新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌多樣性研究
2019-08-20 14:58:33方雷陳根元劉利林周小玲
江蘇農(nóng)業(yè)科學(xué) 2019年8期
方雷 陳根元 劉利林 周小玲
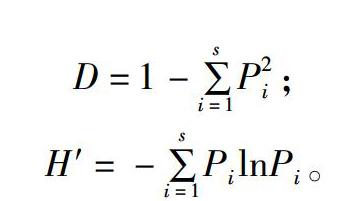

摘要:運用PCR-DGGE技術(shù)研究新疆驢盲腸、腹結(jié)腸、背結(jié)腸中固相食糜細(xì)菌組成多樣性。選用3頭成年、健康新疆驢,自由采食90%稻草+10%精料混合日糧,舍飼30 d后屠宰、分段取樣備用。經(jīng)過微生物基因組提取、目標(biāo)片段擴增、變性梯度凝膠電泳、圖像分析和數(shù)據(jù)處理,結(jié)果表明,不同腸道固相食糜細(xì)菌條帶組成上有相似區(qū)域,也有各自的特異條帶,而同一腸道段的個體樣本間的細(xì)菌條帶具有較高相似性,且盲腸固相食糜細(xì)菌的Shannon-Wiener指數(shù)、Simpson指數(shù)最大,與腹結(jié)腸差異不顯著,但都分別顯著高于背結(jié)腸。新疆驢盲腸固相食糜細(xì)菌多樣性最為豐富,腹結(jié)腸次之,背結(jié)腸相對最弱。
關(guān)鍵詞:新疆驢;盲腸;腹結(jié)腸;背結(jié)腸;固相食糜;細(xì)菌多樣性
中圖分類號: S822.1文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2019)08-0176-03
驢屬于單胃草食動物,與豬禽等單胃動物不同,具有發(fā)達(dá)的盲腸和結(jié)腸,這是驢重要的消化器官。腸道內(nèi)長期寄居著種類豐富、數(shù)量龐大的微生物,彼此間協(xié)同作用發(fā)酵、降解消化道食糜,產(chǎn)生揮發(fā)性脂肪酸,為宿主提供養(yǎng)分。有關(guān)研究結(jié)果表明,驢對稻草、麥秸、玉米秸稈日糧中的中性洗滌纖維(NDF)消化率為40%~50%,與牛羊等反芻動物對這3種秸稈日糧的NDF消化能力相當(dāng),說明驢的盲腸、結(jié)腸的腸道微生物對秸稈粗飼料有較強的降解能力[1],其功能類似于反芻動物的瘤胃。反芻動物的瘤胃微生物主要黏附在飼料顆粒上、存在于瘤胃液中和附著于瘤胃內(nèi)壁上[2],且瘤胃固相食糜中的微生物數(shù)量高于液相,是瘤胃中的主要功能微生物[3-5]。瘤胃微生物的多樣性受飼料組成[6]、宿主遺傳背景[7]和環(huán)境等因素影響[8];反芻動物的瘤胃、網(wǎng)胃、瓣胃和皺胃4個胃室中菌群多樣性也不一樣,呈現(xiàn)先高后低再升高的趨勢[9]。驢的盲腸、腹結(jié)腸、背結(jié)腸是日糧中纖維素、半纖維素等結(jié)構(gòu)性碳水化合物降解的重要場所,不同消化部位的固相食糜細(xì)菌多樣性如何變化是本研究的切入點。
1材料與方法
1.1試驗動物、時間及地點
試驗于2017年6月5日至7月5日在塔里木大學(xué)動物科學(xué)學(xué)院試驗養(yǎng)殖基地進(jìn)行。試驗選用3頭成年、健康、3~4歲新疆驢,舍飼喂以含有90%粉碎稻草和10%精料(70%玉米粉+30%棉粕)的混合日糧,自由采食、飲水,飼養(yǎng)30 d。
1.2樣品處理
飼養(yǎng)結(jié)束后,屠宰前1 d晚上禁食、供水,屠宰前先用鈍器打擊頭部致暈,立即頸動脈放血、開膛、暴露內(nèi)臟;找到“逗號”狀盲腸,隨后向后繼續(xù)找到膨大腹結(jié)腸、膨大背結(jié)腸,用繩子扎緊各段間狹小接口,水平放置,從各腸道段中部取樣;分別采集盲腸、腹結(jié)腸、背結(jié)腸食糜,并用4層滅菌紗布過濾,將過濾后的固相食糜迅速收集于數(shù)個2 mL滅菌指形管中,立即投入液氮罐于實驗室,分類保存于-80 ℃冰箱備用。
1.3食糜微生物總DNA提取
參照Kang等[10]和Laure等[11]的方法洗脫固相食糜上附著的微生物,用天根生物技術(shù)有限公司微生物基因組DNA提取試劑盒(DP328)提取微生物基因組總DNA,具體步驟按說明書操作。
1.4目標(biāo)片段擴增及檢測
試驗采用細(xì)菌16S rDNA的V3區(qū)通用引物對目標(biāo)片段進(jìn)行擴增,上游引物HDA1-GC序列為:5′-[JP9]CGCCCGGGGCGCGCCCCGGGCGGGGCGGGGGCACGGGGGGACTCCTACGGGAGGCAGCAGT-3′,下游引物HDA2序列為:5′-[JP9]GTATTACCGCGGCTGCTGGCAC-3′。以上引物由上海生工生物技術(shù)有限公司合成,可擴增長約200 bp的目標(biāo)片段。
PCR反應(yīng)條件如下:95 ℃預(yù)變性5 min;95 ℃變性30 s,56 ℃退火30 s,72 ℃延伸50 s,共30個循環(huán)。PCR產(chǎn)物由 1.0% 瓊脂糖凝膠電泳檢測。
1.5變性梯度凝膠電泳
試驗中DGGE采用10%聚丙烯酰胺凝膠,丙烯酰胺和甲叉雙丙烯酰胺比例為37.5 ∶1,變性劑濃度梯度為45%~60%。電泳采用Dcode DGGE系統(tǒng)(Bio-Rad,美國),在 0.5×TAE緩沖液中、60 ℃恒溫下進(jìn)行,200 V預(yù)電泳10 min,85 V電泳16 h。電泳結(jié)束后,用0.5 μg/mL的EB溶液染色30 min,用凝膠成像系統(tǒng)(Gel DOCTM XR+,Bio-Rad美國)檢測、拍照。
1.6圖譜處理與數(shù)據(jù)分析
用Quantity One 4.62軟件對DGGE圖譜進(jìn)行分析,將各泳道條帶光密度值計算、輸出用于計算Simpson指數(shù)和Shannon-Wiener指數(shù)[12-13],采用SAS 8.0軟件對細(xì)菌多樣性指數(shù)進(jìn)行Duncans檢驗和統(tǒng)計分析,P<0.05表示差異顯著,P<0.01表示差異極顯著。用NTSYSpc 2.10S對圖譜條帶聚類分析。
D=1-∑si=1P2i;
H′=-∑si=1PilnPi。
式中:Pi為某樣品中某1條帶光密度值占該樣品中總光密度值的比率,D為Simpson指數(shù),H′為Shannon-Wiener指數(shù)。
2結(jié)果與分析
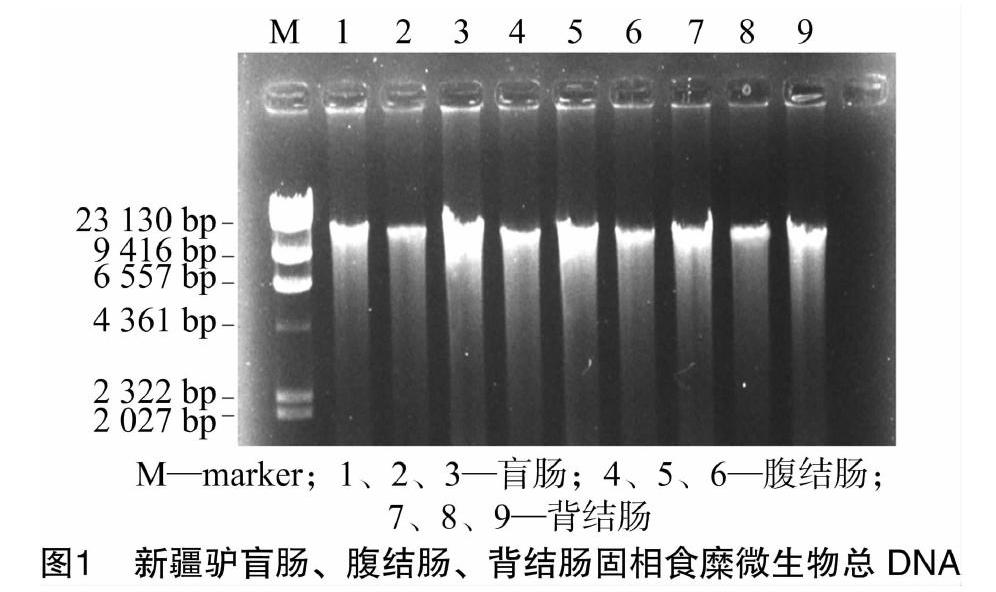
2.1新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜微生物總DNA
新疆驢不同段腸道固相食糜微生物總DNA提取結(jié)果見圖1。由圖1可見,盲腸、腹結(jié)腸、背結(jié)腸各代表樣品分別在 1~9 泳道均有明亮條帶,與marker相比,條帶大小在2.3 ku左右,有稍許拖尾,比較完整,D260 nm/D280 nm比值為1.71,可以用作后續(xù)PCR模板。
2.2新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌16S rDNA V3區(qū)PCR擴增產(chǎn)物
以新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜微生物總DNA為模板,細(xì)菌16S rDNA V3區(qū)通用引物進(jìn)行PCR擴增,經(jīng) 1.0% 瓊脂糖凝膠電泳、染色、拍照,結(jié)果表明,不同腸道固相食糜各樣本均獲得整齊、明亮的目標(biāo)PCR產(chǎn)物,大小為 200 bp(圖2),符合引物擴增預(yù)期產(chǎn)物結(jié)果,可以用于后續(xù)變性梯度凝膠電泳。
2.3新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌16S rDNA V3區(qū)PCR-DGGE指紋圖譜
新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌V3區(qū)PCR擴增產(chǎn)物經(jīng)45%~60%變性梯度凝膠電泳、染色、照相,結(jié)果表明,各腸道段各樣本在各自泳道均出現(xiàn)豐富、明亮、清晰條帶,同一腸道段樣本間有大量相似條帶,不同腸道間既有部分相似條帶,也有所屬腸道段的特異條帶(圖3)。
2.4新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌DGGE指紋圖譜聚類分析
新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌PCR-DGGE圖譜聚類分析結(jié)果見圖4。各段腸道固相食糜細(xì)菌條帶都以高于0.93的相似性分別聚合成盲腸(1、2、3)、腹結(jié)腸(4、5、6)和背結(jié)腸(7、8、9)三大類群,腹結(jié)腸和背結(jié)腸又以0.67左右的相似性聚合成大結(jié)腸類群,最后再以0.56相似性和盲腸細(xì)菌條帶聚合成一大簇。
2.5新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌多樣性指數(shù)分析
新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜細(xì)菌PCR-DGGE凝膠圖片經(jīng)Quantity One軟件對各泳道條帶光密度值輸出、多樣性指數(shù)計算和SPSS統(tǒng)計分析后,結(jié)果(表1)顯示,新疆驢盲腸固相食糜細(xì)菌的Shannon-Wiener指數(shù)和Simpson指數(shù)均為最大,分別為3.209和0.947,與腹結(jié)腸差異不顯著,但均分別極顯著高于背結(jié)腸(P<0.01)。
3討論與結(jié)論
反芻動物瘤胃微生物的存在方式,主要以黏附固相飼料顆粒、游離于瘤胃液中和附著于瘤胃內(nèi)壁上,其中固相食糜黏附微生物占瘤胃微生物總數(shù)的50%~70%[14],以大量纖維降解菌為主,纖維素降解也是瘤胃代謝的限速步驟[15]。通過表1 細(xì)菌多樣性指數(shù)來看,盲腸固相食糜的Shannon-Wiener和Simpson指數(shù)值都是最大,分別為3.209和0.957,與腹結(jié)腸的3.168和0.927相比差異不顯著,但都分別極顯著高于背結(jié)腸的2.950和0.915(P<0.01)。也就是說,盲腸固相食糜上附著的細(xì)菌種類更加豐富,腹結(jié)腸次之,背結(jié)腸相對最弱;瘤胃微生物在降解日糧過程中,主要包括附著和酶解2個過程,其中附著過程是瘤胃微生物降解日糧的前提[16-17],靠附著微生物間協(xié)同作用降解食糜;推測盲腸在對固相食糜顆粒降解的作用要強于腹結(jié)腸,強于背結(jié)腸;可能一方面盲腸是驢大腸微生物發(fā)酵降解食糜重要的起始段,胃、小腸等消化液消化后,食糜中不易被消化或逃脫的易消化養(yǎng)分首先來到盲腸進(jìn)行微生物降解,食糜中的養(yǎng)分組成和含量有利于盲腸微生物的生長繁殖,使其食糜附著細(xì)菌多樣性更好;另一方面,隨著盲腸、腹結(jié)腸和背結(jié)腸微生物對食糜的逐級降解,食糜中易被利用和能被利用的養(yǎng)分也越來越少,也導(dǎo)致了固相食糜上附著的細(xì)菌多樣性逐漸降低。
從新疆驢盲腸、腹結(jié)腸和背結(jié)腸固相食糜指紋圖譜(圖3)可以看出,在相同試驗條件下,不同消化道段都獲得了較豐富的細(xì)菌條帶,有共同、特異和很亮的優(yōu)勢細(xì)菌條帶,但試驗中均未回收、克隆和測序,無法獲知其分類信息;從聚類信息(圖4)上看,各不同腸道處理內(nèi)的細(xì)菌條帶組成都有0.9以上的相似系數(shù),而驢的不同腸道處理間只有0.56的相似系數(shù)。本試驗中,我們對新疆驢盲腸、腹結(jié)腸、背結(jié)腸固相食糜附著細(xì)菌多樣性有了初步認(rèn)識,這些不同消化道微生物差異表現(xiàn)在微生物什么分類水平上,是什么原因造成的仍需進(jìn)一步研究和探討。
新疆驢的相同腸道段固相食糜細(xì)菌組成具有較高的相似性,不同腸道段間細(xì)菌組成有所差異;盲腸固相食糜的細(xì)菌多樣性最為豐富,腹結(jié)腸次之,背結(jié)腸相對最弱。
參考文獻(xiàn):
[1]方雷,礦理揚,牛志濤. 新疆驢對4種秸稈日糧采食與消化的研究[J]. 新疆農(nóng)業(yè)大學(xué)學(xué)報,2009,32(3):45-48.
[2]Mcallister T A,Bae H D,Jones G A,et al. Microbial attachment and feed digestion in the rumen[J]. J Ainm Sci,1994,72:3004-3018.
[3]馮薇,王加啟,劉開朗,等. 運用PCR-DGGE分析比較瘤胃中不同飼料固相黏附微生物區(qū)系[J]. 畜牧獸醫(yī)學(xué)報,2010,41(12):1556-1562.
[4]Peter R M,F(xiàn)orster R J,Yang W,et al. Characterization of rumen bacterial diversity and fermentation parameters in concentrate fed cattle with and without forage[J]. Journal of Applied Microbiology,2012,112(6):1152-1162.
[5]Huws S A,Lee M R,Muetzel S M,et al. Forage type and fish oil cause shifts in rumen bacterial diversity[J]. FEMS Microbiology Ecology,2010,73(2):396-407.
[6]吳小燕,彭全輝,王之盛,等. 不同蛋白質(zhì)飼料對宣漢黃牛瘤胃固相黏附纖維降解微生物數(shù)量的影響[J]. 動物營養(yǎng)學(xué)報,2014,26(11):3298-3306.
[7]Benson A K,Kelly S A,Legge R,et al. Individuality in gut microbiota com position is a complex polygenic trait shaped by multiple environmental and host genetic factors[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(44):18933-18938.
[8]Uyeno Y,Sekiguchi Y,Tajima K,et al. An r-RNA-based analysis for evaluating the effect of heat stress on the rumen microbial com position of Holstein heifers[J]. Anaerobe,2010,16(1):27-33.
[9]曾燕,曾東,倪學(xué)勤,等. 應(yīng)用PCR-DGGE技術(shù)比較綿羊瘤胃、網(wǎng)胃、瓣胃和皺胃菌群的多樣性[J]. 動物營養(yǎng)學(xué)報,2015,27(1):298-304.
[10]Kang S H,Denman S E,Morrison M I. An efficient RNA extraction method for estimating gut microbial diversity by polymerase chain reaction[J]. Current Microbiology,2009,58(5):464-471.
[11]Larue R,Yu Z T,Parisi V A,et al. Novel microbial diversity adherent to plant biomass in the herbivore gastrointestinal tract,as revealed by ribosomal intergenic spacer analysis and rrs gene sequencing[J]. Environmental Microbiology,2005,7(4):530-543.
[12]崔振亮,孟慶翔,吳浩,等. PCR-DGGE技術(shù)研究青貯桑葉對肉牛瘤胃細(xì)菌區(qū)系的影響[J]. 飼料研究,2011(1):1-4.
[13]許晴,張放,許中旗,等. Simpson指數(shù)和Shannon-Wiener指數(shù)若干特征的分析及“稀釋效應(yīng)”[J]. 草業(yè)科學(xué),2011,28(4):527-531.
[14]Yang W,Beauchemin K,Rode L. Effect of dietary factors on distribution and chemical composition of liquid-or solid-associated bacterial populations in the rumen of dairy cow[J]. Journal of Animal Science,2001,79(10):2736-2746.
[15]Koike S,Yoshitani S,Kobayashi Y,et al. Phylogenetic analysis of fiber-associated rumen bacterial community and PCR detection of uncultured bacterial[J]. FEEMS Microbiol Lett,2003,229(1):23-30.
[16]Michalet-Doreau B,F(xiàn)ernandez I,F(xiàn)onty G. A comparison of enzymatic and molecular approaches to characterize the cellulolytic microbial ecosystems of the rumen and the cecum[J]. Journal of Animal Science,2002,80(3):790-796.
[17]Merry R,Mcallan A. A comparison of the chemical composition of mixed bacteria harvested from the liquid and solid fractions of rumen digest[J]. British Journal of Nutrition,1983,50(03):701-709.
