蟲全部死亡,葉片鮮嫩,基本無危害,且蟲孔只有針孔大小。第7天時陰性對照鄭58活蟲個體較大,有較強的爬行活力,葉片受玉米螟危害較嚴重,有較多蟲孔。室內接蟲3 d后,咬食HGK60和鄭單958H死亡率如表5所示,取食轉基因玉米心葉的玉米螟幼蟲死亡率顯著高于陰性對照,經t檢驗,差異極顯著(P<0.01),表明轉Bt cry1Ah玉米材料HGK60和鄭單958H對玉米螟幼蟲有顯著殺蟲效果,該結果與宋苗等[15]結果一致。

表5 取食HGK60及鄭單958H葉片的亞洲玉米螟幼蟲死亡率/%

圖6 取食HGK60葉片的亞洲玉米螟室內生測
2.6.2 玉米螟田間生物活性檢測 由圖7可知,轉基因玉米的葉片在接蟲兩周后基本無玉米螟危害,只有少量針孔大小的蟲孔,而陰性對照葉片受害嚴重,有多個超過火柴頭大小的蟲孔。按照判定標準,HGK60田間抗性等級是1.27±0.61級,為高抗級別,相同處理的陰性對照平均抗性等級為6.59±1.43級,為感蟲級別。玉米吐絲期將玉米螟幼蟲接于玉米花絲,HGK60和鄭單958H玉米的雄穗和雌穗受害微小,按照判定標準抗性級別為1.74±0.52級,屬高抗級別,而陰性對照的雄穗和雌穗受害較嚴重,抗性級別為7.58±1.95級,屬感蟲級別。證明HGK60及其雜交種對玉米螟有很強的抗性,此結果與室內抗蟲性鑒定結果一致。田間生測結果同時也與宋苗等[15]生測結果一致,證明該材料具有良好的田間抗蟲穩定性。室內和田間蟲測結果與熒光定量PCR、ELISA結果一致。

圖7 轉基因玉米的田間生測
3 討論
采用農桿菌介導法獲得的轉基因植株,整合到植物基因組中的外源基因一般遵循孟德爾遺傳分離規律并能夠穩定遺傳和表達[17]。Paz等[18]將已經商業化17年的3種MON810轉基因玉米進行Southern雜交,在插入序列和側翼序列均未發現序列重排,進一步分析28個不同的MON810品種基因組側翼序列中存在的點突變和插入缺失,得出轉入的基因突變率與玉米內源基因的突變率相同的結論。兩地連續三代種植轉基因玉米HGK60及其雜交種鄭單958H經過PCR檢測、實時熒光定量PCR檢測結果顯示能穩定遺傳與表達Bt cry1Ah基因。新表達蛋白Cry1Ah表達量在同一生長期同一部位趨于穩定,不同生長期不同部位存在差異。研究發現雄穗中Cry1Ah表達量在玉米抽穗期和灌漿期均居于最高水平,葉片的表達量次之,而苞葉和花絲的蛋白表達量在灌漿期增高。玉米螟在抽穗期取食幼嫩的雄穗,雄穗中高表達的Bt蛋白能有效防治玉米螟的危害。玉米螟在灌漿期咬食花絲、苞葉和籽粒,苞葉和花絲的表達量提升能減少授粉期間玉米螟對苞葉和花絲的危害。田間生測HGK60和鄭單958H雄穗沒有出現折斷及被咬食的情況,而大部分鄭58和鄭單958的雄穗在玉米吐絲期被咬食嚴重甚至被折斷。HGK60和鄭單958H的苞葉、花絲和籽粒玉米螟均無危害,而鄭58和鄭單958的苞葉有大量蟲孔,花絲被咬食嚴重,籽粒有部分發霉。本研究表明2個材料均表現較好的抗螟性。在抽穗期和灌漿期,HGK60和鄭單958H在雄穗、展開葉和苞葉均有較高的轉錄水平,這與蛋白表達水平基本吻合。造成鄭單958H在抽穗期葉片的轉錄水平比雄穗低,與自交系HGK60的結果相反,而蛋白水平是雄穗的要比葉片的高,造成RNA轉錄水平和蛋白質表達水平不一致的原因還未做深入研究。Makarevitch等[19]研究認為特定品種的遺傳背景可以影響轉基因的表觀遺傳調控,DNA甲基化[20]、組蛋白修飾[21]、非編碼RNA調控[22]等可以造成基因序列不發生改變而轉錄水平產生差異,其是否與表觀遺傳調控相關,還有待進一步研究。
從3個世代的物候期調查結果可以得出轉入Bt cry1Ah對玉米生育期的影響不大。而玉米生育期與緯度變化及種植當地的積溫相關,有研究統計顯示隨著緯度的北移,播種-出苗和出苗-吐絲的時間顯著增加,緯度每增加1°,播種-出苗和出苗-出吐絲的時間分別延長0.7 d和1.25 d,吐絲-出成熟的時間縮短0.8 d,玉米營養生長期占總生育期的比值隨著緯度的北移顯著增加,緯度每增加1°,比值增加1.34[23]。海南的積溫比廊坊積溫要高,緯度比廊坊低,在海南玉米的生育期要比在廊坊玉米的生育期短。因此,統計的3個世代物候期的差異是由于積溫和緯度等因素造成的,與轉入的Bt cry1Ah基因無關。
康領生等[24]將cry1F轉入玉米獲得轉基因玉米SW12-859,接蟲與不接蟲小區和鄭單958不接蟲小區比較,玉米株高、穗位高、產量無顯著差異,玉米株型、生育期都一致。玉米SW12-859與鄭單958通過接蟲區測產、考種比較,玉米穗長、穗粗、百粒重、產量存在顯著差異。本研究中鄭單958H與鄭單958相比,不接玉米螟時,在同一世代株高、穗位高、穗長、穗粗、穗行數、禿尖長、行粒數、千粒重無顯著差異,接蟲后的產量要比不接蟲的產量低,接蟲后的轉基因植株產量要比非轉基因產量要高,且差異顯著。接蟲后的鄭單958H和鄭單958產量都降低,這可能與田間模擬大量蟲害爆發有關,葉片被咬食嚴重,受害葉片喪失功能,直接影響玉米光合作用,導致玉米植株生長發育受阻,株高和產量受損。研究表明,培育出的鄭單958H雜交種有較好的育種價值。
從玉米種子上來看,本研究中轉基因玉米鄭單958H與非轉基因玉米鄭單958及HGK60與鄭58在發芽勢、發芽率和發芽指數上沒有顯著差異,外源Bt cry1Ah基因的導入對玉米發芽活力方面沒有影響。這與王延鋒[25]將轉基因玉米與受體X90進行比較得出外源cry1Ah的導入對玉米發芽活力沒有影響結果一致。
無論是自交種HGK60還是雜交種鄭單958H,都顯示出比鄭58及鄭單958較強的抗蟲優勢。在農藝性狀等基本保持穩定的前提下,還具有能穩定遺傳的良好抗蟲性,具有較強的育種前景。
4 結論
轉基因抗蟲玉米自交種HGK60及雜交種鄭單958H具有良好的遺傳穩定性和抗蟲性,農藝性狀趨于穩定。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
