
FOXO1 和FOXO3a 基因靶向轉導對缺氧/復氧小鼠心肌細胞的作用研究
2020-04-17 00:20:20李繼紅阿爾祖古麗米爾阿卜杜拉金曉倩黃鶯通訊作者馬依彤馬翔劉芬陳邦黨劉成
醫藥前沿 2020年35期
李繼紅 阿爾祖古麗·米爾阿卜杜拉 金曉倩 黃鶯(通訊作者) 馬依彤 馬翔 劉芬 陳邦黨 劉成
(1 新疆醫科大學第一附屬醫院冠心病二科 新疆 烏魯木齊 830054)
(2 新疆心血管病研究所 新疆 烏魯木齊 830054)
心肌梗死是心力衰竭是主要原因之一。研究其發病機制,有助于尋找新的干預措施,來提高預防及治療效率,達到提高患者的生存率及生活質量、改善冠心病心肌梗塞預后的目的。與衰老有關的氧化應激是缺血性心力衰竭的發病機制之一,氧化應激所產生的機體活性氧簇(ROS)促使動脈硬化斑塊破裂,導致冠狀動脈血栓形成;急性心肌梗塞發生時,ROS 會參與到組織損傷反應[1-2]。研究顯示,ROS 的不良作用可能被抗氧化應激反應所抑制,起到改善缺血性心力衰竭預后的作用。叉形頭轉錄因子O 亞族(forkhead box transcription factor O,FoxO)是重要的抗氧化應激調節因子,過表達FOXO1 可以抑制ROS 并減少心肌細胞死亡[3],但轉基因的方法會導致胚胎致死,因此需要尋找合適的載體進行靶向基因轉導。據報道,Fox O1、Fox O3a 及 Fox O4 對維持心功能和調節心臟應力有至關重要的作用[4]。其中,FoxO3a 能夠抑制腫瘤進展,調節血管內皮細胞、心肌細胞及血管平滑肌的生物學功能,同時有明確的抗氧化應激作用,因此,可能可用于心血管疾病的診治。因此本研究擬通過觀察重組腺病毒靶向轉導Fox O1、Fox O3a基因對缺氧/復氧(H/R)心肌細胞的作用,探討其抗氧化作用及由此帶來的細胞功能變化,報告如下。
1.資料與方法
1.1 一般資料
將H9C2 小鼠的心肌細胞先缺氧(H)3h 后,復氧(R)6h,然后建立缺氧/復氧損傷模型;然后隨機分為5 個組:A 組為Control 組,B 組為缺氧/復氧(H/R)組),C 組為缺氧/復氧+FOXO1 組,D 組為缺氧/復氧+FOXO3a 組、E 組為缺氧/復氧+FOXO1+FOXO3a 組。
1.2 方法
1.2.1 檢測細胞中的蛋白表達情況 細胞裂解后進行蛋白定量,20μg/孔,加上樣緩沖液沸水浴后離心取上清上樣,PAGE膠放入電泳槽并加緩沖液,加樣,80V 20min;電泳后膠移入轉膜緩沖液,剪下PVDF 膜,甲醇浸泡3min,水洗2min,剪6 塊濾紙與PVDF 膜,然后在轉膜緩沖液中平衡15min。轉膜裝置設定為恒流200mA,40min。膜洗1 次(放入TBST 中),然后置于麗春紅染色液中,5min 后水洗膜至水變清無色蛋白條帶清晰。5% BSA 室溫封閉1 h,一抗和膜4 ℃中孵育過夜。孵育一抗的膜洗滌(實驗室TBST)3 次后按1 ∶5000 稀釋二抗,與膜37℃孵育1 h。洗滌3 次,于化學發光儀中進行拍照。
1.2.2 氧化應激相關指標活性氧的水平 稀釋好的蛋白0.1ml 加入,4℃放置一夜。次晨,清洗。在已包被好的反應孔加 0.1ml稀釋好的標本,37℃培育1h。將稀釋好的酶標抗體 0.1ml加入到實驗孔,37℃培育0.5 ~1h。把TMB 溶液滴人各反應孔中,培養(在37℃,10 ~30 min)后使用 H2SO4( 0.05ml)終止實驗。對其吸光度OD 值進行測量:調整ELISA 檢測儀波長為450nm,或用波長為410nm 的ABTS 顯色。見圖1。
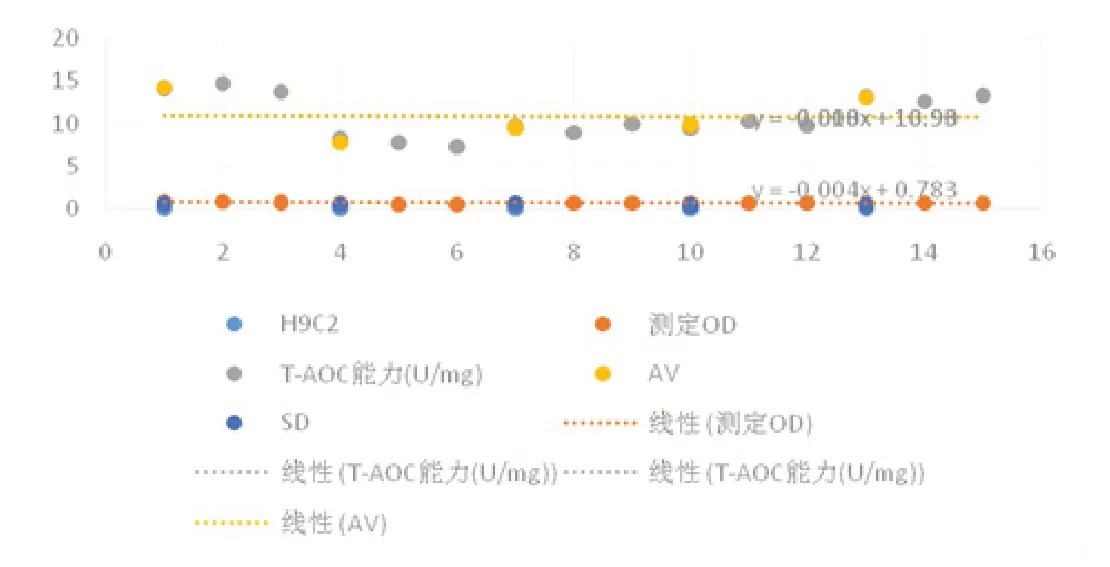
圖1 組織細胞中T-AOC 能力
1.2.3 檢測細胞凋亡情況 爬片細胞固定(4%多聚甲醛)20 ~30min,進行室溫(用百分之一TritonX-100)通透3 ~5min;滴加100μl DNase I 反應液,37℃30min,并且配制TdT 酶反應液: 37 度避光反應60 min,每個樣本中滴加50μl Streotavidin-Fluorescein 標記液后放入濕盒,37 度避光反應30min,Hoechst 復染細胞核:用pH7.4 的PBS 洗滌3 次,去除PBS 后,加入Hoechst 染液。孵育10min(避光室溫)。鏡下觀察并拍照。
1.2.4 Brdu 法檢測細胞增殖活力 H9C2 細胞以5×104cells/ml細胞數接種于24孔板中,1ml/孔,溫度為37 ℃,培養(在5% CO2培養箱中)24h。對各組細胞進行不同處理,在48h 后終止細胞培養之前,加入BRDU(50μM),37℃孵育40min。吸棄培養液,靜置5min 后吸出液體,重復5 次。甲醇/醋酸固定10min。經固定的玻片空氣干燥,用0.3% H2O2-甲醇30min 滅活內源性氧化酶。用封閉(5%的BSA)。變性核酸5min(100℃的甲酰胺)。冷卻。使用PBS 洗滌,加入抗大鼠BrdU 單抗,4℃過夜孵育。加PBS 洗滌,并重復5 次。加熒光二抗,在37℃中孵育1h。加PBS洗滌,再重復5 次。使用Hoechst 33258 染液孵育30min(室溫避光)后,鏡下觀察拍照保存。
2.結果
2.1 轉染后H9C2 細胞蛋白表達
WB 結果顯示:FOXO1 在B 組表達最低,FOXO1 病毒作用后表達會明顯的上升,且高于A 組;FOXO3a 病毒基本不會影響FOXO1表達;FOXO3a 蛋白在B 組中表達也是會最低,FOXO3a 病毒作用后其表達則會明顯的上升,且高于A 組,FOXO1 病毒對FOXO1 蛋白的表達有一定影響。見圖2。
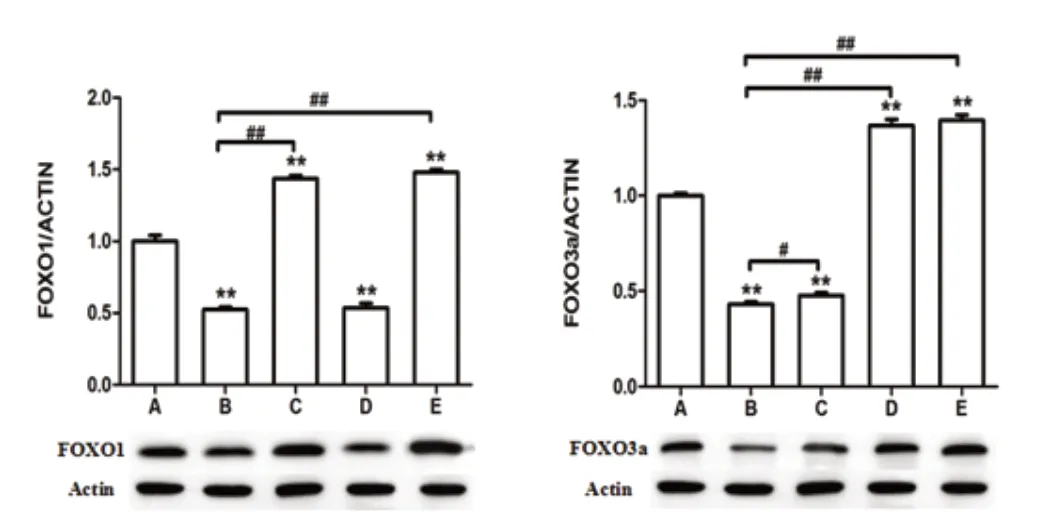
圖2 Western blot 檢測細胞中FOXO1 與FOXO3a 的表達
2.2 FOXO1 與FOXO3a 對H9C2 細胞缺氧/復氧時氧化應激相關指標的影響
ELISA實驗結果提示:SOD與T-AOC活性的表達趨勢基本相同,均為B 組中表達最低。FOXO1 與FOXO3a 基因轉導后表達升高明顯;MDA 含量、ROS 活性及NO 含量檢測趨勢基本相同,均為B 組中表達最高,FOXO1 或/與FOXO3a 基因轉導后表達降低明顯。見圖3。
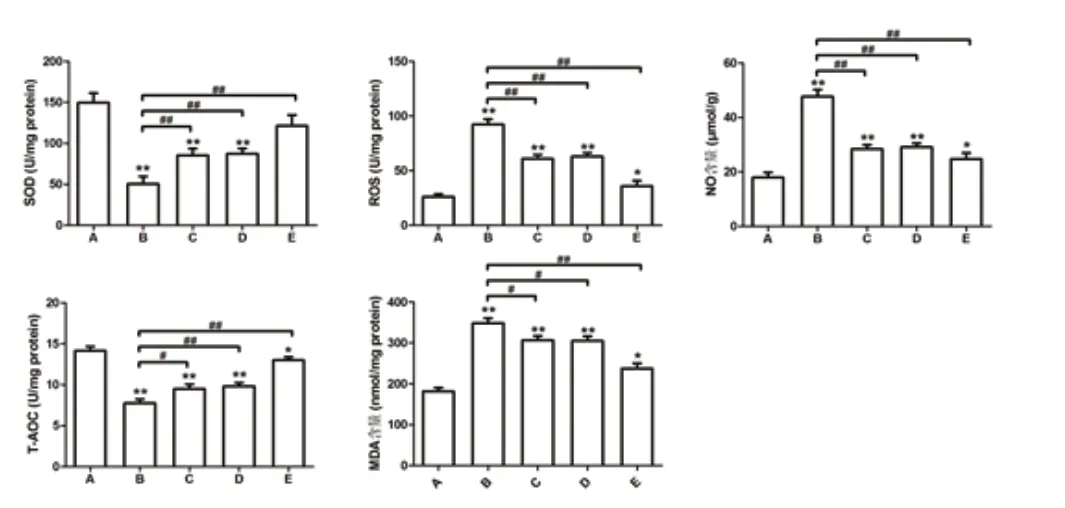
圖3 用ELISA 法檢測細胞氧化應激相關指標的水平
2.3 FOXO1 與FOXO3a 的基因轉導對缺氧/復氧H9C2 細胞凋亡的影響
TUNEL 結果提示:藍光代表正常細胞核,綠光代表凋亡的細胞核;A 組的凋亡是最少的,B 組的凋亡情況最多,C 和D 組的凋亡率顯著低于B組,而E組的凋亡進一步下降,但仍高于A組。見圖4。
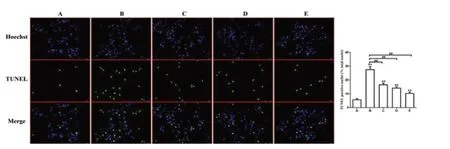
圖4 TUNEL 檢測H9C2 細胞的凋亡情況
2.4 FOXO1 與FOXO3a 基因轉導對于缺氧/復氧H9C2 細胞增殖活力的影響
Brdu 檢測結果提示:藍光代表正常的細胞核,紅光代表增殖的細胞核,A 組的增殖能力顯著高于其他四組;B 組的增殖能力最低;C、D 組的增殖能力要顯著高于B 組,但比A 組低;E 組的增殖能力高于C 組及D 組,比A 組略低。見圖5。
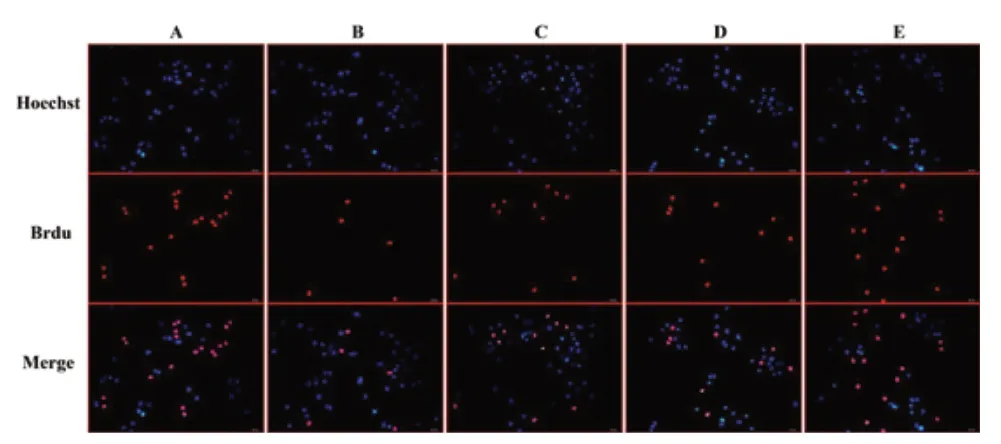
圖5 Brdu 法檢測缺氧/復氧H9C2 細胞的增殖活力
3.討論
目前,缺血性心肌損害已成為心力衰竭的首要病因,而氧化應激反應在心力衰竭的發生及進展中起到了至關重要的作用。FoxO 是重要的抗氧化應激調節因子。既往研究顯示:在肝細胞中,通過上調FOXO1 的蛋白的表達,可抑制裸鼠皮下移植性肝腫瘤的生長[3];成骨細胞中增強FOXO1 的活性可降低Bcl-2 表達,使成骨細胞生長受到抑制。有研究表明,FOXO1 還可激活FasL、CD95、Bim 促凋亡蛋白的表達[5-6],提示了FOXO1 重新激活可能是細胞存活與凋亡的關鍵調節因子之一。而通過調節FOXO3 的表達,能夠阻礙缺血性損傷和心肌梗死患者病情的進展。FoxO3a轉錄因子已被證實可抑制 VSMCs 增殖并有抗氧化應激作用,繼而可能會降低高血壓對機體的影響[7]。心血管疾病中Fox O3a 有保護和損傷的雙重作用,如何更好地利用其調控機制治療心血管疾病值得進一步研究。
本研究顯示,FoxO1、FoxO3a 被重組腺病介導后轉染缺氧/復氧心肌細胞,有效的減少了細胞的凋亡。據此得出結論,FOXO1 與FOXO3a 的基因轉導,減輕了氧化應激反應對H9C2 心肌細胞的損傷。H9C2 心肌細胞的增殖、存活能力明顯的改善。此研究結果,為缺血性心臟病心力衰竭的基因治療提供了新的研究數據,并開拓了新的研究領域,為藥物研究提供了新的靶點。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
海峽科技與產業(2016年3期)2016-05-17 04:32:12
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
