接種不同AM真菌對滇重樓幼苗功能基因表達的影響
2019-03-08 08:46:52杜慧慧郭冬琴汪茂佳
天然產(chǎn)物研究與開發(fā) 2019年2期
張 華,杜慧慧,郭冬琴,楊 敏,汪茂佳,周 濃,*
1重慶三峽學(xué)院生物與食品工程學(xué)院 三峽庫區(qū)道地藥材綠色種植與深加工重慶市工程實驗室,重慶 404120;2大理大學(xué)藥學(xué)與化學(xué)學(xué)院,大理 671000
滇重樓(Parispolyphyllavar.yunnanensis)作為國家二級瀕危藥用植物,在我國西部藥用植物中占主要地位,其多以云南、四川、重慶、貴州、廣西等省市最為豐富[1-3]。隨著市場需求的不斷擴大,而滇重樓自然繁殖率較低、生長周期較長,以及人們的肆意亂挖亂采,嚴重破壞了其生存環(huán)境,使得滇重樓的野生資源蘊藏量急劇下降。
叢枝菌根(Arbuscular mycorrhizal,AM)真菌可以促進宿主對土壤營養(yǎng)物質(zhì)的吸收、促進植物移栽成活率、提高植物抗病能力、產(chǎn)量和品質(zhì)、調(diào)節(jié)植物種群和群落結(jié)構(gòu)以及維持生態(tài)系統(tǒng)穩(wěn)定性[4,5]。藥用植物的次生代謝產(chǎn)物是特定的基因型和特定的生境共同作用的產(chǎn)物[6]。重樓中含有豐富的三萜皂苷類等次級代謝化合物[7]。研究表明,鯊烯環(huán)氧酶基因(Squaleneepoxidase,SE)在調(diào)控碳流流向初級代謝或次級代謝中起到關(guān)鍵作用,是植物甾醇、三萜類化合物生物合成的關(guān)鍵酶[8],其微小的改變即可引起下游產(chǎn)物的大幅變化[9]。此外,AM 菌根共生早期信號途徑中涉及關(guān)鍵的基因包括共生受體樣蛋白激酶基因(Symbiosisreceptor-likekinase,SYMRK),產(chǎn)生鈣離子振蕩的通道蛋白基因(Doesn’tmakingfections1,DMI1) 和鈣/鈣調(diào)依賴性蛋白激酶基因(Calcium/calmodulin-dependentproteinkinase,CCaMK),這些基因所編碼的蛋白對于植物識別和應(yīng)答 AM 真菌早期信號途徑中是必需的[10-12]。與其他藥用植物相比,滇重樓的遺傳背景信息較少,研究AM菌根共生影響其相關(guān)產(chǎn)物基因表達對于獲取和利用藥用植物有效活性成分具有重要的意義。前期研究發(fā)現(xiàn),接種AM真菌能提高滇重樓根莖中甾體皂苷的含量[13]。因此本研究擬采用實時熒光定量PCR技術(shù)分析上述4種功能 基因的表達水平,研究不同AM真菌對這四個基因的表達效果的影響,以期尋找優(yōu)勢AM真菌,對于更好地保護和利用滇重樓資源具有理論和現(xiàn)實意義。
1 材料和方法
1.1 供試AM真菌
本試驗所用AM真菌通過美國國際叢枝菌根真菌種質(zhì)資源保藏中心(INVAM)購得相應(yīng)AM真菌純凈菌劑,接種菌劑為帶有孢子、菌絲的栽培基質(zhì)(見表1)。
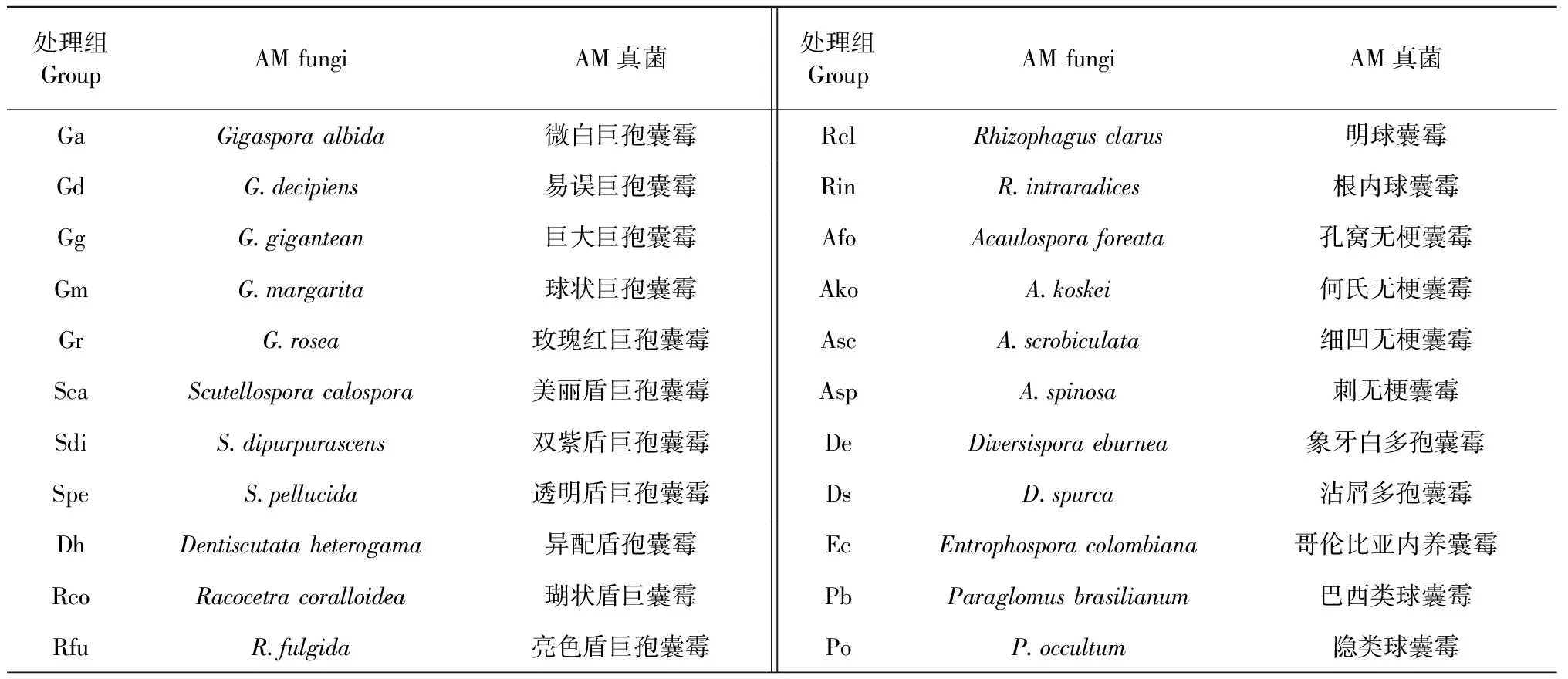
表1 不同處理組及其接種AM真菌
續(xù)表1(Continued Tab.1)

處理組GroupAM fungiAM真菌處理組GroupAM fungiAM真菌SdeSeptoglomus deserticola沙荒球囊霉AleAmbispora leptoticha薄壁兩性囊霉SviS.viscosum黏質(zhì)球囊霉AtrArchaeospora trappei崔氏原囊霉FmFunneliformis mosseae摩西球囊霉CKCK groups對照組CcClaroideoglomus claroideum近明球囊霉
1.2 滇重樓種子
滇重樓的新鮮種子于2012年10月18日采自大理州農(nóng)業(yè)科學(xué)推廣研究院種植基地的滇重樓健壯植株,常溫下河沙貯存4個月,并經(jīng)重慶三峽學(xué)院生物與食品工程學(xué)院周濃教授鑒定為百合科植物滇重樓P.polyphyllavar.yunnanensis的成熟種子,種子采用單株保存、保證種質(zhì)資源的穩(wěn)定性和均一性。
1.3 試劑與儀器
SGTriEx 高純總RNA提取試劑盒(# R1002),Thermo First cDNA Synthesis Kit(# Q1010),2×SG PCR MasterMix(# Q1009),2×SG Green qPCR Mix(with ROX)(# Q1002)(SinoGene公司,中國);DNase I,RNase-free(Fermentas公司,加拿大);3K15型冷凍離心機(Sigma公司,美國);JY-SPFT型電泳槽和JY300C型電泳儀和JY04S-3C型凝膠成像系統(tǒng)(北京君意東方電泳設(shè)備有限公司,中國);Biophotometer型分光光度計(Eppendorf公司,德國),StepOnePLUS型定量PCR(Applied Biosystems公司,美國)。
1.4 實驗設(shè)計
栽培基質(zhì)為重慶三峽學(xué)院百安校區(qū)的菜園土與河沙的混合物(體積比3∶1,過2 mm篩,121 ℃高壓滅菌鍋內(nèi)滅菌2 h)。采用室溫盆栽方法,設(shè)AM(接種28種AM真菌)組和CK(對照)組共29種處理。每處理6個重復(fù),每盆栽種滇重樓15株。將栽培袋用10%次氯酸鈉溶液消毒15 min后,并用流水清洗干凈。
1.5 總RNA提取、反轉(zhuǎn)錄cDNA
在同一接種框內(nèi),采用5點法取重樓1年生幼苗,每個點取5~10株混合為一個樣品,平行3份,快速洗凈泥土,用液氮速凍,存放于-80℃冰箱。按照SGTriEx高純總RNA提取試劑盒說明書提取29組樣品。利用1%瓊脂糖凝膠電泳和分光光度計檢測總RNA的濃度和純度。經(jīng)Thermo First cDNA Synthesis Kit反轉(zhuǎn)錄試劑盒反轉(zhuǎn)錄為cDNA后,以適量的 cDNA 為模板進行熒光定量PCR反應(yīng)。
1.6 實時熒光定量PCR
以適量的cDNA為模板,在熒光定量PCR儀(Applied Biosystems Step One PLUS)上分析滇重樓甾體皂苷合成關(guān)鍵酶:鯊烯環(huán)氧酶(SE);滇重樓與AM真菌共生相關(guān)基因:共生受體樣蛋白激酶(SYMRK),鈣/鈣調(diào)依賴性蛋白激酶(CCaMK)以及離子通道(ion channel)蛋白(DMI1)4個功能基因的相對表達量,以Actin為內(nèi)參,所用基因的引物見表2。采用比較Ct方法計算,每處理組重復(fù)3次。
熒光定量PCR的反應(yīng)體系為15 μL:2×SG Green qPCR Mix 7.5 μL,上下游引物各0.25 μL,cDNA 2 μL,nuclease-free water 6 μL。PCR反應(yīng)條件:95 ℃ 10 min,95 ℃ 20 s,60 ℃ 30 s,cycle 40,95 ℃ 15 s,60 ℃ 30 s,95 ℃ 15 s。

表2 所用基因的引物
1.7 數(shù)據(jù)處理
使用GraphPad Prism 6軟件作圖及進行多樣本間的單因素方差分析,P< 0.05 為顯著性差異,P< 0.01 為極顯著性差異。
2 結(jié)果與分析
2.1 滇重樓幼苗總RNA濃度與純度分析
所提取的28種AM真菌處理組和CK組滇重樓幼苗的總RNA經(jīng)1%瓊脂糖凝膠電泳和Biophotometer分光光度計檢測。完整的RNA是有28SrRNA和18SrRNA及5SrRNA 3條帶。如圖1所示,18SrRNA和28SrRNA的電泳條帶完整和清晰,含量大約2∶1,說明獲得的總RNA沒有發(fā)生降解。分光光度計檢測發(fā)現(xiàn),所有的總RNA的吸光光譜曲線平滑, OD260/OD280的比值在1.88~2.1之間,濃度最高為337.8 ng/μL。因此,所獲得的滇重樓幼苗的總RNA能夠用于后續(xù)的實驗。

圖1 不同AM真菌處理組和CK組滇重樓幼苗總RNA電泳圖Fig.1 The total RNA electrophoretogram of Paris polyphylla var.yunnanens is inoculated by different foreign arbuscular mycorrhizal fungi species
2.2 接種不同AM真菌 SE在滇重樓幼苗中表達差異
通過熒光定量PCR結(jié)果分析發(fā)現(xiàn),Po組,Ale組和Atr組能極顯著地提高SE基因在滇重樓幼苗中的表達量,其P<0.01(如圖2所示),而其余各組并不能提高該基因的表達量。鯊烯環(huán)氧酶(SE)的活性決定了環(huán)氧化鯊烯生物合成的效率,也會影響以環(huán)氧化鯊烯為前體的甾體皂苷等化合物的生物合成。根據(jù)本課題組前期研究發(fā)現(xiàn),滇重樓的幼苗根莖中以重樓皂苷Ⅶ含量最高,其次是重樓皂苷Ⅵ和重樓皂苷Ⅱ,重樓皂苷Ⅰ的含量最低;并且發(fā)現(xiàn)Po組,Ale組和Atr組的重樓皂苷Ⅶ和重樓皂苷Ⅵ均能高于CK組[14]。與本實驗的結(jié)果一致,Po組,Ale組和Atr組中SE基因的表達量與CK組呈顯著性上升。推測SE基因被Po,Ale和Atr誘導(dǎo)而高表達時重樓皂苷的含量也會增加。因此,不同的AM真菌能夠影響滇重樓幼苗中SE基因的表達差異,從而影響重樓皂苷成分的合成,最終影響滇重樓的品質(zhì)。

圖2 接種不同AM真菌 SE在滇重樓幼苗中的相對表達量Fig.2 The relative expression levels of SE in the rhizome of Paris polyphylla var.yunnanensis inoculated by different foreign arbuscular mycorrhizal fungi species注:數(shù)據(jù)經(jīng)獨立樣本的T檢驗*和**分別表示P<0.05和P<0.01的差異顯著水平,n=3,下同。Note:Date were analyzed by Independent-sample T test,*and ** indicate significant differences at P<0.05 and P<0.01,respectively,n=3.
2.3 接種不同AM真菌SYMRK在滇重樓幼苗中表達差異
在接種28株AM真菌處理后的滇重樓幼苗中,SYMRK基因的表達出現(xiàn)明顯的差異,其中除了Spe組,Ale組和Atr組能夠提高該基因在滇重樓幼苗的表達情況。Spe組中SYMRK基因的表達量顯著上升(P<0.05),Ale組和Atr組能誘導(dǎo)SYMRK基因高量表達,且呈極顯著的趨勢(P<0.01),而其余的AM真菌的處理組中均不能增加SYMRK基因的表達量,如圖3所示。前期研究發(fā)現(xiàn),Spe組、Ale組和Atr組能夠不同程度的侵染滇重樓并形成菌根。因此,加入Spe、Ale和Atr能夠增加滇重樓幼苗菌根生活力,并能夠與滇重樓幼苗根系形成良好的互惠共生的關(guān)系,促進其幼苗的生長發(fā)育。

圖3 接種不同AM真菌 SYMRK在滇重樓幼苗中的相對表達量Fig.3 The relative expression levels of SYMRK in the rhizome of Paris polyphylla var.yunnanensis inoculated by different foreign arbuscular mycorrhizal fungi species注:數(shù)據(jù)經(jīng)獨立樣本的T檢驗*和**分別表示P<0.05和P<0.01的差異顯著水平,n=3,下同。Note:Date were analyzed by Independent-sample T test,*and ** indicate significant differences at P<0.05 and P<0.01,respectively,n=3.
2.4 接種不同AM真菌CCaMK在滇重樓幼苗中表達差異
根據(jù)熒光定量PCR結(jié)果分析發(fā)現(xiàn),Ale組(P<0.05)和Atr組(P<0.01)能夠顯著性的提高CCaMK基因的表達量,而其余AM真菌組中該基因的表達量均出現(xiàn)不同程度的下降,如圖4所示。CCaMK基因可解碼Ca2+信號,從而引起共生體效應(yīng)因子的活性。在接種外源AM真菌的初級階段,真菌等微生物共生體能夠引起Ca2+的變化,從而引起叢枝菌根和共生體系侵染植物根部的細胞信號轉(zhuǎn)導(dǎo),引發(fā)一系列的生理變化。因此,CCaMK在這些過程中扮演著非常關(guān)鍵的角色,外源Ale和Atr可增加該基因的表達量,且能夠促進對滇重樓的侵染率,推測能夠在滇重樓的生長發(fā)育和品質(zhì)發(fā)揮重要的作用。
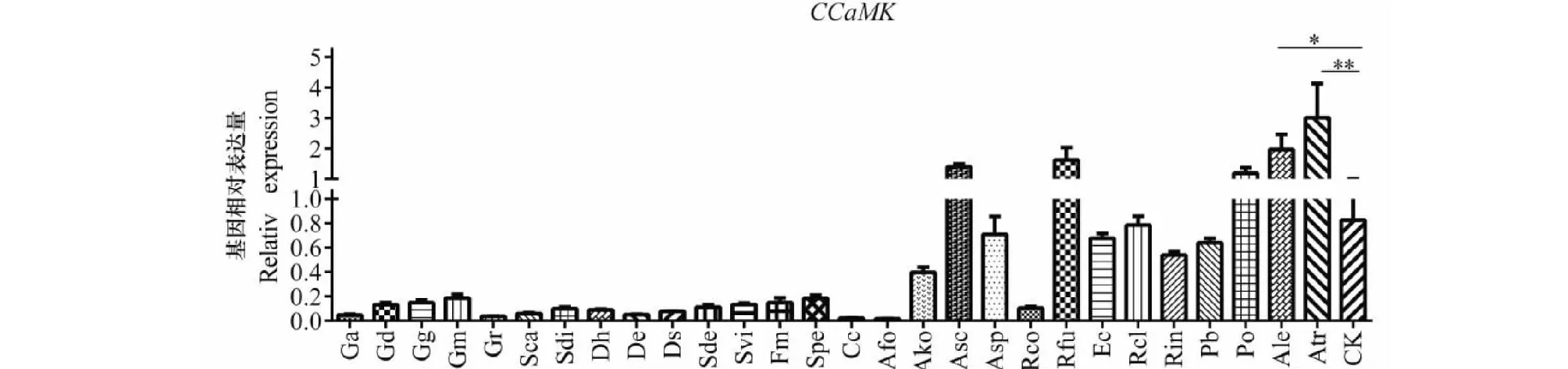
圖4 接種不同AM真菌 CCaMK在滇重樓幼苗中的相對表達量Fig.4 The relative expression levels of CCaMK in the rhizome of Paris polyphylla var.yunnanensis inoculated by different foreign arbuscular mycorrhizal fungi species注:數(shù)據(jù)經(jīng)獨立樣本的T檢驗*和**分別表示P<0.05和P<0.01的差異顯著水平,n=3,下同。Note:Date were analyzed by Independent-sample T test,*and ** indicate significant differences at P<0.05 and P<0.01,respectively,n=3.
2.5 接種不同AM真菌DMI1在滇重樓幼苗中表達差異
離子通道蛋白基因DMI1能夠編碼陽離子通道蛋白,調(diào)節(jié)細胞核內(nèi)外的離子流,對共生信號的傳導(dǎo)具有重要的作用。通過熒光定量PCR結(jié)果分析可知,Asc、Rfu、Ec、Rcl、Rin、Ale和Atr能夠顯著性的增加DMI1的表達量,且有些組能夠極顯著的增加(圖5所示)。說明接種不同AM真菌DMI1基因在滇重樓幼苗中表達增加,從而能夠介導(dǎo)植物與AM真菌和其他微生物之間的共生關(guān)系的建立。
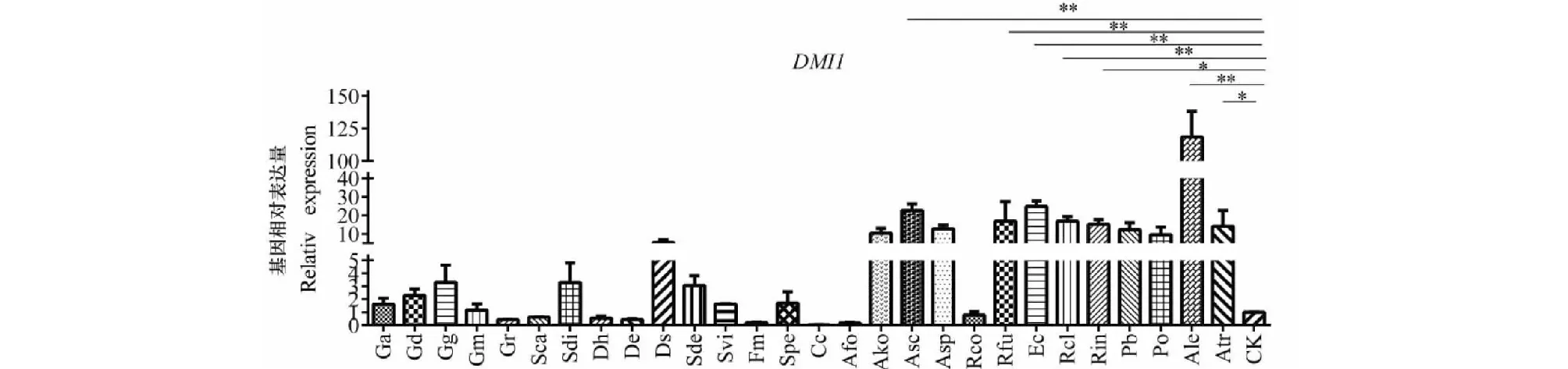
圖5 接種不同AM真菌 DMI1在滇重樓幼苗中的相對表達量Fig.5 The relative expression levels of DMI1 in the rhizome of Paris polyphylla var.yunnanensis inoculated by different foreign arbuscular mycorrhizal fungi species注:數(shù)據(jù)經(jīng)獨立樣本的T檢驗*和**分別表示P<0.05和P<0.01的差異顯著水平,n=3,下同。Note:Date were analyzed by Independent-sample T test,*and ** indicate significant differences at P<0.05 and P<0.01,respectively,n=3.
3 結(jié)論
野生重樓資源缺乏,重樓塊根生長緩慢,以及種植周期長和育苗存在問題等等,在很大程度上影響了重樓商品化種植和品質(zhì)的提高[15]。因此,篩選滇重樓優(yōu)勢菌株,提高滇重樓的產(chǎn)量是丞待解決的首要問題。AM真菌與植物之間具有一種普遍的共生現(xiàn)象,其中80%以上的陸生高等植物能與AM真菌形成共生體[16],同時作為重要的藥用植物根系與土壤菌根真菌的互惠共生體,可以顯著提高藥用植物的產(chǎn)量和品質(zhì)[17,18]。已有研究表明,多種AM真菌均可以促進滇重樓成年植株根莖次生代謝產(chǎn)物的形成,且不同AM真菌對滇重樓根莖中不同甾體皂苷的影響不盡相同[18,19]。重樓的主要有效成分是甾體皂苷,鯊烯環(huán)氧酶(SE)作為甲羥戊酸途徑的關(guān)鍵酶之一,該基因有助于調(diào)控活性物質(zhì)的合成從而提高產(chǎn)量。研究發(fā)現(xiàn)SE是控制人參皂苷的生物合成的關(guān)鍵酶之一[20],本研究發(fā)現(xiàn)接種外源AM真菌(Ale和Atr)共培養(yǎng)的滇重樓幼苗可以提高SE的表達量,從而可以促進滇重樓的品質(zhì)。
作為植物識別菌根真菌誘導(dǎo)而產(chǎn)生的特異分子的共生受體樣蛋白激酶(SYMRK)是控制共生形成的關(guān)鍵組分。AM真菌感染植物后會誘導(dǎo)共生基因的表達,而目前SYMRK在豆科植物中研究的較為清楚,但在非豆科植物尤其是藥用植物中的功能分析研究較少,本研究以接種不同AM真菌對SYMRK在滇重樓幼苗中的表達差異性研究,為該基因在藥用植物中的研究奠定了良好的理論基礎(chǔ)。
本研究主要分析4種功能基因的表達差異,為進一步研究重樓皂苷合成的代謝機制和利用次生代謝工程技術(shù)提高重樓的品質(zhì)奠定了分子基礎(chǔ)。后期可繼續(xù)研究4個功能基因在接種外源AM真菌與滇重樓幼苗共培養(yǎng)后的不同組織中的差異表達情況,盡可能闡明滇重樓有效成分合成及調(diào)控機制。根據(jù)本課題組前期研究發(fā)現(xiàn),接種本研究的28株AM真菌能夠使滇重樓幼苗的化學(xué)成分發(fā)生變化,能夠影響滇重樓幼苗菌根生活力、根莖生物量和重樓皂苷產(chǎn)量,結(jié)合本研究發(fā)現(xiàn)的Ale和Atr兩種真菌能夠顯著提高4種功能基因的表達量。綜上所述,建議滇重樓種子共生萌發(fā)時可接種薄壁兩性囊霉Ambisporaleptoticha(Ale)和崔氏原囊霉Archaeosporatrappei(Atr),進一步提高滇重樓幼苗的成活率和提高產(chǎn)量,為滇重樓的藥用價值奠定良好的基礎(chǔ)。結(jié)合后續(xù)優(yōu)勢菌株的篩選,并與之進行組合回接,以期能提高滇重樓的生長發(fā)育和在惡劣環(huán)境中的存活能力,從而為滇重樓育苗生產(chǎn)上帶來巨大的經(jīng)濟效益。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
