陽澄湖原產及“洗澡”中華絨螯蟹背甲的幾何形態分析
2020-06-12 12:51:48薛竣仁陳修報駱仁軍劉洪波
水生生物學報 2020年3期
關鍵詞:差異
薛竣仁 姜 濤 陳修報 駱仁軍 劉洪波 楊 健,
(1. 南京農業大學無錫漁業學院, 無錫 214081; 2. 中國水產科學研究院長江中下游漁業生態環境評價與資源養護重點實驗室,中國水產科學研究院淡水漁業研究中心, 無錫 214081)
中華絨螯蟹(Eriocheir sinensis)又稱河蟹、大閘蟹, 隸屬于節肢動物門(Arthropoda)、甲殼總綱(Crustacea)、十足目(Decapoda)、絨螯蟹屬(Eriocheir)[1]。該蟹原產于我國, 目前已入侵至歐洲[2]及美洲[3]。大閘蟹為我國傳統的名貴水產品和重要的經濟甲殼類動物。2017年其年總產量已經達到了7.5×108kg[4]。其可食部分富含礦質元素[5]、粗蛋白、粗脂肪與氨基酸等[6], 具有較高的營養價值。
我國中華絨螯蟹養殖主要集中在長江流域, 其中江蘇省為我國中華絨螯蟹養殖的主產地, 年產量3.5×108kg[4], 約占全國產量的50%[7]; 特別是蘇州市陽澄湖中華絨螯蟹因其歷史悠久、品優味美最為著名, 且于2005年被原國家質檢總局認定為“陽澄湖大閘蟹”地理標志產品實施了原產地保護。值得注意的是, 陽澄湖蟹年產量僅2×106kg, 但市面上銷售標稱為陽澄湖的蟹量卻遠超陽澄湖蟹的實際產量[8]。假冒陽澄湖蟹進行不法銷售的情況給原產大閘蟹的聲譽造成了嚴重損害。雖然近年來防偽技術不斷改進, 但假冒原產蟹的違法行為仍然很難防范[9]; 其中“洗澡蟹”(即將外地蟹移至陽澄湖水域養殖數日, 甚至過水后即冒充“陽澄湖大閘蟹”銷售)是最為主要的假冒方式[10,11]。但迄今尚無針對“洗澡蟹”和原產蟹形態及品質異同的比較研究。本研究嘗試運用幾何形態分析法, 利用陽澄湖實地養殖的原產蟹(從扣蟹一直養到成蟹)及養成于其他湖泊產地, 被筆者特意引入到前者養殖水域“洗澡”的大閘蟹; 對其在相同水域“洗澡”1個月始末時的外部形態進行比較研究, 以期為確認幾何形態學特征在區別陽澄湖原產和“洗澡”蟹上的應用潛力提供理論依據和數據支撐。筆者曾對中華絨螯蟹的背甲和腹甲進行過前期的幾何形態分析法研究效果的比對。由于對于判別不同產地中華絨螯蟹準確率而言, 腹甲的效果較差(雌、雄形狀相異, 前后者平均分別為83.75%、98.75%), 而背甲的效果很好(雌、雄形狀相似, 兩者均達100%)[12]; 故本研究中只使用背甲來進行上述比較研究。
1 材料與方法
1.1 實驗材料
中華絨螯蟹的陽澄湖成蟹樣本分別于2018年9月和10月采自陽澄湖圍網養殖區(31°26′N, 120°49′E)。非陽澄湖成蟹樣本于同年9月采集于江蘇省同屬長江流域的另一湖泊圍網養殖區。所采的非陽澄湖圍網養殖樣本一部分個體運回實驗室后進行“洗澡”前的形態測量, 剩余個體帶放至陽澄湖上述原產蟹相同圍網養殖區內“洗澡”養殖1個月后, 與10月陽澄湖原產樣本蟹同日采集, 進行“洗澡”后的形態測量。外來“洗澡”蟹兩類中華絨螯蟹和原產蟹所使用的苗種均來自于江蘇南通如東大閘蟹的苗種場(親本均為長江水系中華絨螯蟹)。“洗澡”始末時兩類蟹各選取規格相近的中華絨螯蟹20只(♀、♂各10只)活體進行上述形態學比較測定(表1),所有陽澄湖原產蟹均符合國標GB/T19957-2005[13]所定義的原產蟹(即至少從扣蟹養到成蟹保持同一水域)要求。
1.2 地標點的建立與提取
在設定的高度使用數碼相機(Nikon COOLPIX P6000)對非陽澄湖9月的樣本、“洗澡”后的10月樣本和陽澄湖成蟹樣本進行拍攝。地標點的選擇需特征明顯且易識別, 主要可分為3類, 即以不同組織間交點為特征的Ⅰ型地標點, 以結構中凹陷或凸起點為特征的Ⅱ型地標點和以結構極值點為特征的Ⅲ型地標點[14]。利用tpsdig2軟件對拍攝并經過處理的背甲照片建立地標點(圖1和表2), 獲取地標點的x、y坐標值(2D), 建立地表點的數據文件。
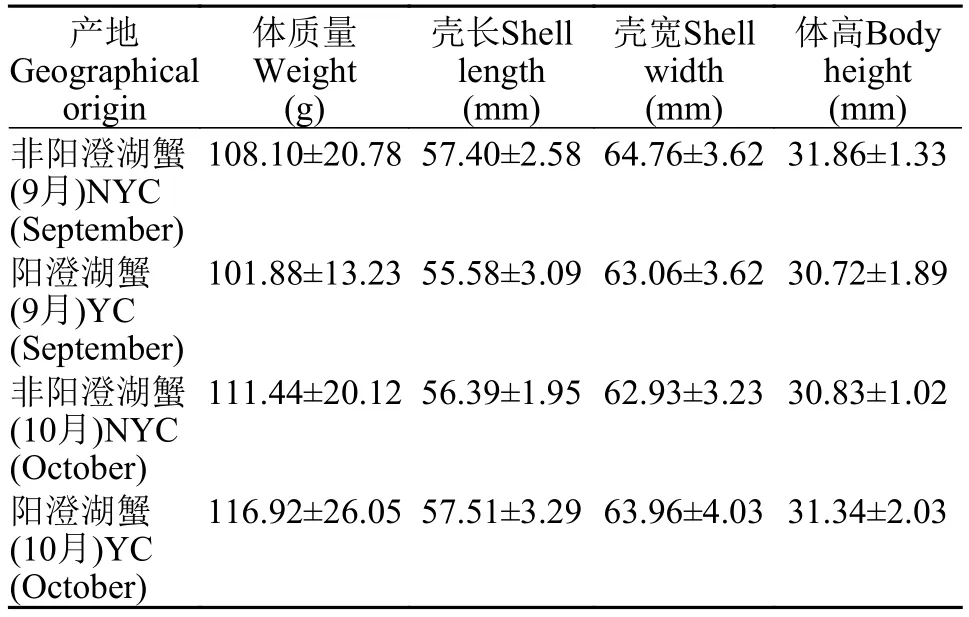
表1 本研究中中華絨螯蟹群體的形態測量信息Tab. 1 Morphometric information of E. sinensis populations in the present study (n=20, Mean±SD)
1.3 平均形與薄板樣條分析
地標點的有效性利用tpsSmall軟件的最小平方和法則回歸分析來檢驗。用tpsRelw進行普氏疊加等操作, 計算出地標點質心距離, 進而獲取平均型(Mean shape)、相對扭曲指數矩陣和數據報告。通過tpsRegr軟件的薄板樣條分析, 獲取成蟹背甲的網格圖, 以用于把握背甲的形態變化特征[12]。
1.4 判別分析
采用Bayes法基于各樣本的相對扭曲得分(Relative warps scores)實施逐步判別分析。用SPSS 23.0進行相關統計學分析。
2 結果
2.1 幾何形態分析
9月和10月中華絨螯蟹背甲地標點數據的切空間距離(Distance in tangent space,y軸)與普氏距離(Procruste distance,x軸)回歸系數分別為0.999830和0.999819, 均接近1, 說明本研究所選取的地標點是有效的, 可以應用于后續的幾何形態研究。
利用tpsRelw軟件在疊印過程中計算出所有樣本的平均形(圖2)。在對9月中華絨螯蟹背甲地標點數據進行的地標點法分析中, 貢獻率最高的地標點分別為11、12、16和17地標點, 共解釋了47.80%的形態差異, 其余31個地標點解釋了52.20%的形態差異。在對10月中華絨螯蟹進行的地標點法分析中, 貢獻率最高的地標點是16、17、18和19地標點, 共解釋了42.25%的形態差異, 其余31個地標點解釋了57.75%的形態差異。
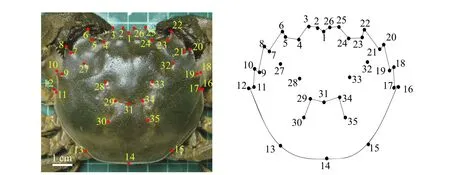
圖1 中華絨螯蟹背甲地標點位置圖Fig. 1 Landmark points for morphological measurements on the carapace of E. sinensis
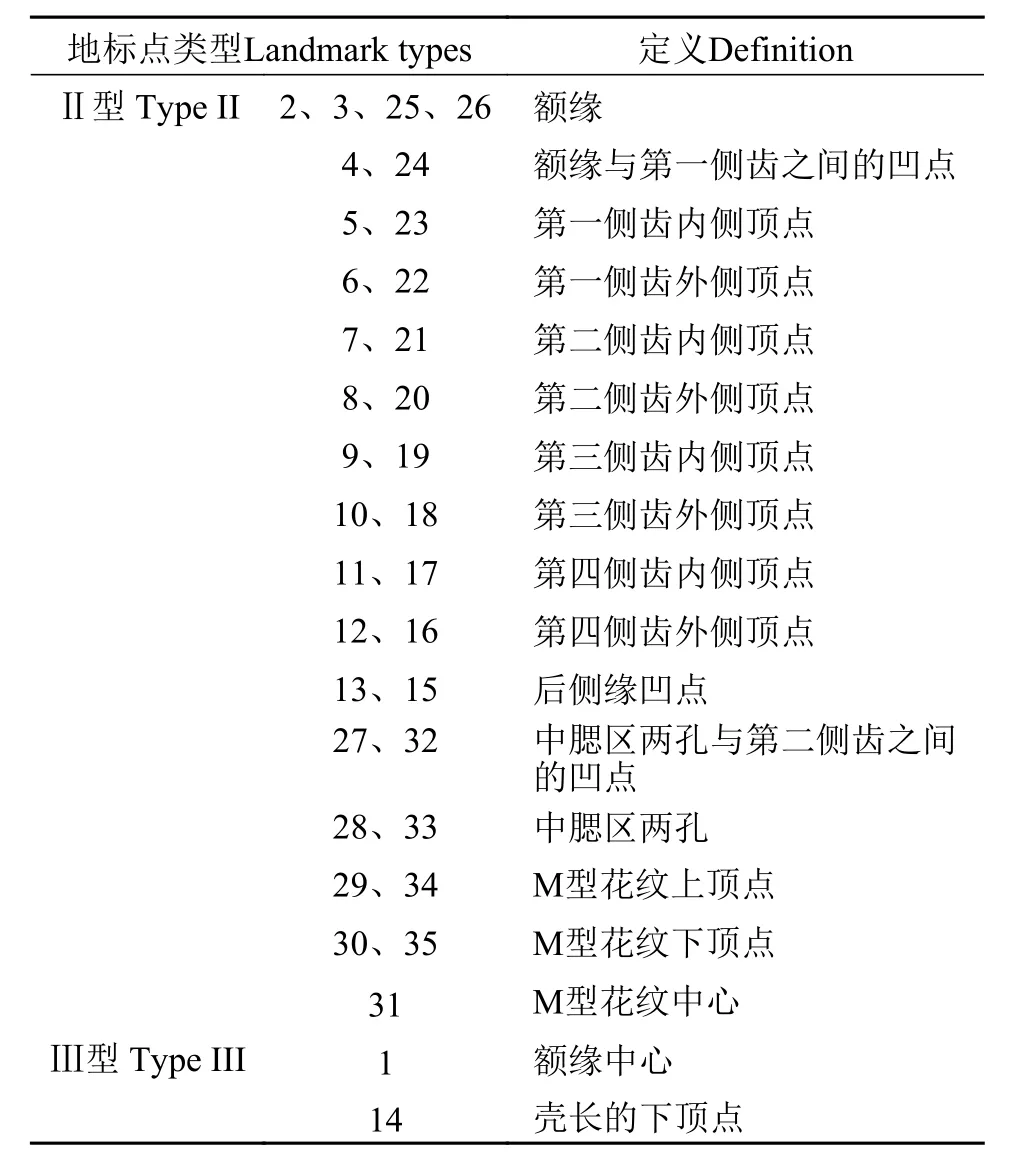
表2 地標點類型與定義Tab. 2 The landmark types and definition
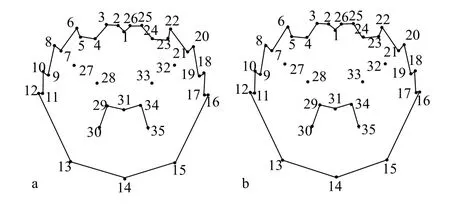
圖2 陽澄湖蟹和非陽澄湖蟹背甲的總體平均型Fig. 2 General mean types of the carapace for E. sinensis of Yangcheng Lake and Non-Yangcheng Lake origin
對9月的非陽澄湖和陽澄湖中華絨螯蟹背甲地標點數據進行相對扭曲主成分分析, 前3個主成分的總貢獻率為7.72%, 表明各因子線性關系弱, 降維效果差, 無法對總體進行有效的解釋。對10月非陽澄湖與陽澄湖中華絨螯蟹背甲地標點數據的相對扭曲主成分分析結果顯示, 前3個主成份的總貢獻率為7.72%, 降維效果差, 同樣無法對總體進行有效的解釋。
使用tpsRegr軟件將不同產地中華絨螯蟹的差異可視化, 進行網格變形分析后, 從網格圖可以發現, 兩個產地之間的中華絨螯蟹背甲差異沒有非常明顯, 所以所有結果均放大10倍進行比較與分析。
從9月份的非陽澄湖蟹與陽澄湖蟹的雄蟹背甲的網格圖(圖3)中可以發現, 非陽澄湖產中華絨螯蟹的第四側齒(11、12地標點)向外側擴展, 背甲后側緣的3個特征點(13、14、15地標點)向內側收縮, 陽澄湖蟹的背甲底部更加平緩。非陽澄湖蟹的M型花紋的頂點(29、30、34、35地標點)均向外擴張, 花紋更寬更長, 其余地標點沒有顯著的差異。
從9月份的非陽澄湖蟹與陽澄湖蟹的雌蟹背甲的網格圖(圖3)中可以發現, 非陽澄湖蟹額緣的端點(1、2、26地標點)向兩邊伸展, 頂部更寬。各側齒之間沒有明顯的差異, 非陽澄湖蟹后側緣3個特征點(13、14、15地標點)均向內收縮, M型花紋左下角與右下角的頂點(30、35地標點)均向外伸展,花紋更寬。其余地標點沒有顯著的差異。
從10月份的“洗澡蟹”與陽澄湖蟹的雄蟹背甲網格圖(圖4)中可以發現, “洗澡蟹”的第一側齒(5、6地標點)向內收縮, 第四側齒(11、12地標點)向外伸展。M型花紋的頂點(29、30、34、35地標點)均向內收縮, 非陽澄湖蟹的M型花紋更短更窄。其余地標點沒有顯著的差異。
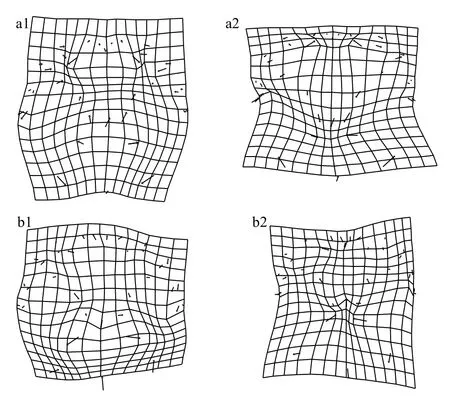
圖3 九月中華絨螯蟹背甲網格變形圖與變異可視化(變異擴大10倍)Fig. 3 Grid deformation and variation visualization of the carapace for E. sinensis in September (variation enlarged 10 times)
從10月份的“洗澡蟹”與陽澄湖蟹的雌蟹背甲網格圖(圖4)中可以發現, 陽澄湖蟹的第四側齒(11、12地標點)均向內收縮, M型花紋(29、30地標點)左側頂點向外伸展, M型花紋更寬。后側緣3個特征點(13、14、15地標點)均向外擴展, 底部更加的平緩。其余地標點沒有顯著的差異。
從圖3和圖4中華絨螯蟹的差異可視化比較可以看出, “洗澡”前后陽澄湖蟹與本研究所使用非陽澄湖蟹形態差異均主要集中于第四側齒、M型花紋和后側緣處。

圖4 十月中華絨螯蟹背甲網格變形圖與變異可視化(變異擴大10倍)Fig. 4 Grid deformation and variation visualization of the carapace for E. sinensis in October (variation enlarged 10 times)
非陽澄湖蟹“洗澡”前后雄蟹與雌蟹的背甲形態均發生了顯著變化; 且導致差異的主要地標點趨勢相同, 即M型花紋的頂點(29、30、34、35地標點)均向內收縮, M型花紋均變得更短更窄。后側緣3個特征點(13、14、15地標點)向外側略微擴展, 底部變得平緩。
2.2 判別分析
通過對9月與10月的陽澄湖蟹與非陽澄湖蟹的逐步判別分析(圖5和表3), 可以發現, 對9月與10月陽澄湖蟹與非陽澄湖蟹的判別準確率可以達到100%正確率。對同一產地的雄蟹與雌蟹也可以100%準確的判別。9月與10月的交互驗證正確率均可以達到95%。在交互驗證中, 9月的非陽澄湖雄蟹有1只誤判入陽澄湖雄蟹, 有1只非陽澄湖雌蟹誤判入陽澄湖雄蟹。10月的交互驗證結果中有兩只陽澄湖雄蟹誤判入非陽澄湖雄蟹中。
通過對陽澄湖與非陽澄湖“洗澡”前后的雄蟹和雌蟹的判別分析散點圖可以發現(圖6), 陽澄湖雄蟹在9月與10月已經趨于穩定, 幾乎沒有發生變化。非陽澄湖中華絨螯蟹雄蟹 “洗澡”前后不僅自身的背甲形態變化較大, 而且與陽澄湖蟹的背甲相比也都存在較大差異。這些形態差異均可以準確的判別出來。對于雌蟹的判別分析散點圖中, 陽澄湖蟹變化也較小, 趨于穩定。非陽澄湖雌蟹在“洗澡”后的自身形態變化雖不如雄蟹的變化明顯; 但無論是“洗澡”前、還是后, 與陽澄湖原產蟹的差異均較大, 不會趨同。
3 討論
3.1 “洗澡蟹”形態差異分析
中華絨螯蟹的消費季節一般開始于每年9月,以10—11月為高峰。這段時間也正是“洗澡蟹”最嚴重的危害期。為不法利益最大化, 只有非陽澄湖產的成蟹才會被用作“洗澡蟹”。本研究中的兩類蟹在“洗澡”始末時均達前人報道的成蟹規格[15]。值得注意的是, 因為兩類蟹苗種來源相似、種質相同, 理論上其背甲的形態應該相像; 但實際上在“洗澡”開始前兩者之間背甲的形態就已經相異(圖5和圖6)。鄭朝臣等[12]在對長江水系的太湖、洪澤湖、高寶湖、長蕩湖、陽澄湖以及巴城、興化“泓膏”養殖水域和長江口崇明自然水域等地產中華絨螯蟹形態上差異的比較研究發現, 即使來源于種質特點相似(長江水系親本蟹在射陽所育大眼幼體或在南通/崇明所育扣蟹)的苗種, 在經過至少從扣蟹養到商品成蟹近1年的養殖或生長時間后, 上述8個產地蟹的背甲會出現顯著的產地差異性變化, 利用幾何形態分析法甚至可獲100%的產地判別準確率。因此可以想見, 成蟹規格的“洗澡蟹”與陽澄湖原產蟹在經過了從扣蟹養到成蟹長期的養殖期后,其背甲的形態應該也已形成了對兩產地水環境相應的適應性變化。Sarà等[16]通過地標點研究尖吻重牙鯛(Diplodus puntazzo)發現, 養殖于不同環境條件(養殖池單養、遠岸網箱單養、與金頭鯛(Sparus aurata) 養殖池混養)下, 魚的形態要在8個月后才有明顯的適應性變化。張秀霞等[17]通過地標點法對8個野生唐魚(Tanichthys albonubes)種群的研究發現, 長期相互獨立的生境條件和地理隔離使唐魚的頭部形態出現差異。由此可以看出, 魚、蟹要改變原有身體形態很可能需要長期棲息(如>8個月)于不同產地生境后才會發生。本研究中“洗澡蟹”僅1個月短期轉換生境養殖于陽澄湖中, 雖然可見適應性的變化, 但其原有產地而形成的形態特征并未消失,仍可通過幾何形態分析其差異與陽澄湖原產蟹進行準確地區分。
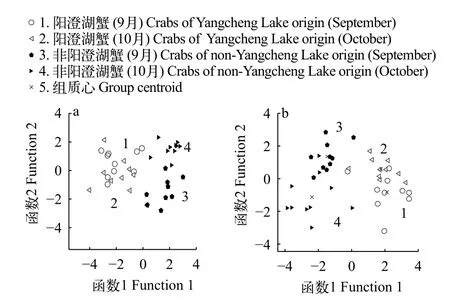
圖6 基于背甲的中華絨螯蟹判別分析散點圖Fig. 6 The discriminate analysis plots of E. sinensis by carapace geometrical morphometry
3.2 地標點的選擇及其對背甲形態判別效果的影響
在對9月中華絨螯蟹背甲的分析中, 貢獻率最高的地標點分別為11、12、16和17地標點, 共解釋了47.80%的形態差異, 其余31個地標點解釋了
52.20%的形態差異。在對10月中華絨螯蟹進行的分析中, 貢獻率最高的地標點是16、17、18和19地標點, 共解釋了42.25%的形態差異, 其余31個地標點解釋了57.75%的形態差異。這些貢獻率較高的地標點均為各側齒內外側頂點, 屬于Ⅱ型地標點。結合本研究結果來看, 所選地標點能夠解釋不同中華絨螯蟹群體形態學上的主要變異信息(判別準確率達100%, 交互驗證結果也高于80%)。在相關研究中, 侯剛等[18]使用地標點法對紅棘金線魚(Nemipterus nemurus)、金線魚(N. virgatus)、深水金線魚(N. bathybius)和日本金線魚(N. japonicus)矢耳石進行研究時, 判別準確率可以達到了91.3%, 其中Ⅰ型與Ⅱ型地標點貢獻率較高。與之相比, 馮波等[19]使用地標點法與傳統形態學方法對不同種群短吻鲾(Leiognathus brevirostris)進行區分的時候, 發現地標點法的區分效果不如傳統形態學方法, 通過分析發現其選取的Ⅲ型地標點較多且解釋了主要的形態學變異信息。由此可見在利用地標點法進行不同群體形態學差異比較時, 以Ⅰ型和Ⅱ型地標點為優, 同時為了避免造成信息缺失, 少量選擇界限較為明顯且不易受到主觀因素影響的Ⅲ型地標點(如本研究中地標點14)。
3.3 背甲形態判別“洗澡蟹”的潛力
幾何形態分析是形態測量學的一種重要方法[20,21]。目前已廣泛應用于招潮蟹(Genus uca)[22]、藍黑鯪(Labeo calbasu)[23]、兔脂鯉(Leporinus cylindriformis)[24]和梭子蟹(Portunus pelagicus)[25]等生物的形態的種內、種間差異性研究或種群判別的研究中。迄今對于不同產地的中華絨螯蟹的判別,已有從形態學[26—28]、分子生物學[29]和生理生化指標[30—32]等不同角度的嘗試, 其中以幾何形態分析[12]、元素“指紋”[33]和電子舌[34]所獲的產地判別準確率較高, 尤其是幾何形態分析具有可進行非致死測定且判別準確的優點。在筆者實驗室的前期研究中,鄭朝臣等[12]通過地標點法對活體中華絨螯蟹的背甲與腹甲進行分析, 準確地判別出了長江水系8個不同湖泊產地的中華絨螯蟹, 其中對雄蟹背甲、雄蟹腹甲、雌蟹背甲、雌蟹腹甲的判別準確率分別達到了100%、98.75%、100%和83.75%。相比于傳統的形態測量較低的判別準確率[35], 不僅在判別準確率上有了明顯的提升, 同時對于背甲的判別準確率還達到了100%, 平均的判別準確率達到了運用元素“指紋”與同位素的的[33]判別準確率。
本研究運用地標點法對“洗澡蟹”與陽澄湖原產蟹的背甲進行判別。判別結果顯示(圖5), 在“洗澡”前, 通過對相對扭曲得分進行逐步判別分析, 對非陽澄湖蟹和陽澄湖蟹的判別準確率可以達到100%, 交互驗證的準確率依然可以達到95%。在將非陽澄湖的中華絨螯蟹于陽澄湖中進行1個月的“洗澡”之后, 再次通過地標點法與同月陽澄湖產中華絨螯蟹進行甄別, 逐步判別準確率仍然可以達到100%, 并且交互驗證準確率依舊可以達到95%, 有效證明了于陽澄湖中“洗澡”的非陽澄湖產的中華絨螯蟹即使“洗澡”1個月時間, 也無法“洗白”變成陽澄湖原產蟹的背甲形態。
本研究通過幾何形態學方法對“洗澡蟹”與陽澄湖原產蟹進行了準確地鑒別, 有效地證明了即使1個月“洗澡”也無法有效地改變非陽澄湖產與陽澄湖原產蟹間的形態差異。幾何形態學方法的判別準確潛力非常高并且可以進行差異可視化分析, 并且還擁有快速, 方便和非致死的優點。在今后的進一步研究中, 擬嘗試將其應用于中華絨螯蟹生長過程中以探索中華絨螯蟹在某固定的環境條件(特別是水質、水環境微量元素含量、水草豐度、流速及水體交換量、養殖密度等)下達到背甲形態穩定所需要的時間, 并且通過形態差異可視化來深入探索中華絨螯蟹生長過程中具體的形態變化, 以便嘗試開發可操作推廣的基于幾何形態分析的背甲識別技術, 為保護陽澄湖原產中華絨螯蟹提供一條創新性的途徑。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
