干旱沙漠區2種深根植物根際微生物數量分布及根際效應研究
2019-09-06 03:38:09高歡歡曾凡江趙秀芳朱心寧張倩丁澤華
現代農業科技 2019年8期
高歡歡 曾凡江 趙秀芳 朱心寧 張倩 丁澤華

摘要 本研究以古爾班通古特沙漠東南緣2種深根植物疏葉駱駝刺、多枝檉柳不同土層深度根際土壤、根外土壤為研究對象,運用平板劃線法研究根際土壤、跟外土壤微生物數量,并對其根際效應進行探討。研究結果顯示,細菌占根際可培養微生物數量中的絕大多數。每種植物根際細菌數量、微生物總數在不同土層有相同的波動變化規律。根際真菌、放線菌在不同植物不同土層也呈現差異性分布。根際細菌數量、微生物總數排序為疏葉駱駝刺>多枝檉柳根際真菌放線菌數量排序為疏葉駱駝刺>多枝檉柳。除多枝檉柳根際細菌放線菌、微生物總數在0~50cm和200~250cm土層呈現負效應,其余樣本根際效應均為正效應。
關鍵詞 根際微生物;平板劃線法;根際效應;干旱沙漠區
中圖分類號 S154.3
文獻標識碼 A
文章編號 1007-5739(2019)08-0172-04
細菌、真菌、放線菌是可培養微生物的主要分類方式,土壤中可培養微生物是微生物群落結構的重要組成部分,由于三大微生物計數手段的簡便易行使其成為評價微生物多樣性及土壤狀況的重要指標"。土壤中三大微生物數量的變化既能反映出微生物對養分、水分、光照、植物分泌物等外界生境因子的變化,又能體現微生物群落自身的應激策略。根際作為特殊的微生物生境,是土壤微生物鏈接植物與環境參與能量物質轉換的重要場所。游走于根際的微生物從植物根系分泌物及枯落物中獲取有效碳源和氮源,同時為植物生長提供必要的小分子元素與微環境吧。在根際中,植物一微生物形成相互依托的互助體。由于根際與根外環境存在顯著差異性,導致根際、根外土壤微生物數量、理化性質等均存在差異,即根際效應。通常情況下,將根際、根外微生物數量的比值作為衡量根際效應的重要指標,其比值大于1說明根際呈現正效應,比值小于1說明根際呈現負效應用。
在荒漠這種干旱生態系統中,降雨稀少、沙漠化嚴重、土壤養分貧瘠是其主要特點可。雖然生活條件非常苛刻,但是在世界各地的沙漠中,微生物數量和種類都很多,達到1.6x107個/g。早期研究發現,放線菌占據了沙漠土壤細菌總數的近50%7,研究發現沙漠野生植物的根際細菌種類比根外土壤多1.5~3.0倍。隨著分子生物學的發展,更多種類的微生物群落已經在荒漠生態系統被發現。在以往的研究中將每個土層中所有根系的根際微生物作為一個整體去研究,而根據前人大量研究發現,不同根系的根功能不同,分為吸收根和運輸根8。GUI等通過人工挖掘法研究了自然條件下不同潛水埋深疏葉駱駝刺克隆分株生長情況和根系結構。挖掘中發現大部分深根系植株擁有發達的地下根系,疏葉駱駝剌主根直達地下水,但細根的含量相對較少。根系吸收養分、水分的具體位置還不確定。因此,不能單純以某段較淺土層根際微生物群落特征代表該植物根際微生物群落的全部特征,垂直梯度上根際微生物的研究十分關鍵。
在本研究中,以古爾班通古特沙漠中2種深根植物根際土壤根外土壤微生物為研究對象,對不同植物不同土壤深度下根際細菌、真菌放線菌、微生物總數數量分布特征及相應根際效應進行研究,探討根際可培養微生物群落多樣性。
1材料與方法
1.1樣品采集
本研究于2014年7月在古爾班通古特沙漠東南緣天然試驗場進行。選取自然生長的2種深根植物疏葉駱駝刺(AlhagisparsifoliaShap.)、多枝檉柳(TamarixramosissimaLedeb.),分別選取立地條件相似、地下潛水埋深基本相同的3個20mx20m的單純植被樣方,共計9個樣方進行調查取樣。
取樣前所有工具、容器都要經過嚴格的消毒,防止微生物交叉污染。正式取樣前,先將土壤表層、根系周圍枯落物清除,然后,將土壤表層1cm左右的腐殖質層剔除。
每個樣方分別隨機選取6~10株植物,對所調研的物種根系進行垂直剖面的挖掘。先用挖掘機挖掘周邊大型土方,制造出取樣剖面,再進行人工挖掘清理根系周圍土壤,進行根際土壤樣品的采集。采集深度3.5m,每隔50cm作為一個取樣深度。采用自然環境中采集的根際、根外土壤樣品作為試驗材料。根際及根外土壤細菌、真菌、放線菌、微生物總數計數選取的土壤樣品均在49C條件下保存。
1.2微生物計數
微生物培養:采用劃線平板法將接種后的培養皿倒置,細菌培養2~3d,真菌培養3~5d,放線菌培養5~7d,然后取出,選擇菌落數適合的培養皿(細菌和放線菌為20~200個,真菌為10~100個),剔除占瓊脂表面超過15%的菌落。按照下式計算菌數:
菌數(CFU/g)=.菌落平均數x稀釋倍數x20x鮮土重千土重
2結果與分析
2.1多年生植物根際三大微生物數量垂直分布特征
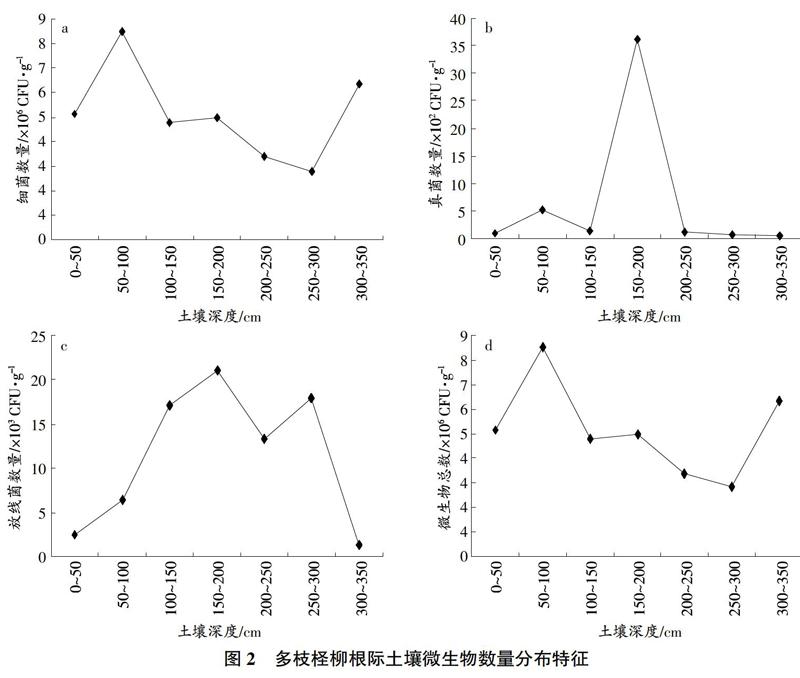
駱駝刺根際土壤細菌、真菌、微生物總數在不同土壤深度呈現雙峰波動分布模式,放線菌數量在不同土壤深度呈現單峰分布模式(圖1)。細菌數量和微生物總數呈現一致的波動規律,即隨士壤深度的增加,細菌數量和微生物總數上升下降交替波動,在100~150cm、200~250cm土層出現峰值,真菌數量也在這2個土層出現峰值。放線菌在150~200cm土層中分布最多。
與疏葉駱駝刺相同的是,多枝檉柳根際細菌數量和微生物總數也具有相同的分布模式,但與疏葉駱駝刺分布模式不同(圖2)。多枝檉柳根際細菌數量和微生物總數在50~150cm土層出現數量最大值,隨后隨土層增大而逐漸減少,在300~350cm土層又增大。真菌數量呈現單峰分布模式,在150~200cm土層最多。放線菌數量分布呈雙峰分布,在150~200cm和250~300cm出現2個峰值。
2.2多年生植物根際效應垂直分布特征
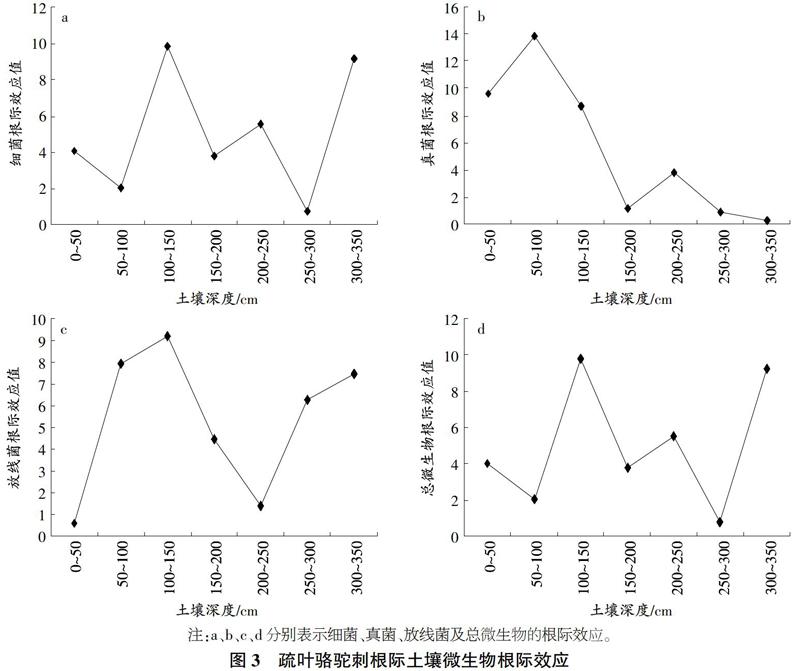
根據ANOVA分析,不同植物種根際土壤中各可培養微生物根際效應都不存在顯著差異,因而不進行不同植物種間對比分析,僅分析每個植物種根際土壤微生物數量分布特征。
如圖3所示,疏葉駱駝刺200~250cm細菌、真菌,250~300cm土層真菌、微生物總數,0~50cm土層放線菌等根際效應呈現負效應,其余各可培養微生物根際效應均呈現正效應。細菌、真菌、放線菌微生物總數根際效應均值分別為5.01、6.32、4.96、5.02。細菌、微生物總數根際效應波動規律相似,在100~150em土層根際效應最大,250~300cem土層根.際效應最小。真菌根際效應呈現波動下降趨勢,在50~100cm土層最大,為13.86;當土壤深度達到300~350cm時,根際效應下降到0.25。放線菌根際效應隨土壤深度的增加呈現先增大后減小再增加的趨勢,在100~150cm土層最大;當土壤深度達到200~250cm時達到最低點,而后又逐漸上升。
多枝檉柳根際土壤細菌、微生物總數根際效應波動變化規律一致(圖4),在0~50cm和200~250cm2個土層呈現負效應,其余土層呈現正效應;在50~100cm根際效應最大,分別為6.13.6.12。真菌根際效應均呈現正效應,波動規律呈現先增大后減小的單峰分布,在150~200cm土層真菌根際效應最大,為116.40。放線菌根際效應隨土層的增加呈現逐漸遞增趨勢,在0~50cm土層呈現負效應;隨土壤深度的增加呈現正效應,當土壤深度到達250~300cm時,放線菌根際效應到達最高值10.17,隨后又降低。
2.3根際微生物數量及根際效應相關性分析
將不同植物不同土壤深度根際士壤各微生物數量及根際效應進行兩兩相關性分析,由表1可知,根際細菌數量與根際微生物總數呈現極顯著正相關性(P<0.01),相關系數接近1.00。根際放線菌數量與根際真菌根際效應呈現極顯著正相關性(P<0.01),相關系數0.81。其余參數之間不存在顯著相關性(P>0.05)。
3結論與討論
根際微生物作為根際生態系統的重要組成部分,其數量及分布對根際生態系統中土壤的健康及植被的恢復具有重要意義0,根際微生物數量受植被類型、土壤性質等影響存在空間異質性分布"。在我國干旱生態系統微生物研究中發現,千旱半干旱區域土壤存在大量可培養微生物,其數量與分布隨土壤狀況變化存在動態變化。與以往研究相似,本研究區域根際可培養微生物總數量化級別10,不同植物種間根際細菌數量、根際微生物總數差異顯著。頭狀沙拐棗根際細菌、微生物總數最多,疏葉駱駝刺次之,多枝檉柳最少。
在2種植物根際土壤可培養微生物中,細菌都是為優勢菌群,量化度為106;放線菌次之,量化度為10;真菌最少,量化度為102。從數量上,疏葉駱駝刺、多枝檉柳的根際細菌分別占微生物總數的99.32%和99.81%。這一結果導致了根際細菌數量與根際微生物總數呈現極顯著的正相關關系。在根際微生物數量隨土壤深度增加的分布中,每種植物根際細菌、微生物總數的分布規律都一致,這也印證了細菌在根際微生物數量中的主體地位。在騰格里沙漠、塔克拉瑪干沙漠的研究中也發現,在千旱半千旱區域的荒漠土壤中,土壤中細菌數量占可培養微生物總數的主體地位,本研究與前人研究一致。
在本研究中,根際真菌與根際細菌呈現不同的空間分布異質性,在古爾班通古特沙漠不同土壤深度中2種植物根際真菌數量及根際效應均呈現單峰分布。但利用劃線平板法測定可培養真菌數量并未發現與已經研究的根際生境因子呈現顯著的相關關系,對根際可培養真菌空間異質性還需要進一步探討。
放線菌在研究區域古爾班通古特沙漠根際土壤微生物數量中所占比重最少,但是以往的研究表明,根際放線菌從植物及土壤中吸收必要的養分與能量,通過生命活動釋放嘌呤類似物調控土壤健康和根系生存環境"5,與土壤、植物存在明顯的互惠互利生存關系,在千旱、貧瘠的沙漠土壤占據重要地位。根際放線菌空間異質性分布主要受土壤可溶性離子影響,可以有效阻隔有害真菌對植物的侵染,分解土壤有機質改善土壤養分狀況",從而促進植物生長與植被修復。本試驗中根際放線菌與根際土壤電導率呈現極顯著的正相關關系,而電導率主要受土壤中可溶性離子影響,說明在本區域電導率范圍內,離子含量越大,根際放線菌數量越多。
根際效應是衡量根際微生物生命活性的重要指標。微生物根際效應:主要受根際微環境因子(如土壤類型、養分狀況、pH值、濕度等)以及植物因子(如物種年齡等)的影響18。根際效應的空間分布異質性反映根際微生物活性的空間分布異質性。本研究中,由于細菌數量占微生物總數的大部分,故2種植物細菌根際效應與微生物總數根際效應都呈現一致的分布規律。試驗結果可以看出,2種植物真菌根際效應均呈現正效應,真菌根際效應與根際真菌數量呈現極顯著相關性(P<0.01),而其他微生物根際效應與根際相對應的微生物數:量無顯著相關性(P>0.05)。這說明極端干旱區土壤中真菌數量雖然不占主體地位,可能由于根際對土壤真菌吸引能力較強,真菌主要活動在根際范圍內造成真菌較高的根際效應。
4參考文獻
[1]吳楠,張元明,潘惠霞,等.古爾班通古特沙漠地衣結皮中可培養細菌多樣性初探[J].中國沙漠,2013,33(3):710-716.
[2]衛云燕,尹華軍,劉慶,等夜間增溫和施肥對川西亞高山針葉林兩種樹苗根際效應的影響[J].生態學報,2011,31(3):698-708.
[3] PHILLIPS R P,FAHEY T J.the influence of soil fertility on rhizosphereeffects in northern hardwood forest soils[J].Soilence Society of AmericaJournal , 2013,72(2):453- -461.
[4]王衛霞.新疆幾種典型荒漠植物根際微生物特征及內生固氮菌的分離促生性能研究[D].烏魯木齊:新疆農業大學,2009.
[5] NOYMEIR I.desert ecosystems : environment and producers [J].AnnualReview of Ecology & Systematics,1973,4(1):25- -51.
[6] SKUJINS J.microbial ecology of desert soils[J]. Advances in MicrobialEcology , 1984,7:49-91.
[7] BHATNAGAR A, BHATNAGAR M.Microbial diversity in desert ecosys-tems[J].Current Science , 2005, 89( 1):91-100.
[8]劉波.多年生植物細根主要功能特征及其變異格局[D].烏魯木齊:中國科學院新疆生態與地理研究所,2013.
[9] GUI D.Characteristics of the clonal propagation of Alhagi sparsifolia Shap.(Fabaceae )under different groundwater depths in Xinjiang, China小Rangeland Journal , 2013 ,35(3):355-362.
[10]何尋陽,蘇以榮,梁月明,等喀斯特峰叢洼地不同退耕模式土壤微生物多樣性[J].應用生態學報,2010,21(2):317-324.
[11] BRUNDRETT M C.Tansley review no.134:Coevolution of roots andmycorrhizas of land plants [J].New Phytologist , 2002, 154(2):275-304.
[12]李從娟,雷加強,徐新文,等.古爾班通古特沙漠土壤水分與化學性質的空間分布[J].生態學報,2014,34(15):4380-4389.
[13]張憲武,許光輝.騰格里沙漠地區沙地士壤微生物學特性的研究[J].土壤學報,1962(3):227-234.
[14]靳正忠,雷家強,李生宇,等.流動沙漠腹地灌木根際土壤微生物比較分析[J].應用與環境生物學報,2010,16(6):759-764.
[15] GUNATILAKA A A L.Natural products from plant- -associated microor-ganisms : Distribution , structural diversity , bioactivity , and implicationsof their occurrence[J].Journal of Natural Products , 2006 , 69(3) :509.
[16] MCLELLAN C A,TURBYVILLE TJ, WIJERATNE E M , et al.A rhizos-phere fungus enhances Arabidopsis thermotolerance through productionof an HSP90 inhibitor[J].Plant Physiology , 2007, 145(1):174-82.
[17]庹利,旭格拉.哈布丁,郭琳,等.羅布泊地區沙生植物根際放線菌多
樣性及生物活性的研究[J].中國抗生素雜志,2012,37(1):21-26.
[18] WARDLE D A.A comparative assessment of factors which influencemicrobial biomass carbon and nitrogen levels in soil [J]. BiologicalReviews, 1992 , 67(3):321-358.
