RseA基因影響恥垢分枝桿菌藥物敏感性的生物信息學分析與初步驗證
2021-03-24 05:54:26熊雨菡李智穎陳俊伊唐佳玲何永林
中國人獸共患病學報 2021年2期
關鍵詞:分析
熊雨菡,李智穎,盧 楠,陳俊伊,唐佳玲,徐 蕾,楊 春,何永林
全球每年約有1 000萬新發結核病例,其中有150萬人死于結核病(Tuberculosis,TB)[1]。而結核分枝桿菌多藥耐藥(MDR)和廣泛耐藥(XDR)突變株的出現和傳播將阻礙結核病的治療[2]。2018年,全球預計有484 000人患上了利福平(結核病最有效的一線藥物)耐藥結核病(RR-TB),其中78%患有多藥耐藥結核(MDR-TB)[1],而結核分枝桿菌變得對藥物治療不敏感可能是由于其進入了非復制狀態[3]。有研究表明,在大多數個體中,最初的感染轉變為無癥狀的潛伏狀態,結核分枝桿菌以無復制的持續狀態存在于宿主肉芽腫中,且代謝活性低[4-5]。導致這種生理狀態建立的不利環境因素包括低氧、營養不良、pH低以及宿主免疫反應產生的氧化和亞硝化應激[5]。這種適應主要歸因于RNA聚合酶參與的能夠調節其復雜基因組表達的轉錄調控網絡[6]。σ因子與RNA聚合酶全酶結合,提供其對特定啟動子的特異性,并在調節基因表達和適應原核生物的壓力中起關鍵作用[7]。結核分枝桿菌基因組編碼13個σ因子,輔助σ因子σE是結核分枝桿菌中最典型的σ因子之一,它對于在不同壓力環境下(包括表面和氧化應激條件)的生存至關重要[8]。我們前期研究發現,σE可通過促進恥垢分枝桿菌進入持留狀態影響抗結核藥物的敏感性[9]。
近來,抑制σ因子功能的抗σ因子也受到越來越多的關注。結核分枝桿菌的抗σE因子RseA由Rv1222基因編碼。恥垢分枝桿菌是一種在安全性、生長特性和遺傳易感性方面用于分枝桿菌研究的模式生物,已被廣泛用于抗生素的作用和耐藥性研究[10]。恥垢分枝桿菌的RseA由基因MSMEG_5071編碼。RseA編碼基因緊鄰下游σE,已被證明是可直接調控σE的特異性抗σE因子[11],而至今針對RseA在抗結核藥物敏感性的機制研究較少。因此,本實驗構建了重組恥垢分枝桿菌pMV261-MSMEG-5071/MS,通過轉錄組測序技術和生物信息學方法篩選出差異表達基因(differentially expression genes,DEGs),并對其進行了基因本體論(Gene Ontology,GO)和京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析,構建了蛋白-蛋白相互作用(protein-protein interaction,PPI)網絡,通過MCODE插件篩選出可能受RseA調控的基因及其可能的分子機制并進行初步驗證,為耐藥結核病的治療提供新思路。
1 材料與方法
1.1實驗材料 質粒pMV261、E.coliDH5α和恥垢分枝桿菌(Mycobacteriumsegmatismc2155,MS)和RpfE蛋白由實驗室保存。HindⅢ、BamH Ⅰ和T4連接酶(Thermo Fisher Scientific),DAB顯色試劑盒(康為世紀),卡那霉素(Genview),膠回收試劑盒、質粒提取試劑盒、蛋白Marker(Takara),小鼠抗組氨酸單克隆抗體(Bioworld Technology),HRP標記的羊抗兔IgG(美國Signalway Antibody),SDS-PAGE凝膠配置試劑盒(EpiZye),異煙肼、利福平、乙胺丁醇、左氧氟沙星、環丙沙星、鏈霉素(北京睿博興科)。
1.2方 法
1.2.1重組恥垢分枝桿菌的構建 根據NCBI GenBank(基因序列號NC_008596.1)中恥垢分枝桿菌MSMEG_5071的基因序列設計引物。以MS基因組為模板進行PCR擴增,將PCR產物和質粒pMV261分別經BamHⅠ/HindⅢ酶切并用T4連接酶連接后,轉化至E.coliDH5α感受態細胞中,涂布在含有卡那霉素的LB固體培養基上,長出單菌落后,挑取單菌落培養并測序驗證。將構建成功的pMV261-MSMEG_5071重組質粒經2.5 KV、4 ms電轉至恥垢分枝桿菌感受態中,涂布在含有卡那霉素的7H10固體培養基上,長出單菌落后,挑取單菌落培養以獲得重組恥垢分枝桿菌pMV261-MSMEG_5071/MS。
1.2.2Western blot驗證蛋白RseA的表達 將構建成功的重組恥垢分枝桿菌pMV261-MSMEG_5071/MS接種于含有卡拉霉素的7H9培養基中,37 ℃培養至OD600為0.6~0.8,分別于40 ℃、42 ℃、44 ℃和46 ℃熱誘導3 h后,37 ℃,160 r/min培養4 h,離心收集菌體后,于-80 ℃反復凍融3次,加入無菌PBS混勻,超聲碎菌,離心后分別收集上清與沉淀。將樣品經金屬浴100 ℃ 10 min變性,通過Western blot驗證。
1.2.3樣本準備及轉錄組測序 將重組菌接種于含有卡那霉素的7H9培養基中,培養至OD600為0.8,42 ℃熱誘導3 h,加入10 μg /mL EMB,37 ℃,180 r/min培養48 h,離心收集菌體后,PBS洗滌3次,置于液氮盒中,送至上海美吉生物醫藥科技有限公司進行轉錄組學測序和生物信息學分析。
1.2.4測序數據質控 通過Illumina Hiseq 測序獲得的原始圖像經base calling轉換為序列數據,即原始數據raw data。使用Sickle和SeqPrep質控軟件除去reads中的adapter序列、5′端含有非A、G、C、T的堿基和含N的比例達到10%的reads,修剪測序質量較低的reads末端 (測序質量值小于Q20),棄置經去adapter及質量修剪后長度小于25 bp的小片段,得到高質量reads,即clean data,并對數據進行質量評估。通過Diamond分析軟件進行核糖體RNA污染率評估,若rRNA含量低于15%,則可進行后續分析。
1.2.5DEGs分析 使用Bowtie2軟件計算出每組中各個基因的表達量,通過DEGseq分析RseA基因過表達恥垢分枝桿菌組及空質粒對照組的DEGs。篩選差異倍數(fold change)≥2且P≤0.001的基因為顯著DEGs。
1.2.6GO功能富集和KEGG信號通路分析 GO是一個在生物信息學領域中廣泛使用的本體,是實現統一基因相關數據、進行數據轉換和開展數據挖掘的基礎,包括分子功能(molecular function,MF)、細胞組分(cellular components,CC)和生物學過程(biological process,BP)。KEGG是從分子水平信息了解高級功能和生物系統的實用程序數據庫資源。本實驗通過DAVID6.8對DEGs進行GO分析及KEGG分析。P<0.05為功能和通路具有統計學意義。
1.2.7PPI網絡的構建 STRING(https://string-db.org/)是可用于PPI網絡功能富集分析的在線數據庫之一。在STRING中輸入DEGs,以得到其PPI關系(sore>0.9),通過Cytoscape 3.7.2軟件構建PPI網絡。通過復雜分子檢測(molecular complex detection,MCODE)插件篩選出最重要模塊,對模塊進行GO功能富集分析和KEGG信號通路分析,P<0.05差異具有統計學意義。并篩選出模塊中連接度最高的前8個hub基因。
1.2.8復蘇指數的測定 將重組菌培養至生長對數期,調節OD600至0.5,用含有卡那霉素的7H9液體培養基洗滌2次,離心后分別用含有100 μg /mL異煙肼(isoniazid,INH)、25 μg /mL利福平(rifampin,RIF)、10 μg /mL乙胺丁醇(ethambutol,EMB)、10 μg /mL左氧氟沙星(levofloxacin,LVFX)、10 μg /mL環丙沙星和6.4 μg /mL鏈霉素(streptomycin,STRE)的7H9培養基重懸,37 ℃,180 r/min培養48 h。離心棄上清,用7H9培養基重懸后分為2組。一組等倍梯度稀釋至106,所有濃度的菌液各吸取10 μL加至7H9固體培養基上,37 ℃培養3 d,計數;另一組用含有 20 ng/mL RpfE(可以促進非復制的持留菌復蘇生長[12])的7H9液體培養基等倍梯度稀釋至106,每濃度菌液吸取200 μL加至96孔板,并設置復孔,96孔板四周用200 μL 7H9液體培養基封板,于37 ℃培養2周后,觀察并記錄陽性細菌孔,查表得到最大可能數(Most probable number,MPN)。通過log10(MPN)-log10(CFU)計算得出的復蘇指數(Resuscitation Index,RI)來表示其復蘇能力[13]。
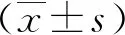
2 結 果
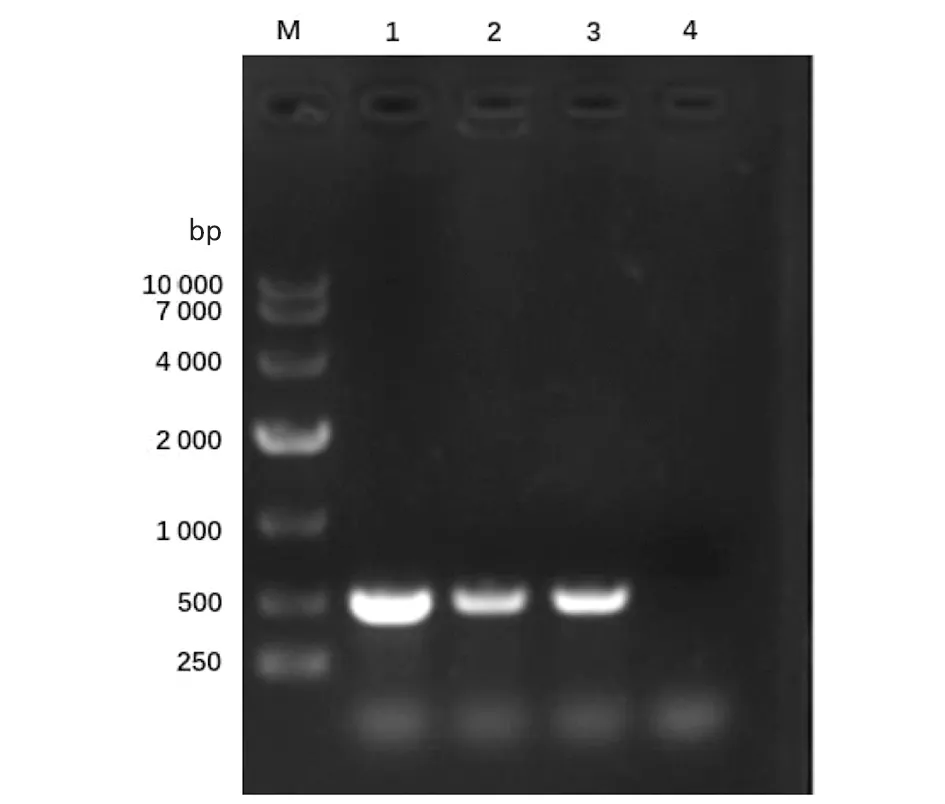
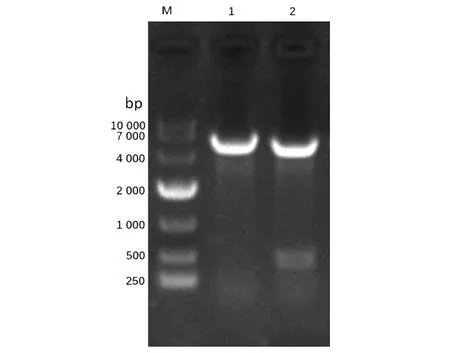
2.1重組恥垢分枝桿菌pMV261-MSMEG_5071/MS的構建及驗證 以MS基因組為模板,擴增得到的PCR產物大小約為400 bp,與預期大小一致(見圖1)。將構建成功的重組質粒pMV261-MSMEG_5071經BamHⅠ/HindⅢ酶切后,酶切片段大小與預期一致(圖2)。將重組質粒pMV261-MSMEG_5071電轉入恥垢分枝桿菌,40 ℃、42 ℃、44 ℃和46 ℃誘導表達,超聲碎菌處理,離心后,收集上清與沉淀,經Western blot驗證,獲得分子量為17 kD左右的條帶,與預期大小一致(圖3)。
2.2兩組間DEGs分析結果 轉錄組測序結果顯示,兩組間共篩選出2 403個DEGs,其中2 335個上調基因,68個下調基因。將2 403個DEGs繪制成火山圖,其中紅色代表上調基因,綠色代表下調基因(圖4)。
M.DNA marker;1-3.MSMEG-5071;PCR product is about 400bp;4.H2O as a control.
M.DNA Marker;1.pMV261-MSMEG-5071;2.Double digestion
M.Marker;1-8.supernatant and precipitation of pMV261-MSMEG-5071 lysate induced at 40 ℃,42 ℃,44 ℃,46 ℃;9.No-load strain induced at 42 ℃ for 3 h.
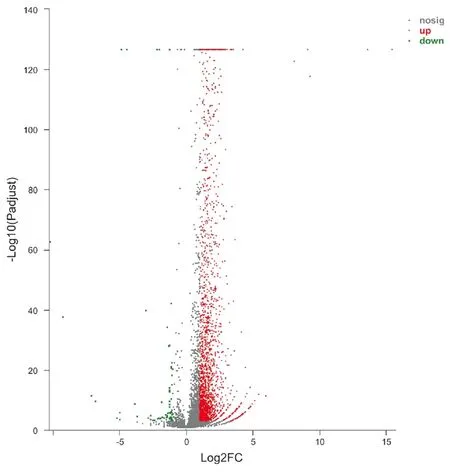
圖4 DEGs火山圖
2.3GO功能富集和KEGG信號通路分析 將篩選出的DEGs進行GO分析,發現DEGs共參與了155個生物學過程(P<0.05),其中MF主要富集在轉錄因子活性,序列特異性DNA結合、核酸結合轉錄因子活性和核酸結合;CC主要富集在細胞內,轉移酶復合物,轉移含磷基團;BP主要富集在氮化合物代謝過程的調控、RNA代謝過程的調控和細胞代謝過程的調控等過程。GO富集的部分生物學過程見圖5。KEGG分析顯示,DEGs參與了8個KEGG信號通路(P<0.05),主要富集在萜類骨架的生物合成、果糖和甘露糖代謝和同源重組等通路(表1)。
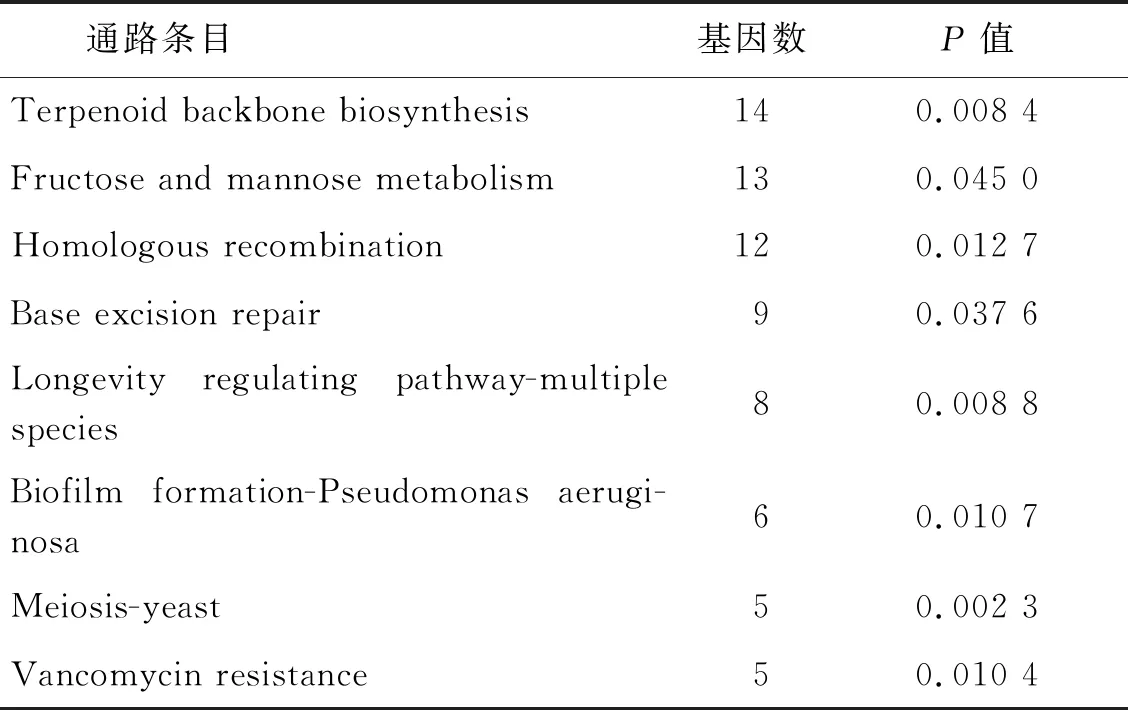
表1 DEGs的KEGG通路分析
2.4PPI網絡 通過STRING在線分析軟件獲得DEGs的PPI關系,構建PPI網絡。結果顯示,共有1 136條邊緣數和594個hub基因(圖6)。使用插件MCODE分析出的最重要模塊含有16個hub基因和119條邊緣數(圖7)。對這16個hub基因進行GO分析和KEGG分析。GO分析結果顯示,這16個hub基因主要富集在翻譯、核糖體的結構成分和核糖體等功能(表2)。KEGG分析結果表明,其主要富集在核糖體通路(表3)。
①:P<0.001

表2 模塊內基因的GO富集分析

表3 模塊內基因的KEGG通路分析

圖6 PPI網絡圖

圖7 PPI中MCODE分析得到的最高分子模塊
2.5RseA過表達對藥物誘導的持留菌復蘇情況的影響 分別用高濃度藥物INH、RIF、EMB、LVFX、CPFX和STRE處理細菌48 h,誘導細菌處于非復制狀態,用含有20 ng/mL RpfE蛋白的7H9促進持留菌復蘇,觀察兩組菌復蘇情況。INH、RIF、EMB、LVFX和CPFX 5種藥物處理后,MSMEG_5071過表達組復蘇指數明顯低于對照組(圖8),差異有統計學意義(tINH=4.37,tRIF=7.26,tEMB=6.75,tLVF=11.13,tCPFX=8.24,tSTRE=0.02;P<0.05,PINH=0.011 966,PRIF=0.001 912,PEMB=0.002 512,PLVF=0.000 371,PCPFX=0.001 185,PSTRE=0.987)。
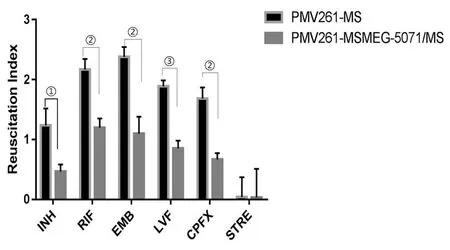
①P<0.05,②P<0.01,③P<0.001
3 討 論
本研究通過生物信息學分析,比較了pMV261-MSMEG_5071和含有空質粒pMV261的恥垢分枝桿菌的轉錄組測序結果,共篩選出2 403個DEGs,其中上調基因2 335個,下調基因68個。同時,分析了這些基因的PPI關系,構建了PPI網絡并通過MCODE插件分析,篩選了RseA影響恥垢分枝桿菌藥物敏感性的關鍵靶向基因。其中,篩選出的前8個基因,rpsG、rplM、rpsO、rpsT、rpmH、rplS、rpsJ、rplT可能在RseA影響恥垢分枝桿菌藥物敏感性過程中具有重要作用。
rpsG編碼核糖體蛋白S7,該蛋白位于核糖體小亞基(30S)頭部,是唯一與mRNA交聯的顯著靶標,也是通過翻譯反饋機制控制核糖體蛋白合成的主要調控元件之一,對于30S亞基的組裝至關重要[14-16]。rplM編碼核糖體蛋白L13,是核糖體大亞基(50S)組裝中的必需蛋白[16]。有研究發現,恥垢分枝桿菌MSMEG_1556(rplM)敲除株無法在42 ℃條件下生長,由此證明了MSMEG_1556(rplM)對于恥垢分枝桿菌的生長至關重要[17]。rpsO編碼核糖體蛋白S15,是位于30S亞基平臺上的唯一一級結合蛋白。S15與16S rRNA的預先結合是二級和三級核糖體蛋白(r蛋白)S6、S18、S11、S21與新興復合物結合所必需的[18]。由此證明,S15參與了亞基締合并在30S亞基組裝中發揮了重要作用[19]。rpsT編碼核糖體30S亞基蛋白S20,該蛋白直接與16S rRNA結合,并獨立于其他蛋白,被定義為一級結合蛋白,由此表明S20在30S亞基的組裝中起著關鍵作用[20]。并有研究表明,缺乏S20的菌株生長速度明顯降低[21]。rpmH編碼核糖體大亞基蛋白L34,L34的缺乏會導致70S核糖體形成的嚴重缺陷和生長速率的降低[22]。rplS編碼核糖體50S蛋白L19,L19可以抵御抗體,并且與氯霉素的結合有關[23]。rpsJ編碼核糖體30S亞基蛋白S10,有實驗表明,該蛋白是參與轉錄抗終止作用的細胞成分之一[24]。rplT編碼核糖體50S蛋白L20,參與50S亞基的早期組裝步驟以及自身基因表達的反饋控制[25]。
PPI網絡模塊分析顯示,RseA過表達導致多種核糖體蛋白異常表達,核糖體的結構蛋白合成和翻譯通路富集。核糖體是核糖核蛋白復合物,由具有21個蛋白和16S rRNA的小亞基(30S)以及具有36個蛋白,23S rRNA和5S rRNA的大亞基(50S)組成,已成功被多種抗生素所靶向[26]。核糖體執行蛋白質生物合成的關鍵功能,主要分為翻譯起始、延伸、終止和再循環4個階段,而翻譯是適應某些環境條件(例如饑餓,物理或化學壓力,尤其是休眠)的主要監管目標[27]。因此,推測RseA過表達后,主要通過涉及核糖體的翻譯過程影響恥垢分枝桿菌對環境的適應能力,從而影響恥垢分枝桿菌的藥物敏感性。
我們采用復蘇指數來衡量持留菌的數量多少。使用高濃度藥物INH、RIF、EMB、LVFX、CPFX和STRE處理細菌,使其處于非復制的持留狀態,通過復蘇促進因子(RpfE)復蘇后,計數最大可能數MPN,即包括持留菌在內的所有活菌;同時計數未通過RpfE處理的菌落CFU,即不包含持留菌的活菌,兩者之差即為復蘇指數RI,以此來反應持留菌的多少。實驗結果觀察到,INH、RIF、EMB、LVFX、CPFX藥物作用后,RseA過表達組的復蘇指數明顯低于對照組,即RseA過表達可能導致持留菌減少;STRE作用后,RseA過表達組與對照組無差異。而STRE是一種氨基糖苷類抗生素,通過靶向核糖體,引起遺傳基因誤讀干擾翻譯啟動,抑制細菌蛋白質合成,從而殺滅細菌或抑制細菌生長[28]。由此可見,過表達RseA可能影響了核糖體通路,從而影響恥垢分枝桿菌對藥物的敏感性。
綜上所述,本研究通過生物信息學分析,發現RseA在恥垢分枝桿菌藥物敏感性的影響中發揮了重要作用,為RseA以后在結核分枝桿菌的治療應用提供了新的思路,后續將開展大量的基礎研究對此結果進行進一步的驗證。
利益沖突:無
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
