獺兔CREB1基因的編碼區(qū)克隆、生物信息學(xué)及表達規(guī)律分析
2019-01-09 07:00:40秦立志趙博昊胡帥帥翁巧琴鮑國連吳信生
江蘇農(nóng)業(yè)科學(xué) 2018年23期
陳 陽, 秦立志, 趙博昊, 胡帥帥, 穆 琳, 翁巧琴, 鮑國連, 吳信生
(1.揚州大學(xué)動物科學(xué)與技術(shù)學(xué)院,江蘇揚州 225009; 2.浙江省余姚市欣農(nóng)兔業(yè)有限公司,浙江余姚 315400;3.浙江省農(nóng)業(yè)科學(xué)院,浙江杭州 310021)
獺兔,即力克斯兔(Rex rabbit),是皮用型兔的代表之一,其被毛顏色豐富多樣,且純天然的被毛不掉色或褪色,深受消費者青睞。目前,美國認可的獺兔毛色有14種,歐洲認可的有30種,我國以青紫藍色、黑色、海貍色、白色為主。一般而言,動物的毛色與皮膚中黑色素的數(shù)量和分布密切相關(guān),同時許多內(nèi)在因子也影響著毛色性狀的表現(xiàn)[1]。哺乳動物毛色主要取決于黑色素沉積,黑色素細胞產(chǎn)生真黑素和棕黑素,二者分布的不同致使哺乳動物形成多種毛色類型[2-3]。筆者所在課題組前期利用表達譜芯片篩選與獺兔毛色形成相關(guān)的基因,發(fā)現(xiàn)CREB1基因在不同毛色的獺兔皮膚中表達差異顯著,表明該基因可能參與到獺兔的毛色形成過程中[4]。
環(huán)腺苷酸反應(yīng)元件結(jié)合蛋白(cAMP responsive element binding protein,CREB)屬于DNA蛋白結(jié)合類亮氨酸拉鏈結(jié)構(gòu)轉(zhuǎn)錄因子超家族,主要與cAMP等信號發(fā)生應(yīng)答反應(yīng)[5]。由于mRNA具有穩(wěn)定性調(diào)節(jié)和翻譯后修飾的調(diào)控作用,機體細胞內(nèi)存在多種CREB基因的剪接方式,可以產(chǎn)生不同的剪接體,CREB1即為該家族中常見成員之一[6-7]。已有文獻報道,磷酸化CREB可以激活下游轉(zhuǎn)錄因子MITF,已知MITF是主控調(diào)節(jié)黑素細胞的發(fā)展、功能和生存的基因,可激活酪氨酸酶基因的轉(zhuǎn)錄[8-10]。基于此,本研究通過分子克隆等技術(shù)手段,獲得了獺兔CREB1基因的編碼區(qū)序列(coding sequence,CDS),并利用熒光定量手段檢測其在不同獺兔皮膚中的表達規(guī)律,為進一步探究CREB1基因的功能及毛色形成機制提供基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 試驗動物 選取成年青紫藍色獺兔、白色獺兔和黑色獺兔各3只,耳緣靜脈注射空氣處死,取約1 cm2相同位置的背部皮膚組織,-70 ℃保存。
1.1.2 試驗試劑 Gel Extraction Kit、DNA marker、dNTPs、TaqDNA聚合酶、反轉(zhuǎn)錄試劑盒、限制性內(nèi)切酶Hind Ⅲ、BamHⅠ、大腸桿菌感受態(tài)細胞DH5α、pMD19-T Simple Vector購于寶生物工程(大連)有限公司;Trizol試劑、質(zhì)粒提取試劑盒、SYBR?Green Master Mix試劑盒購于天根生化科技(北京)有限公司。
1.2 方法
1.2.1 總RNA的提取與檢測 采用Trizol法提取總RNA,通過1%變性瓊脂糖凝膠和NanoDrop 2000核酸蛋白測定儀,檢測總RNA的完整性、濃度和D260 nm/D280 nm。
1.2.2 引物設(shè)計 根據(jù)GenBank數(shù)據(jù)庫中提供的CREB1基因CDS序列(XM_002712603)設(shè)計特異性克隆引物,并分別在上下游引物中引入Hind Ⅲ和BamHⅠ酶切位點,同時設(shè)計兔CREB1、GAPDH基因?qū)崟r熒光定量PCR引物,由生工生物工程(上海)股份有限公司合成(表1)。
1.2.3CREB1基因的克隆 將總RNA進行反轉(zhuǎn)錄反應(yīng),利用CREB1基因特異性引物擴增編碼區(qū)序列。50 μL PCR反應(yīng)體系:ddH2O 33.5 μL,10×buffer 5 μL,dNTP 5 μL,正向引物2 μL,反向引物2 μL,模版2 μL,TaqDNA聚合酶0.5 μL。PCR反應(yīng)條件:95 ℃預(yù)變性5 min;95 ℃變性35 s,60 ℃退火35 s,72 ℃延伸1 min,共35個循環(huán);72 ℃延伸 10 min,4 ℃保存。構(gòu)建pMD19-T-CREB1克隆質(zhì)粒,用Hind Ⅲ和BamHⅠ進行雙酶切鑒定,選取陽性質(zhì)粒送生工生物工程(上海)股份有限公司測序。

表1 CREB1基因克隆引物序列
注:限制性內(nèi)切酶位點即序列中下劃線的部分。
1.2.4 生物信息學(xué)分析 進行氨基酸序列同源性比對,利用軟件MEGA 5.1構(gòu)建系統(tǒng)進化樹;蛋白質(zhì)組成及疏水性分析:http://cn.expasy.org/tools/protparam.html/;蛋白質(zhì)信號肽預(yù)測:http://www.cbs.dtu.dk/services/SignalP/;蛋白質(zhì)跨膜區(qū)預(yù)測:http://www.cbs.dtu.dk/services/TMHMM-2.0/;蛋白質(zhì)亞細胞定位分析:http://psort.hgc.jp/form2.html/;用Hopfield神經(jīng)網(wǎng)絡(luò)預(yù)測CREB1蛋白的二級結(jié)構(gòu)(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html);用swiss-model(http://swissmodel.expasy.org/workspace/index.php)預(yù)測CREB1蛋白的三級結(jié)構(gòu)。
1.2.5 實時熒光定量PCR 根據(jù)熒光定量試劑盒說明書,分別對青紫藍色、白色和黑色獺兔總RNA通過反轉(zhuǎn)錄反應(yīng),進行qRT-PCR試驗。20 μL反應(yīng)體系:AceQTMqPCR SYBR?Green Master Mix 10 μL,正向引物0.4 μL,反向引物0.4 μL,ROX Reference Dye 2 0.4 μL,DNA模板2.0 μL,滅菌蒸餾水6.8 μL。反應(yīng)程序:95 ℃預(yù)變性5 min;95 ℃變性 10 s,60 ℃退火30 s,40個循環(huán)。融解曲線:95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。以白色獺兔為對照組,以GAPDH基因為內(nèi)參基因,采用2-ΔΔCT法計算CREB1基因的相對表達量,利用SPSS 22.0統(tǒng)計軟件進行差異顯著性分析。
2 結(jié)果與分析
2.1 獺兔CREB1基因編碼區(qū)的克隆
利用1%瓊脂糖凝膠電泳檢測獺兔背部皮膚總RNA,顯示28S、18S和5S 3條帶,說明RNA完整性較好。用NanoDrop 2000檢測發(fā)現(xiàn),總RNA無DNA和蛋白質(zhì)污染,符合RT-PCR要求。利用CREB1基因特異性引物進行擴增,經(jīng)瓊脂糖凝膠電泳檢測,其產(chǎn)物大小與預(yù)期(984 bp)相符,且條帶清晰、特異性好(圖1-A)。
利用TA克隆原理,將CREB1基因連接至pMD19-T Simple載體上,獲得克隆質(zhì)粒pMD19-T-CREB1,并采用限制性內(nèi)切酶Hind Ⅲ和BamHⅠ進行雙酶切鑒定,結(jié)果顯示大小約2 600、1 000 bp的2條帶(圖1-B),與預(yù)期結(jié)果(2 692、984 bp)相符,說明CREB1基因克隆成功。
2.2 獺兔CREB1基因的序列分析
經(jīng)比對,克隆序列與NCBI(美國國立生物技術(shù)信息中心)已知序列相似度為99.6%,將克隆序列提交至GenBank(登錄號為KJ139984)。CREB1基因CDS含984個核苷酸(51.63% A+T,48.37% C+G),共編碼327個氨基酸,相對分子量約為35.05 ku。利用ExPASy中的工具ProtParam分析得出,CREB1蛋白等電點為5.30,不穩(wěn)定性系數(shù)為61.13,為不穩(wěn)定性蛋白;CREB1蛋白預(yù)測GRAVY值為-0.443,為親水蛋白。通過SignalP 4.0和TMHMM預(yù)測發(fā)現(xiàn),該蛋白無信號肽、無跨膜區(qū),亞細胞預(yù)測顯示該蛋白主要定位于細胞核(表2),推測該蛋白為核內(nèi)可溶性蛋白。
用Hopfield神經(jīng)網(wǎng)絡(luò)和Swiss-model分別對CREB1蛋白的二級結(jié)構(gòu)和三級結(jié)構(gòu)進行預(yù)測。結(jié)果如圖2所示,CREB1蛋白二級結(jié)構(gòu)中含有33.33%α螺旋(h),44.95%無規(guī)卷曲(c),21.71%延伸鏈(e);三級結(jié)構(gòu)為1條螺旋鏈。
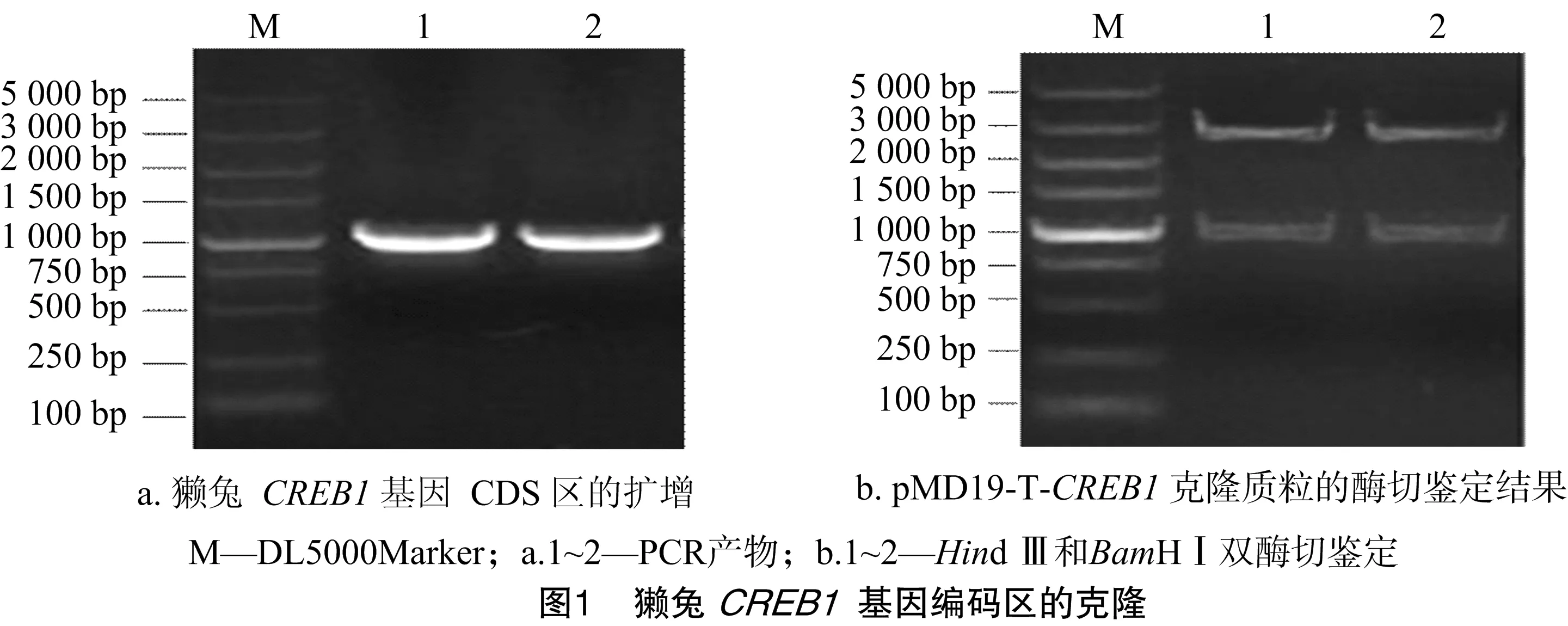

表2 CREB1蛋白質(zhì)的亞細胞定位分析
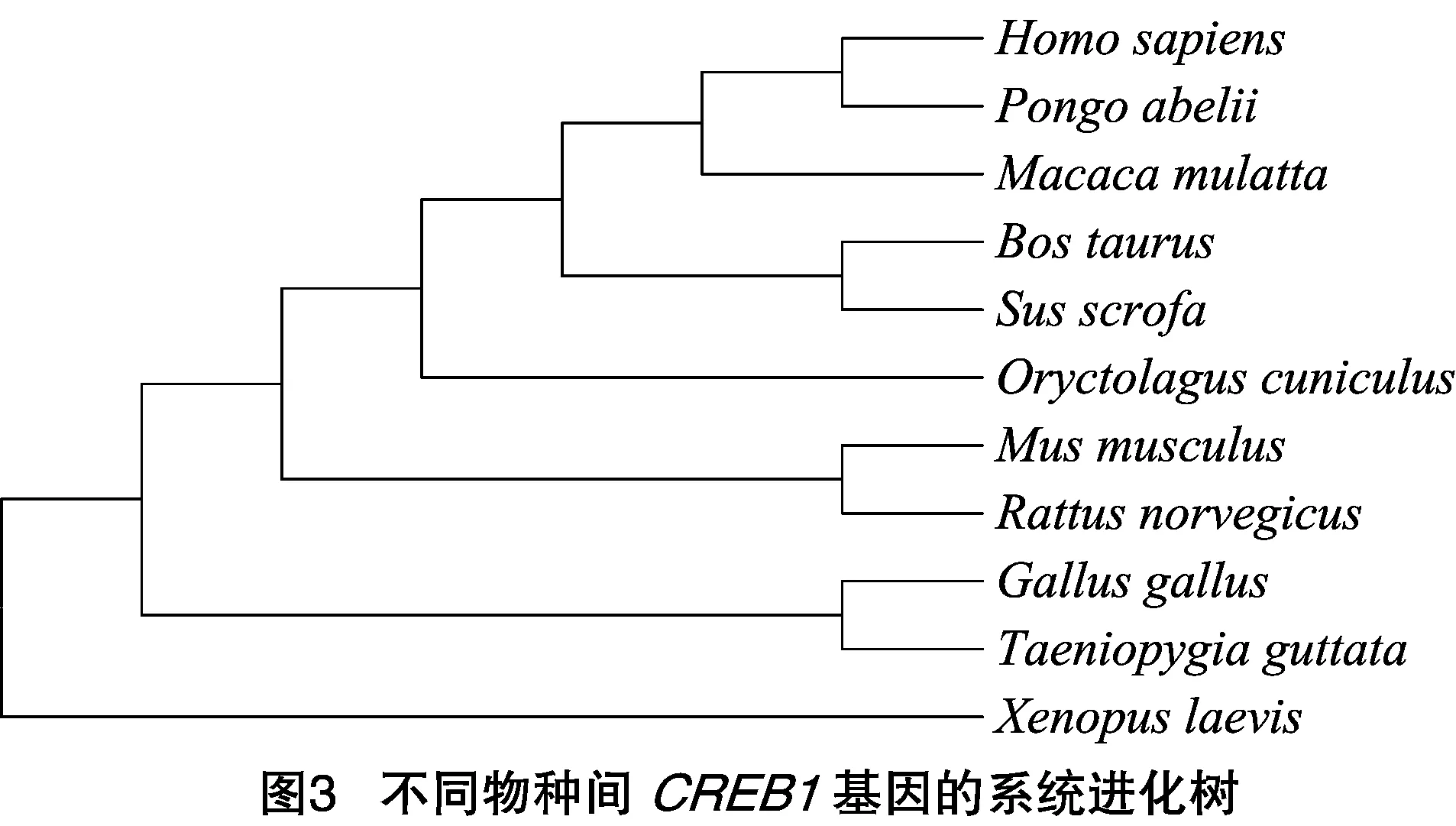
2.3 獺兔CREB1蛋白與其他物種的同源性比較及系統(tǒng)進化分析
將克隆測序得到的獺兔CREB1基因CDS與人(Homosapiens)、牛(BosTaurus)、豬(Susscrofa)、雞(Gallusgallus)、小鼠(Musmusculus)、獼猴(Macacamulatta)、猩猩(Pongoabelii)、大鼠(Rattusnorvegicus)、斑胸草雀(Taeniopygiaguttata)、非洲爪蟾(Xenopuslaevis)進行同源性比較發(fā)現(xiàn),獺兔CREB1基因氨基酸序列與人、豬等哺乳動物的同源性較高,為92.9%~95.4%,而與其他非哺乳動物(雞、斑胸草雀等)的同源性較低。利用MEGA 5.1軟件構(gòu)建鄰接法(NJ)系統(tǒng)進化樹,發(fā)現(xiàn)兔與人、猩猩、獼猴聚為1支,系統(tǒng)進化情況基本上與其親緣關(guān)系遠近一致,結(jié)果如圖3所示。

2.4 CREB1基因在不同毛色獺兔皮膚中的表達分析
通過qRT-PCR檢測黑色、白色和青紫藍色獺兔皮膚組織中CREB1基因mRNA的表達水平,結(jié)果發(fā)現(xiàn),CREB1基因在黑色獺兔和青紫藍色獺兔中的表達量均極顯著高于白色獺兔(P<0.01),且在黑色獺兔中的表達量最高(圖4)。

3 討論
cAMP反應(yīng)元件(cAMP response element,CRE)是指基因啟動子中的一段特定的DNA序列5′-TGACGTCA-3′[11]。CREB(環(huán)磷腺苷效應(yīng)元件結(jié)合蛋白)是PKA(蛋白激酶A)、MAPK(絲裂原激活的蛋白激酶)、PKC(蛋白激酶C)、CaMKs(鈣調(diào)蛋白激酶)等多種蛋白激酶的磷酸化底物,這些蛋白激酶可將CREB蛋白KID(激酶誘導(dǎo)區(qū))的Ser(絲氨酸)-133位點磷酸化,從而影響CREB的活性[12-15]。已知,cAMP-PKA信號通路介導(dǎo)的磷酸化反應(yīng)在黑色素生成過程中發(fā)揮了重要作用,CREB則是活化的PKA的主要靶標(biāo)蛋白,磷酸化后CREB激活MITF的轉(zhuǎn)錄,促使MITF與TYR基因啟動子Mbox結(jié)合,從而激活TYR的表達,促進黑色素生成。反之,降低CREB的磷酸化水平,MITF和酪氨酸酶也會受到一定的抑制[16]。在筆者所在課題組前期研究中,通過高通量技術(shù)手段發(fā)現(xiàn)CREB1基因與獺兔的毛色形成有關(guān),與已有報道相符。
在本研究中,筆者發(fā)現(xiàn)獺兔CREB1基因的氨基酸序列與NCBI中其他哺乳動物具有很高的同源性,即CREB1基因在哺乳動物中非常保守,那么CREB1基因在獺兔毛色形成中是否具有同樣的功能呢?已有報道指出,CREB1基因可調(diào)控MC1R基因與MITF基因之間色素沉著信號的傳遞,進而影響黑色素的沉積[17]。通過對黑色素生成信號通路的進一步研究發(fā)現(xiàn),MC1R對CREB1有激活作用,增強MITF基因的表達,活化了與黑色素形成密切相關(guān)的基因,如TYR、TYRP1和KIT,所以CREB1在黑色皮膚中的表達量較高[18]。本研究中獺兔CREB1基因在黑色獺兔和青紫藍色獺兔皮膚中表達水平高于白色獺兔,也印證了其他學(xué)者的觀點,CREB1基因?qū)谏氐男纬捎忻黠@的促進作用。
本研究利用分子克隆手段獲得獺兔CREB1基因的編碼區(qū)序列,通過生物信息學(xué)預(yù)測發(fā)現(xiàn),該蛋白為無信號肽、無跨膜區(qū)的核內(nèi)可溶性蛋白,且在哺乳動物中非常保守。同時發(fā)現(xiàn),CREB1基因的表達量與毛色的黑色素沉積存在一定的關(guān)聯(lián)性,本研究為探索獺兔毛色形成機制提供了一定的基礎(chǔ)資料。

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- 區(qū)域形象、區(qū)域文化認同與農(nóng)產(chǎn)品區(qū)域品牌購買行為
——情景因素的調(diào)節(jié)作用 - 農(nóng)業(yè)供給側(cè)結(jié)構(gòu)性調(diào)整內(nèi)生動能培育研究:基于共享農(nóng)業(yè)的視角
- 省部共建國家重點實驗室建設(shè)與運行思考
- 不同收入水平新型職業(yè)農(nóng)民培訓(xùn)需求差異
——以河北省為例 - 城鎮(zhèn)化建設(shè)質(zhì)量提升的科技訴求及供應(yīng)不足的成因與對策
- 超富集植物紫蘇活性炭的磷酸法制備與性能表征