海藻糖和甘露醇對凍融循環引起的蝦蛄肌原纖維蛋白結構和功能特性變化的影響
2019-09-06 07:52:42陳金玉何麗麗梁喚喚韋英霞韓業勤張坤生
食品科學 2019年16期
關鍵詞:甘露醇
陳金玉,李 彬,何麗麗,梁喚喚,韋英霞,韓業勤,張坤生*
(天津市食品生物技術重點實驗室,天津商業大學生物技術與食品科學學院,天津 300134)
冷凍貯藏是原料肉和肉制品保鮮貯藏的主要方式。在低溫條件下微生物和酶活性受到抑制,但是肌肉品質的劣變仍然無法避免。尤其目前我國冷鏈技術尚不完善,在肉品的長途運輸、貯藏及消費過程中,由于溫度波動不可避免地出現反復凍融現象。肉在反復凍融過程中會發生肉質硬化、柔軟性下降、持水力降低、組織纖維化等變化,產生這些現象的根本原因是肌原纖維蛋白的變性[1]。肌原纖維蛋白是由肌球蛋白、肌動蛋白、原肌球蛋白、肌鈣蛋白、輔肌動蛋白等所組成的鹽溶性蛋白。反復凍融使樣品中小冰晶體重結晶,從而在肌原纖維中重新分布,導致細胞結構被破壞,肌原纖維蛋白變性,降低肉產品的食用價值[2]。因此研究反復凍融對肌肉品質的影響以及控制肉蛋白冷凍變性具有重要的現實意義。
冷凍變性保護劑又稱抗凍劑,是目前防止蛋白冷凍變性的最主要方法。保護劑的種類很多,蔗糖因其價格低廉、抗凍效果較好且不易發生美拉德反應而廣泛應用于生產中,但此類抗凍劑甜度和熱量相對較高,與當前人們對健康飲食的追求不太相符,因此低甜度、低熱量的抗凍劑成為新的研究對象。目前海藻糖和甘露醇作為低熱量功能性糖醇類抗凍劑受到越來越多關注。Sei等[3]發現海藻糖在抑制冰晶增長和冰晶形態不穩定方面,比蔗糖更具優勢。薛勇等[4]采用10%海藻糖作為抗凍劑,測定凍藏后鳙魚肌原纖維蛋白性質的變化,結果表明海藻糖的抗冷凍效果優于蔗糖和山梨醇混合物。然而,目前關于海藻糖和甘露醇復配對反復凍融引起的肌原纖維蛋白氧化變性的影響還鮮有報道。
在眾多肉類食品中,蝦蛄(Oratosquilla oratoria)作為一種營養豐富、肉質鮮美的海產品,深受消費者喜愛。蝦蛄收獲的季節性非常強,為便于保鮮,常采用低溫貯存。在貯藏加工過程中,蝦蛄肉品質的變化直接影響其消費價值,其中肌原纖維蛋白對蝦蛄產品的保水性、質地等品質都起著決定性作用。因此,本實驗以蝦蛄為對象,研究反復凍融處理對肌原纖維蛋白氧化、結構和功能特性的影響,探索凍融循環對蝦蛄蛋白品質的影響規律。在此基礎上,探討4 g/100 mL海藻糖+4 g/100 mL甘露醇抗凍劑對蝦蛄肌原纖維蛋白的冷凍保護作用和機制。
1 材料與方法
1.1 材料與試劑
實驗所用口蝦蛄捕自渤海灣(天津)近海海域,購于天津市北辰區老板娘水產城。
海藻糖、甘露醇、2,4-二硝基苯肼(2,4-dinitrophenyl hydrazine,DNPH)、5,5’-二硫代雙(2-硝基苯甲酸)(5,5’-dithiobis-(2-nitrobenzoic acid),DTNB) 美國Sigma公司;其他試劑均為分析純 上海阿拉丁試劑有限公司。
1.2 儀器與設備
Avanti J-E高效冷凍離心機 美國Beckman公司;U5100紫外-可見分光光度計、F-4600熒光分光光度儀、SU3500掃描電子顯微鏡 日本Hitachi公司;Physica MCR301高級旋轉流變儀 奧地利Anton Paar公司;TA.XT Plus質構儀 英國Stable Micro System公司;POLYTRON PT 2100勻漿機 瑞士Kinematica公司。
1.3 方法
1.3.1 凍融循環處理
蝦蛄去頭、尾,剝殼,取肉剁碎。將蝦蛄肉糜分為兩份,每份1 000 g,一份為對照組,一份為實驗組。將實驗組肉糜浸泡在冷凍保護劑溶液中2 h,然后將瀝干的肉糜樣品(約0.5 cm×0.5 cm×0.5 cm)放入聚乙烯自封袋中(5 份,每份200 g,分別編號為實驗1、2、3、4、5組)。對照組做同樣的分組處理。將對照組和實驗組樣品置于-20 ℃冰箱中冷凍,5 d后取出,室溫解凍至其中心溫度為0 ℃,然后放回-20 ℃冰箱中貯藏,此為一個凍融處理。從5 組樣品中隨機抽取3 份進行各項指標的測定,然后將剩余樣品放回-20 ℃繼續冷凍5 d。依此方法分別進行0、1、3、5 次凍融處理,并進行相應的指標測定。考慮到目前工業上采用的商業抗凍劑為4%蔗糖+4%山梨糖醇,因此本實驗選擇4 g/100 mL海藻糖+4 g/100 mL甘露醇為冷凍保護劑溶液。
1.3.2 肌原纖維蛋白提取
參考Park等[5]的方法。向肉糜中加入4 倍體積蛋白提取液(0.1 mol/L NaCl、0.002 mol/L MgCl2、0.001 mol/L EDTA-2Na、0.1 mol/L磷酸鹽緩沖液,pH 7.0),11 000 r/min勻漿25 s,4 ℃、5 000 r/min離心15 min,棄上清液,留沉淀,再加入蛋白提取液重復上述操作2 次得粗蛋白。將所得粗蛋白與4 倍體積0.1 mol/L NaCl溶液混合,勻漿25 s,4 ℃、5 000 r/min離心15 min,棄上清液,留沉淀,再加入NaCl溶液重復上述操作2 次即得蝦蛄肌原纖維蛋白。采用雙縮脲法測定蛋白質濃度,以牛血清蛋白為標準品。
1.3.3 凍融循環對蝦蛄肌原纖維蛋白氧化及結構的影響
1.3.3.1 羰基含量測定
采用DNPH法測定蛋白羰基含量[6]。取0.5 mL蛋白溶液(5 mg/mL)于離心管中,加入1 mL 10 mmol/L DNPH(溶于2 mol/L鹽酸),室溫避光反應1 h,每隔10 min渦旋一次,然后加入等體積20%三氯乙酸溶液,5 000 r/min離心10 min,棄上清液,所得沉淀用1 mL乙醇-乙酸乙酯(1∶1,V/V)充分洗滌3 次,以除去未反應的DNPH。加入4 mL鹽酸胍溶液(6 mol/L,溶于20 mmol/L磷酸鹽緩沖液,pH 6.5),37 ℃溶解沉淀20 min,10 000 r/min離心5 min,取上清液于370 nm波長比色。羰基含量根據摩爾消光系數22 000 L/(mol?cm)計算。空白對照組加入1 mL 2 mol/L鹽酸(不含DNPH),其余步驟同上。
1.3.3.2 總巰基含量測定
蛋白巰基含量通過DTNB法測定[7]。肌原纖維蛋白(5 mg/mL)溶解于含5%十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)的Tris緩沖液(0.1 mol/L,pH 8.0),80 ℃水浴45 min,8 000 r/min離心10 min。收集上清液用于巰基含量的測定。用相同緩沖液配制10 mmol/L DTNB。取0.5 mL上清液與2 mL 5% SDS/Tris緩沖液混合,隨后加入0.2 mL DTNB避光反應30 min,混合液于波長412 nm測定吸光度。空白以5% SDS/Tris緩沖液替代蛋白樣品。采用摩爾消光系數13 600 L/(mol?cm)計算總巰基含量。
1.3.3.3 表面疏水性測定
參考Chelh等[8]的方法。向1 mL肌原纖維蛋白分散液(5 mg/mL)中加入200 μL 1 mg/mL溴酚藍(bromophenol blue,BPB)(去離子水溶解)。空白組用20 mmol/L磷酸鹽緩沖液(pH 6.0)替代。室溫下充分振蕩10 min,5 000 r/min離心15 min。取上清液稀釋10 倍后在595 nm波長處測吸光度A。疏水性用BPB結合量表示,計算如式(1)所示:

1.3.3.4 內源性色氨酸熒光強度測定
色氨酸熒光強度的變化通過F-4600熒光分光光度儀進行檢測。用0.5 mol/L NaCl溶液將蛋白稀釋為質量濃度0.04 mg/mL,室溫條件下于295 nm波長處激發,記錄下310~400 nm波長處發射光譜供后續分析。扣除溶劑發射光譜(在相同條件下獲得)以排除干擾。
1.3.4 凍融循環對蝦蛄肌原纖維蛋白凝膠性能的影響
1.3.4.1 動態流變測試
根據Li Chunqiang等[9]的方法。將20 mg/mL蛋白樣品(分散于0.5 mol/L NaCl)置于離心管內,于4 ℃離心1 min脫氣。將脫氣后的蛋白均勻涂布于流變儀的下平板上,將上平板降至1 mm時用柔軟的實驗用紙輕輕擦去周圍多余的樣品,并在邊緣涂上硅油以防加熱過程中水分流失。在振蕩模式下以1 ℃/min的升溫速率加熱樣品,記錄20~80 ℃的升溫曲線。設置振蕩頻率為0.1 Hz,控制最大應力為0.02。凝膠性能通過儲存模量(G’)進行評價。
1.3.4.2 凝膠強度及持水力測定
將蛋白質量濃度調整為40 mg/mL,取20 mL于平底玻璃瓶中,加蓋密封,水浴加熱。具體升溫程序如下:起始溫度20 ℃,以1 ℃/min線性升溫至73 ℃。加熱結束后,立即將樣品放入冰水混合物中冷卻30 min,然后置于4 ℃冰箱過夜。在測定凝膠性能之前,將凝膠樣品從4 ℃取出放在室溫下平衡2 h。利用質構儀測定凝膠強度,選用P0.5圓柱探頭,測試全過程設定探頭速率均為1 mm/s,凝膠強度定義為破壞凝膠結構需要的初始力。
凝膠持水力通過離心的方法測定。將一定質量凝膠于4 ℃、6 000 r/min離心15 min,記錄離心管(m)以及離心前后的總質量分別為m1和m2。持水力的計算如式(2)所示:
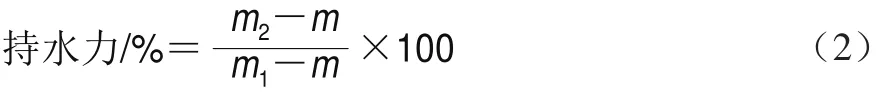
1.3.4.3 掃描電子顯微鏡觀測凝膠微觀結構
用0.1 mol/L pH 7.4的磷酸鹽緩沖液配制2.5%戊二醛溶液固定凝膠樣品12 h,從而獲得樣品塊(3 mm×3 mm×3 mm)。然后用0.1 mol/L磷酸鹽緩沖液(pH 7.4)洗滌3 次,在30%、50%、70%、90%、95%和100%乙醇溶液中梯度脫水。最后將樣品冷凍干燥后即可進行電子顯微鏡掃描。
1.3.5 凍融循環對蝦蛄肌原纖維蛋白乳化特性的影響
將8 mL 1 mg/mL的蛋白溶液與2.0 mL大豆油混合,15 000 r/min勻漿1 min,分別在第0、10分鐘時在距離心管底部0.5 cm處取100 μL勻漿液,加入到5 mL 0.1% SDS溶液中,混勻,以0.1% SDS為空白,500 nm波長處測定吸光度。蛋白乳化活性及乳化穩定性由式(3)、(4)計算:
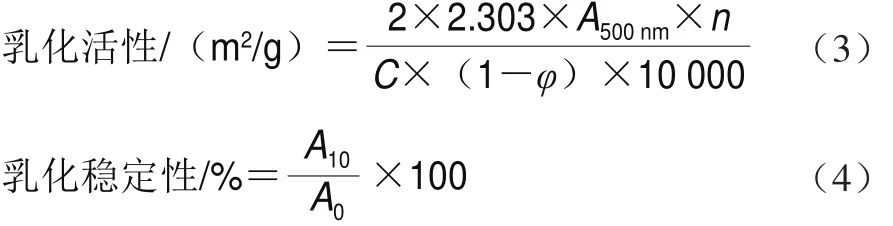
式中:A500nm為500 nm波長處的吸光度;n為稀釋倍數;C為蛋白質量濃度/(mg/mL);φ為乳化液中油的體積分數/%;A0和A10分別為第0、10分鐘時500 nm波長處吸光度。
1.4 統計學分析
2 結果與分析
2.1 凍融循環對蝦蛄肌原纖維蛋白氧化及結構的影響
2.1.1 蛋白羰基
蛋白羰基側鏈含—NH—或—NH2的氨基酸非常容易受到活性氧(reactive oxygen species,ROS)的氧化攻擊而轉化為羰基。因此,蛋白羰基含量的變化是目前最為廣泛用于衡量蛋白質氧化程度的指標。蛋白羰基產生途徑主要包括:氨基酸側鏈的氧化、肽鍵的斷裂、外源性羰基的引入(脂肪氧化產物中的醛類以及酚類物質的氧化產物半醌或醌類共價鏈接到蛋白側鏈)。一般來說,羰基含量越高表明蛋白氧化程度越高。經不同凍融次數處理后的蝦蛄肌原纖維蛋白羰基含量的變化如圖1所示。未經凍融處理的對照組和實驗組的蛋白羰基含量無差異,這說明冷凍保護劑海藻糖和甘露醇對肌原纖維蛋白羰基含量沒有影響。在進行1、3、5 次凍融處理后,蛋白發生不同程度的氧化變性,表現為羰基含量顯著上升,尤其在反復凍融5 次后較未經凍融的蛋白羰基含量增加了47.4%(P<0.05)。這與姜晴晴等[2]的研究結果一致。海藻糖和甘露醇的添加有效抑制了氧化誘導的羰基含量上升:其抑制效率在5.2%~14.4%之間。在冷凍過程中,蝦蛄肉中的水分部分凍結導致細胞內外溶質濃度增加,同時,冰晶生長的物理效應會破壞肌肉細胞,導致線粒體和溶酶體酶等滲入肌漿中,從而加速蛋白、脂肪的氧化反應[10]。冷凍保護劑如蔗糖、海藻糖、甘露醇、山梨糖醇等可以降低肉品的冰點[11],而且,海藻糖和甘露醇等可以與蛋白質分子的功能基團通過氫鍵或離子鍵相結合,這樣每個蛋白質分子都被冷凍保護劑分子所包覆,降低了因反復凍融引發的凍結和氧化傷害[12]。
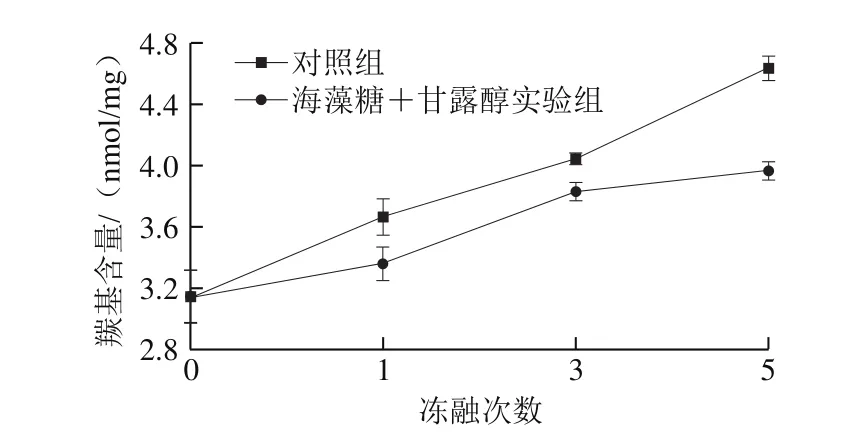
圖1 凍融處理對肌原纖維蛋白羰基含量的影響Fig. 1 Effects of freeze-thaw cycles on carbonyl content of myofibrillar protein
2.1.2 總巰基含量
肌原纖維蛋白主要由肌球蛋白和肌動蛋白組成,肌球蛋白約占其總量的43%,每分子含有40 個巰基,不含二硫鍵;肌動蛋白約占22%,每分子含有5 個巰基,也不含二硫鍵。巰基易受自由基攻擊而轉化成二硫鍵。因此,巰基含量的減少以及二硫鍵的形成是蛋白氧化的重要前期指標之一。由圖2可知,凍融處理對肌原纖維蛋白巰基含量的影響與羰基變化趨勢恰好相反。未經凍融處理的蛋白總巰基含量約為(44.26±0.74)nmol/mg,凍融處理1、3、5 次后,巰基含量分別下降了12.7%、17.7%、37.6%。這與Benjskul等[13]在大西洋白鱈冰藏過程中得到的研究結果相似。反觀實驗組,經1、3、5 次凍融處理后,蛋白巰基含量分別下降了6.2%、9.6%、25.5%,4 g/100 mL海藻糖+4 g/100 mL甘露醇顯著抑制了蛋白巰基含量的減少。在凍藏過程中肌原纖維蛋白巰基含量的下降可能是由于水形成冰結晶后,破壞肌原纖維蛋白的空間結構,使巰基暴露出來,進而被氧化成二硫鍵使巰基含量下降。Hamre等[14]同樣發現,大眼海魚在-18 ℃冷藏時總巰基含量隨冷藏時間延長而下降,而二硫鍵含量卻上升。海藻糖和甘露醇等冷凍保護劑作為蛋白質分子的防護屏障,能阻止肌肉中各種促氧化成分如自由基等對蛋白分子的攻擊,從而有效抑制蛋白巰基向二硫鍵的氧化轉化[15]。Li Yanqing等[16]研究發現蔗糖和山梨糖醇對凍藏6 個月后的魚糜蛋白總巰基含量也有顯著的保護作用。Zhou Aimei[17]和Etemadian[18]等也得到了類似的結論。
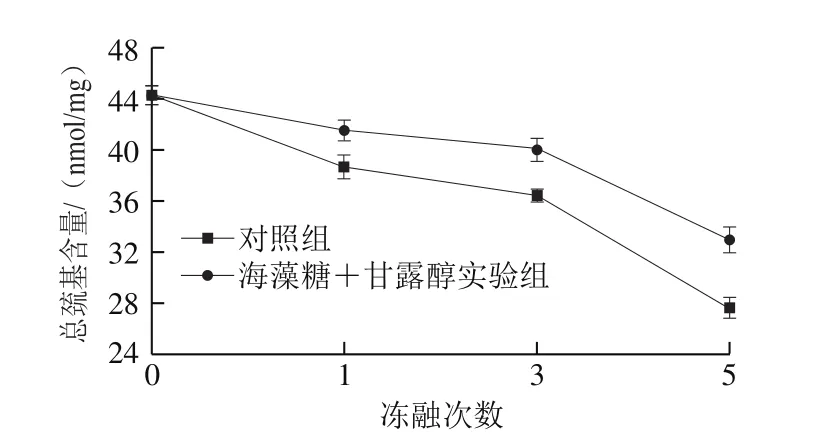
圖2 凍融處理對肌原纖維蛋白總巰基含量的影響Fig. 2 Effects of freeze-thaw cycles on total sulfhydryl content of myofibrillar protein
2.1.3 表面疏水性
BPB可以結合于蛋白質分子表面的疏水性位點,因此蛋白質所能結合的BPB量可被用作反應蛋白表面疏水性的指標[8]。一般來說,表面疏水性增加是蛋白質分子結構展開的一個重要指標。氧化導致蛋白結構展開,暴露出更多的疏水性氨基酸殘基使得蛋白表面疏水性增強。如圖3所示,與未經凍融處理的蛋白樣品相比,經1、3、5 次凍融處理的蛋白所結合的BPB量明顯上升,尤其是在5 次凍融循環后表面疏水性增加了1.48 倍,說明反復凍融導致肌原纖維蛋白結構展開、疏水性上升。這可能是因為凍融處理導致蛋白質氧化程度加深,表現為蛋白羰基化、肽主鏈的斷裂以及分子間二硫鍵的形成,蛋白質解折疊,一些疏水性的脂肪族與芳香族氨基酸側鏈基團從蛋白分子內部暴露,從而引起表面疏水性的增加[2,19]。海藻糖和甘露醇的添加對蛋白表面疏水性上升具有顯著的抑制作用,抑制率分別為10.7%(1 次凍融)、23.2%(3 次凍融)、19.6%(5 次凍融)。Herrera等[20]研究表明冷凍保護劑能減少非極性氨基酸側鏈基團的暴露,從而降低蛋白的表面疏水殘基數量,阻止蛋白分子因疏水相互作用引發的過度聚集。
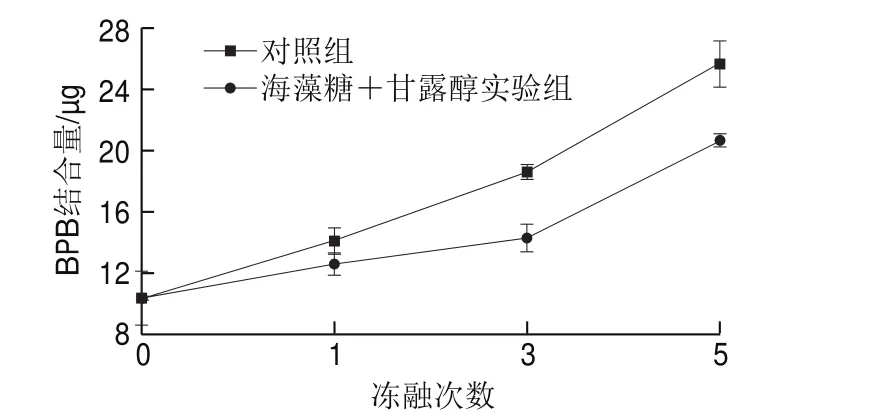
圖3 凍融處理對肌原纖維蛋白表面疏水性的影響Fig. 3 Effects of freeze-thaw cycles on surface hydrophobicity of myofibrillar protein
2.1.4 內源色氨酸熒光
蛋白質內源性色氨酸熒光對于其周邊微環境的極性非常敏感。當蛋白質處于折疊狀態時,色氨酸殘基主要位于蛋白質內核這樣的疏水環境中,此時被激發的色氨酸具有相對較高的熒光強度;當蛋白質部分或者完全展開時,色氨酸殘基就會更多的暴露于蛋白質分子表面,此時被激發的色氨酸熒光強度降低。因此,內源性色氨酸熒光特性常被用于蛋白三級結構的檢測[21]。如圖4所示,隨著凍融處理次數的增加,肌原纖維蛋白的熒光強度顯著下降,這表明蛋白三級結構的展開。反復凍融破壞了肌原纖維蛋白天然的排列方式,蛋白展開變性使得原本位于內部的色氨酸殘基暴露于表面而處于溶劑的極性環境中,從而引起內源熒光強度下降。添加海藻糖和甘露醇使熒光強度有不同程度回升,說明在二者的保護作用下,蛋白分子變性程度較低,結構變化相對較小,色氨酸殘基暴露少,這與上述表面疏水性結果相吻合。

圖4 凍融處理對肌原纖維蛋白內源色氨酸熒光強度的影響Fig. 4 Effects of freeze-thaw cycles on tryptophan fl uorescence intensity of myofibrillar protein
2.2 凍融處理對蝦蛄肌原纖維蛋白凝膠性能的影響
2.2.1 動態流變特性

圖5 凍融循環對肌原纖維蛋白在熱凝膠過程中彈性G’的影響Fig. 5 Effects of freeze-thaw cycles on storage modulus (G’) of myo fi brillar protein during thermal gelation
如圖5所示,未經凍融的蛋白在加熱過程中出現兩個典型的流變轉變峰,其峰值分別在44 ℃和50 ℃左右,第1個峰的出現可歸因于肌球蛋白頭部變性及交聯,第2個峰應是肌球蛋白尾部解旋并聚集的結果。在加熱早期階段,肌球蛋白的聚集主要是涉及二硫鍵交換的頭部-頭部結合,在隨后的加熱過程中是部分不可逆的尾部解螺旋及尾部-尾部交聯,緊接著就是三維凝膠網絡的形成[22]。
經1、3、5 次凍融循環后,肌原纖維蛋白G′在第1個峰值處急劇下降,較未凍融蛋白分別下降了14.6%、70.4%、86.7%,且最終G′值也隨著凍融次數的增加而顯著下降,這說明3 次及以上凍融循環嚴重破壞了肌原纖維蛋白的凝膠性能。此外,經凍融處理的蛋白G′的起始上升溫度與轉變峰的溫度較未凍融蛋白均有所提前,可能由于反復凍融破壞了蛋白結構,使得埋藏于蛋白分子內部的活性官能團暴露出來,從而加強了蛋白之間的相互作用[23]。海藻糖和甘露醇的添加顯著提高了蛋白在加熱過程中的G′,尤其在3、5 次凍融循環后,第1個轉變峰值較對照組分別提高了1 倍和2.8 倍。反復凍融導致肌原纖維蛋白變性,結構過度展開,蛋白聚集沉淀,活性功能基團被屏蔽,從而破壞蛋白凝膠性能。海藻糖和甘露醇等冷凍保護劑能與蛋白在連續相中相互作用,從而產生具有黏彈性的非晶態網絡結構[24]。Li Xingke等[25]的研究得到與本實驗類似的結論,他們發現殼聚糖對鰱魚鹽溶蛋白的凝膠性能同樣有促進作用。
2.2.2 凝膠強度及持水力
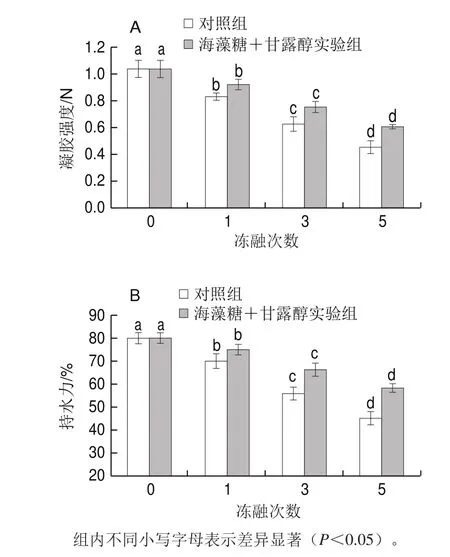
圖6 凍融循環對肌原纖維蛋白凝膠強度(A)和持水力(B)的影響Fig. 6 Effects of freeze-thaw cycles on gel strength (A) and waterholding capacity (B) of myofibrillar protein
如圖6所示,蝦蛄肌原纖維蛋白的熱誘導凝膠強度隨著凍融次數的增加顯著下降(P<0.05),蛋白經1、3、5 次凍融循環處理后其凝膠強度較未凍融蛋白分別下降了20.2%、39.7%、56.4%。Erikson等[26]也發現凍藏會顯著降低三文魚里脊肌原纖維蛋白的凝膠硬度。4 g/100 mL海藻糖+4 g/100 mL甘露醇則可以有效抑制蛋白凝膠強度的下降,抑制效率在11.2%~33.6%之間。凍融循環導致蝦蛄肌球蛋白和肌動球蛋白變性,以致于在隨后的熱處理過程中無法形成致密的凝膠網絡結構[27],因此得到較低的凝膠強度。海藻糖和甘露醇的加入抑制了蛋白氧化和變性,降低蛋白聚集程度,Kong Baohua等[15]也證實了這一點。冷凍保護劑與蛋白質分子結合后,使得肌原纖維蛋白凝膠網絡結構中的自由水被捕獲,有效阻止蛋白冷凍變性[27]。因此海藻糖和甘露醇可以提高蝦蛄蛋白的成膠能力,這與動態流變結果一致。
凝膠持水力指凝膠保持自身水分或補充水分的能力,與凝膠內部結構的粗糙程度有關。蝦蛄肌原纖維蛋白凝膠持水力的變化趨勢與凝膠強度一致。肌球蛋白組分對凝膠的形成起關鍵作用[28]。樣品中肌球蛋白參與凝膠的程度隨著蝦蛄凍融次數的增加而降低,因此持水力也隨之下降。海藻糖和甘露醇的添加有利于凝膠持水力的提升,相較于對照組分別提高了7.1%(1 次凍融)、18.7%(3 次凍融)、29.0%(5 次凍融)。海藻糖和甘露醇等冷凍保護劑一方面可以通過氫鍵作用結合自由水,固定肉糜中的水分;另一方面也可以減少水分子在網絡結構中的位移,從而提高持水力[27]。
2.2.3 凝膠微觀結構
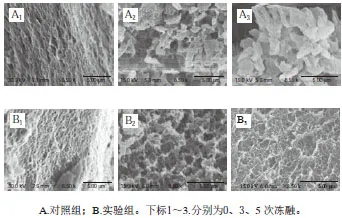
圖7 凍融循環對肌原纖維蛋白凝膠微觀結構的影響Fig. 7 Effects of freeze-thaw cycles on microstructure of myofibrillar protein gels
通過分析動態流變、凝膠強度與持水力結果,本實驗選取未經凍融和經3、5 次凍融處理的蛋白進行凝膠微觀結構表征。如圖7所示,對照組和實驗組的未凍融蛋白的熱制凝膠結構致密,表面較為平整,孔隙分布均勻。經3、5 次凍融處理后(放大掃描電子顯微鏡觀察倍數),蛋白凝膠結構劣變且程度逐漸加深,產生大量不規則的聚集體,三維網狀結構不明顯,說明肌原纖維蛋白在反復凍融過程中發生聚集,這與動態流變、凝膠強度與持水力的結果一致。添加海藻糖和甘露醇顯著改善了蛋白的凝膠結構,表現為促使不規則的聚集體參與凝膠而形成多孔且孔徑均一有序的三維網絡結構,對應提高的凝膠強度及持水力。
2.3 凍融處理對蝦蛄肌原纖維蛋白乳化特性的影響
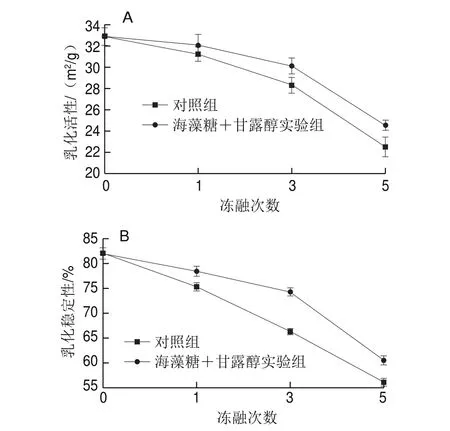
圖8 凍融循環對肌原纖維蛋白乳化活性(A)和乳化穩定性(B)的影響Fig. 8 Effects of freeze-thaw cycles on emulsifying properties of myofibrillar protein
如圖8所示,隨著凍融次數的增加,肌原纖維蛋白的乳化活性和乳化穩定性都顯著下降(P<0.05),其中凍融處理5 次后乳化活性和乳化穩定性下降了31.6%,這與Xia Xiufang等[29]在豬肉中的研究結果一致。此結果表明,反復凍融破壞了肌原纖維蛋白結構的完整性,導致蛋白與脂肪交聯的能力下降,從而使其乳化活性及乳化穩定性下降。添加海藻糖和甘露醇后,蛋白的乳化活性和乳化穩定性均有不同程度回升。蛋白的乳化性受眾多因素影響,其中蛋白變性是導致乳化能力下降的主要原因,氧化變性會限制蛋白在油滴表面吸附的靈活性[27]。此外,Srikar等[30]研究表明脂肪和游離脂肪酸的氧化產物也會降低蛋白的溶解性和乳化能力。結合研究結果和相關文獻[15],冷凍保護劑能顯著抑制蛋白羰基化和脂質氧化,防止蛋白變性,從而起到穩定乳狀液的作用。
糖醇類物質的抗凍作用機理是分子中的羥基與蛋白質分子的某些功能基團結合,使蛋白質分子處于飽和狀態,從而避免蛋白質分子之間的聚集變性。同時,糖醇類物質的游離羥基還能有效地束縛水分子,從而降低“共晶點”溫度,減少冰晶體的形成量,形成一個不完全凍結區域,隔離和減緩蛋白質分子的聚集,進而防止蛋白質的凝聚變性。一般,糖醇類物質的添加量與防止蛋白質冷凍變性的效果呈正比[31]。
3 結 論
本實驗研究海藻糖和甘露醇對反復凍融引起的蝦蛄肌原纖維蛋白結構和功能特性變化的影響。結果顯示經過反復凍融處理,肌肉結構完整性破壞,蛋白氧化變性,表現為蛋白質羰基含量和表面疏水性增加,總巰基含量和色氨酸熒光強度顯著下降,同時蛋白的功能性質(凝膠和乳化特性)下降,影響其食用及加工利用品質。而海藻糖和甘露醇的添加能有效抑制蛋白的變性程度,減少蛋白結構的變化,顯著提高蛋白在熱凝膠過程中的G’,改善蛋白的凝膠特性(凝膠強度、持水力和三維網絡結構)、乳化活性和乳化穩定性。海藻糖和甘露醇作為食品添加劑添加到低溫肉制品中,可改善肉品的膠著性、持水性和柔嫩性,減少營養損失,提高產品質量,具有廣闊的應用前景。
猜你喜歡
云南醫藥(2021年6期)2022-01-08 04:07:38
中國民間療法(2021年5期)2021-06-09 09:21:26
華聲文萃(2019年12期)2019-09-10 07:22:44
中西醫結合心血管病電子雜志(2017年17期)2017-11-17 15:54:41
上海農業學報(2017年4期)2017-04-10 12:40:22
中西醫結合心血管病雜志(電子版)(2017年17期)2017-01-11 06:37:25
食藥用菌(2016年6期)2016-03-01 03:24:27
中國繼續醫學教育(2015年4期)2016-01-07 07:38:11
護士進修雜志(2015年5期)2015-03-18 19:22:53
中國藥業(2014年24期)2014-05-26 09:00:09
