水稻粒形遺傳調控研究進展
2019-03-18 12:56:08曾智鋒楊維豐柯善文
安徽農業科學 2019年5期
尉 鑫,曾智鋒,楊維豐,韓 婧,柯善文*
(1.華南農業大學農學院,廣東廣州 510642;2.亞熱帶農業生物資源保護與利用國家重點實驗室,廣東廣州 510642)
水稻(OryzasativaL.)是重要的糧食作物,全球50 %以上的人口以稻米為主食[1]。隨著人類活動的影響以及工業化生產的加劇,有效耕地面積持續下降,近年來世界水稻的產量一度停滯不前[2]。為了解決糧食供應不足的問題,提高水稻單產已經成為糧食總產量增加的重要途徑。水稻產量主要由3個重要因素決定:千粒重、有效穗數和穗粒數。其中,有效穗數由植株的分蘗能力決定,穗粒數主要取決于枝梗數和結實率,而千粒重則受籽粒的形狀、大小以及灌漿飽滿度的影響[3]。
稻米的長度、寬度和長寬比影響水稻的粒形,而粒形是稻米品質鑒定的重要組成部分。中國優質稻谷標準規定:長寬比不低于2.8的列為優質秈稻品種。育種家在選育優質稻米品種過程中將谷粒形狀作為重要指標之一[4]。多數QTL(quantitative trait loci)的分析結果表明,水稻粒形的遺傳受多基因控制,且存在一因多效的現象,不同粒形性狀間有不同程度的相關性[5-9]。筆者從水稻粒形相關基因的克隆及功能分析方面綜述了前人的研究成果并展望研究前景,以期為水稻粒形基因克隆及高產育種提供理論指導。
1 水稻籽粒的結構
水稻加工成的消費產品主要來源于籽粒,籽粒中主要包含淀粉等營養成分,這就為人體的健康飲食提供了良好的材料。通常植物種子主要由3部分組成:胚、胚乳和種皮。與其他谷類作物一樣,水稻種子在結構上明顯不同于雙子葉植物的種子,例如擬南芥(圖1)。擬南芥種子中的胚占據了大部分的空間,其主要營養物質儲存于子葉中,而水稻種子中的大部分空間被胚乳占據,并且胚乳中儲存了大量的淀粉、少量的蛋白質和脂質以及其他微量物質(圖1),因此水稻的胚乳部分是人們主要的采食部位。水稻的粒形可以影響胚乳部分的大小并決定其重量,而其重量與粒重直接相關,所以水稻粒形是決定水稻產量的重要因素。

圖1 水稻和擬南芥成熟種子的縱切面[10]Fig.1 Longitudinal sections of mature seeds of Arabidopsis and rice [10]
2 水稻粒形相關基因的克隆
圖位克隆技術是挖掘功能基因的重要技術,該方法首先利用表型上差異較明顯的材料進行雜交處理來構建初級定位群體,例如F2群體、RIL群體等;然后采用分子標記篩選并結合表型分析的操作來發現效應值較大的QTLs,并且在此基礎上繼續發展新的作圖群體,進一步縮短目標區間的遺傳距離;最后對定位區間的候選基因進行分析與驗證,以此來確定目的基因的正確位置。
到目前為止,研究學者們已經克隆了一些直接或間接與水稻粒形調控相關的基因(表1)。其中僅有少數幾個基因對水稻的粒形起主要調控作用,其他的大部分基因均表現出一因多效的現象,它們是水稻粒形調控的微效基因,這些微效基因對水稻其他組織器官發育的影響更為顯著。
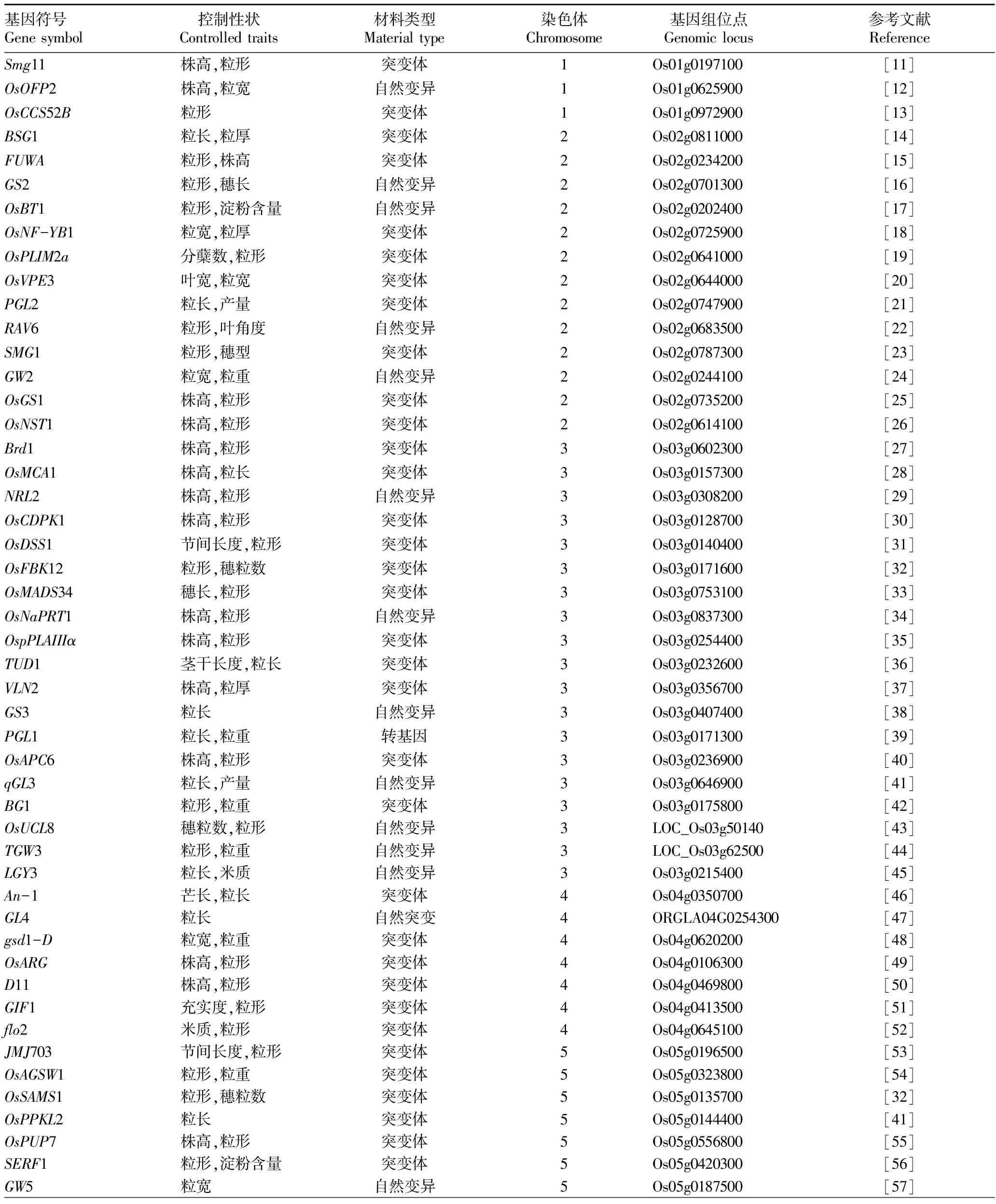
表1 已克隆的水稻粒形相關基因

接下表
注:數據主要來源于國家水稻數據中心,基因數據庫,http:∥www.ricedata.cn/gene/
Note:Data originated from China Rice Data Center and gene database,http:∥www.ricedata.cn/gene/
總結發現,93個已被克隆的水稻粒形相關基因分布于12對染色體上的數目各不相同,其中第3號染色體上的最多,第1、9、12號染色體上的較少(圖2)。經過多年來研究學者的努力,比較多的水稻粒形調控基因被成功克隆,但是與利用不同背景材料得到定位的粒形QTLs相比仍然很少,而且被克隆的粒形基因中大多數源自突變體材料,很難直接運用于生產實踐,所以基于自然變異條件下的基因定位、克隆對作物品種的改良和應用具有更加重要的意義。

圖2 已克隆水稻粒形基因在染色體上的分布Fig.2 Distribution of cloned grain shape genes on chromosomes in rice
3 水稻粒形主效基因的克隆
近年來,許多重要的水稻粒形基因都是利用栽培品種間表型差異較大的自然變異對目標QTL進行克隆而得到的。這類基因通常只對水稻粒形性狀具有顯著效應,并且不影響植物體的正常形態和發育,例如GW2、GS2、GS3、qGL3、TGW3、GL4、GW5、GS5、TGW6、GW7、GLW7和GW8等。
調控水稻粒長的主效基因主要包括GS3、qGL3、GL4和GLW7。Fan等[38]利用大粒的明恢63與小粒的川7構建NIL群體,利用5 740個BC3F2個體中的1 384個隱性個體和11個分子標記,將GS3定位在第3號染色體上著絲粒附近的7.9 kb區間內。Zhang等[41]選用小粒的N643和大粒的N411進行雜交,通過分析BC2F3代的結果將qGL3范圍縮小到RM15548和RM3513標記之間,高精度連鎖分析將qGL3進一步定位到XJ39和XJ26之間的46.6 kb區間內。Wu等[47]將滲入系中大粒的GIL25和小粒的IRGC102305進行雜交,對F2代的186個單株進行檢測,將目標區域確定在第4號染色體上的RM3335和RM5608之間,最后利用后代純合的分離株系將GL4定位在M3和M4之間的5.9 kb區域內。Si等[74]利用小粒性狀和大粒性狀的381個不同水稻品種進行全基因組關聯分析(GWAS),成功克隆了一個位于第7號染色體上控制粒長的主效QTL-GLW7,其編碼植物特殊的轉錄因子OsSPL13。
調控水稻粒寬的主效基因主要包括GW2、GW5、GS5和GW8。Song等[24]利用小粒的FAZ1與大粒的WY3構建BC2F2群體,利用加密標記不斷地縮短定位區間,最后通過對BC3F4群體的檢測,將GW2定位在第2號染色體的標記W024和W004之間的8.2 kb區間內。Shomura等[97]利用窄粒Kasalath和寬粒日本晴雜交自交產生的F2代分離群體進行QTL定位分析從而將qsw5精細定位于第5號染色體短臂端的2 263 bp區間內,但是Liu等[57]重新將GW5定位于第5號染色體短臂端,編碼鈣調素結合蛋白。Li等[59]利用珍汕97和H94構建DH群體,與此同時多次回交珍汕97發展作圖群體,從而將GS5定位在第5號染色體上短臂端的11.6 kb區間內。Wang等[83]利用華粳秈74(受體親本)和Basmati385、Basmati370(供體親本)構建的單片段代換系,鑒定出位于第8號染色體上長臂端的主效QTL-GW8,繼續進行精細定位以及高精度連鎖分析,將GW8定位在第8號染色體上的7.5 kb區間內。
調控水稻粒形長寬比的主效基因主要包括GS2、TGW3和GW7。Hu等[16]利用小粒Zhonghua11和大粒Baodali構建分離群體,將GS2定位在第2號染色體上的7.4 kb區間內。Ying等[44]將秈稻小粒品種黃華占(HHZ)和粳稻大粒品種JZ1560進行雜交構建RIL群體,利用SLAF-seq測序技術構建遺傳圖譜并進行QTL定位分析,成功檢測到一個位于第3號染色體上控制籽粒大小和千粒重的主效QTL-TGW3。Wang等[72]利用TaifengA(TFA)和HJX74構建作圖群體,最終將GW7基因準確定位于第7號染色體的2.6 kb區間內,其表達量上調可導致細長谷粒的形成。
綜上所述,水稻粒形主效基因的克隆主要涉及粒長、粒寬和長寬比3個方面,但是定位方法有所不同,大致分為3種:第一種,利用表型差異較大的雙親品種構建F2代分離群體、高回交世代群體、DH群體、NIL群體和RIL群體等作圖群體進行基因克隆;第二種,利用滲入系、CSSL和SSSL排除遺傳背景干擾,針對目標性狀進行基因克隆;第三種,利用測序技術,例如GWAS,針對某一性狀對大量不同品種材料進行測序分析,從而直接找到目標QTL,然后進行基因的克隆。就目前來看,利用測序分析技術進行基因克隆的方法與其他2種相比占有一定的優勢,不但節省了科研人員的工作量而且大大縮短了試驗時間。
4 水稻粒形基因間的互作
4.1GW2、GS3、GIF1和GW5/qSW5之間的互作Yan等[98]為了研究GS3、GW2、qSW5/GW5和GIF1這4個基因間的關系,對GS3-RNAi、GW2-RNAi系和qSW5的CSSL進行了基因表達分析。發現qSW5和GW2正向調控GS3的表達,GW2的表達能夠下調qSW5的表達,qSW5正向調控GIF1的表達,而GW2和GS3負向調控GIF1的表達;此外,利用一個含有180個水稻品種的自然群體,詳細分析了qSW5和GS3的等位基因作用,結果表明qSW5對粒長的作用受到GS3的影響,GS3對粒寬的作用受到qSW5的影響(圖3)。該研究為更加深入理解水稻籽粒大小發育的分子機制及提高水稻產量提供了有用的信息。

圖3 GS3、GW2、qSW5/GW5和GIF1之間的互作Fig.3 Interaction between GS3,GW2,qSW5/GW5 and GIF1
4.2qGL3/GL3.1、OsPPKL2和OsPPKL3之間的互作Zhang等[41]發現位于OsPPKL1中第2個Kelch功能域上保守的AVLDT區域的D364E的稀有等位變異qgl3導致長粒表型;OsPPKL1和OsPPKL3在水稻粒長調控中發揮負調節子的作用,而OsPPKL2是一個正調節子;Kelch功能域在OsPPKL1的負調節功能中是充分必要的(圖4)。田間試驗表明,優異等位基因qgl3的利用可以通過增加粒長、灌漿速率和粒重而顯著提高常規稻和雜交稻的產量。

圖4 OsPPKL1、OsPPKL2和OsPPKL3之間的互作Fig.4 Interaction between OsPPKL1,OsPPKL2 and OsPPKL3
4.3PGL1、PGL2和APG之間的互作Heang等[39]的研究表明一對拮抗作用的堿性螺旋-環-螺旋(bHLH)蛋白通過調控內外穎細胞長度,參與決定水稻籽粒長度。在內外穎過量表達PGL1,轉基因水稻籽粒長度和重量增加,是一種結合DNA的典型bHLH的抑制子。在內外穎過量表達PGL2同樣可以增加水稻粒長和粒重,并且這種增加與轉基因表達水平相關。PGL1的互作因子APG與其產生拮抗作用,是一個負調控因子。PGL1和APG在體內互作,都定位在核內,沉默APG的表型與過量表達PGL1一致,2個已知的籽粒長度相關基因GS3和SRS3的表達在PGL1過量表達株系和APG沉默植株中基本不受影響。PGL2與典型bHLH蛋白APG在體外和體內互作,與PGL1功能冗余,通過與APG形成異源二聚體抑制APG的功能,從而正向調節水稻籽粒長度。PGL1-APG代表了一種新的籽粒長度和重量調控的途徑(圖5)。

圖5 PGL1、APG、GS3和SRS3之間的互作Fig.5 Interaction between PGL1,APG,GS3 and SRS3
4.4其他粒形基因間的互作Wang等[72]研究發現,GW7編碼一種募集基蛋白(TONNEAU1),在細胞分裂的過程中調控橫向細胞分裂減少,縱向細胞分裂增加,過量表達可使籽粒變得更加細長。GW8(OsSPL16)是對粒寬具有調控作用的包含SBP結構域的轉錄因子,可以與GW7啟動子結合從而抑制其表達,并且OsSPL16-GW7模塊的相互作用可同時高稻米品質和稻米產量。與此同時,gs3和gw8的雙突變材料(以HJX74為背景的NIL-gw8gs3)表明谷粒細長的程度與親本相比呈現出累加作用,從而證明了GW8與GS3的調控途徑是相互獨立的。
Hu等[99]研究證明,TGW3編碼一種蛋白激酶OsGSK5/OsSK41,此蛋白激酶屬于GSK3/SHAGGY-Like家族。OsARF4是一種轉錄抑制因子,功能缺失可增加水稻粒長,在植物生長素途徑中發揮作用。兩者相互作用會使OsARF4磷酸化,而OsSK41和OsARF4的共同表達導致OsARF4的轉錄抑制功能增強。進一步的研究表明,GL3.3(TGW3)與GS3之間存在相互作用,其疊加效應可使水稻籽粒顯著增大,已經應用于大粒粳稻的品種選育中[100]。
5 水稻粒形調控的網絡途徑
在水稻和擬南芥中,調控種子粒形的信號途徑是相對保守的,均是通過影響胚乳和胚的生長發育來調控籽粒大小,并且是通過多種信號途徑共同作用的結果。將這些信號途徑總結歸納主要分為6種:IKU信號途徑、泛素-蛋白酶體途徑、G-蛋白信號途徑、MAPK信號途徑、植物激素途徑和轉錄調控因子途徑,其中植物激素途徑中主要包括的激素是油菜素內酯和生長素[101]。
這些途徑中所涉及到的已克隆基因可能通過大量不同的信號通路以及某些未知通路來參與調控細胞分裂和細胞增殖的過程。其中GW2和GW5/qSW5參與泛素-蛋白酶體途徑的調控;GS3和DEP1(qPE9-1)參與G-蛋白信號途徑的調控;OsMKK4(SMG1)和OsMAPK6參與MAPK信號途徑的調控;qGL3、CycT1;3和GS5參與植物激素途徑中的油菜素內酯調控途徑;BG1和TGW6參與植物激素途徑中的生長素調控途徑;而調控水稻粒形的大部分基因在轉錄調控因子途徑中發揮作用,例如GW6a(OsglHAT1)、GL7、GW8、GS2、SRS5等(圖6)。

圖6 水稻主要粒形基因的調控解析[101]Fig.6 The major signaling pathways of grain shape control in rice[101]
目前并沒有發現已克隆水稻粒形相關基因參與IKU信號途徑的調控,其主要存在于擬南芥中。IKU信號途徑與細胞分裂素和脫落酸信號相調控共同參與擬南芥種子結構中胚乳的形成,起到了正向調控的作用[101]。對胚乳形成具有反向調控作用的TGW6是一個控制水稻粒長和籽粒充實度的基因,其調控決定了水稻籽粒最終形態的形成[68],與IKU信號途徑一樣直接作用于種子胚乳的形成過程。
TGW3、LGY3、GL4、GSN1、WTG1和GS9為最新發現的水稻粒形主效調控基因,其中LGY3作用于G-蛋白信號途徑[45],GSN1作用于MAPK信號途徑[60],WTG1作用于泛素-蛋白酶體途徑[81],GS9作用于油菜素內酯調控途徑[85],而TGW3和GL4的作用途徑尚不明確[44,47]。但毋庸置疑的是水稻粒形基因調控網絡已初步被闡明。
6 小結與展望
由于世界人口的不斷增加,耕地面積的不斷減少,作為主要糧食作物的水稻產量問題一直受到人們的高度關注,而水稻粒形又與水稻產量有著非常密切的聯系,因此水稻粒形基因的克隆已經成為水稻產量和米質性狀研究的熱點之一。然而水稻自身遺傳背景的復雜性為水稻粒形的研究增加了困難,選擇粒形差異較大或者遺傳遠緣的品種材料會起到至關重要的作用。
經過多年來研究學者的不斷努力,已經有很多水稻粒形相關基因被成功克隆。筆者歸納了93個已克隆的與水稻粒形相關的基因,并且對多基因間的相互作用進行了詳細介紹,同時揭示了水稻粒形調控網絡。通過總結發現,一些基因既能獨立對水稻粒形性狀產生調控作用,也會相互聯系共同存在于同一個信號調控網絡,以此來實現控制細胞分裂和細胞伸長的功能,其中許多基因在育種選擇過程中顯著提高了水稻產量。這些發現既為科研人員針對水稻粒形性狀的分子調控機制的進一步研究提供了理論基礎,也為作物遺傳育種的基因改良工作創造了有利的條件。
如今關于水稻粒厚方面的研究報道比較少,而控制水稻粒形的基因是緊密聯系、相互作用的,粒厚基因的遺傳同樣受主效基因的調控,其遺傳研究與粒長、粒寬等相比較還不夠成熟[102]。利用新開發的基因組編輯技術CRISPR/Cas9在相同的遺傳背景下生成已知水稻粒形基因的突變體,將有助于建立控制水稻籽粒大小的遺傳調控網絡[101]。值得注意的是,相同的等位基因突變可能會導致不同遺傳背景下水稻的不同表型,研究為何相同的突變在不同品種上有不同的效果,將有助于育種工作者合理利用等位基因突變來提高特定遺傳背景下的水稻產量。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中等數學(2022年2期)2022-06-05 07:10:50
軍事文摘(2021年22期)2021-11-26 00:43:51
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
