閩楠葉斑病病原菌的分離鑒定及其生物學特性
2021-09-22 01:23:02姚英,于存
農技服務 2021年6期
關鍵詞:生長
姚 英, 于 存
(貴州大學 林學院, 貴州 貴陽 550025)
閩楠(Phoebebournei)俗稱楠木,屬樟科(Lauraceae)楠屬(Phoebe)常綠大喬木,是我國特有物種,為國家二級保護植物,分布于福建、江西、湖南、廣西、廣東、貴州、浙江和湖北等地[1]。其材質通直并帶有特殊香味,紋理精美,廣泛用于建筑、家具、雕刻行業和精密木模的制作。同時,楠木冠大蔭濃,樹姿優美,是優良的園林綠化樹種[2]。此外,閩楠還有凈化空氣、隔音降噪和殺菌驅蟲等生態功能,廣泛用于香料提取及化妝品研發領域[3]。由于野生資源過渡采伐,導致楠木瀕臨滅絕,現已開始大量人工栽植[4-5],但其種植過程中常伴有多種病蟲害的發生,尤其是葉斑病已成為閩楠葉部的主要病害之一,且遍布各閩楠育苗基地及幼齡林分布區。閩楠葉斑病染病初期受害部位出現紅褐色圓形斑點,向外擴展,由許多小病斑塊融合成不規則的大斑,最后導致葉片退綠,嚴重致其死亡。炭疽菌屬(Colletotrichumspp.)是主要的植物致病菌和病原菌,分布廣、寄主多,可危害谷物、水果蔬菜和核桃等多種農林植物,對農林業生產威脅極大[6]。膠孢炭疽菌(Colletotrichumgloeosporioides)是炭疽菌屬的重要種類,可引起橡膠、芒果和香蕉等多種植物炭疽病[7],C.gloeosporioides具有集合種和復合種的特點,種內菌株間變異大,有數個專化型或生理小種[8]。目前,在閩楠上已發現小孢擬盤多毛孢(Pestalotiopsismicrospora)可引起葉斑病[9]。筆者在調查閩楠病害時發現,除P.microspora可引起閩楠葉斑外,還有其他病原菌可引起閩楠葉斑病。因此,探明引起某一植物病害的膠孢炭疽菌生物學特性,明確其引發病規律和發病特點,對該原菌引起的植物病害的防治具有重要意義。從貴州省都勻市螺絲殼自然保護區采集閩楠典型葉斑病病葉,對其進行病原菌的分離與鑒定,探明其生物學特性,以期為閩楠葉斑病的防治提供科學依據。
1 材料與方法
1.1 材料
閩楠典型葉斑病病葉樣品共計30片,采自貴州省都勻市螺絲殼自然保護區。
1.2 方法
1.2.1 病原菌的分離 采用常規組織分離法分離閩楠葉斑病病原菌,首先用1%次氯酸鈉進行葉片表面消毒3~5 min,后用無菌水清洗3~5次,無菌條件下,剪取病健交界處約3 mm×3 mm的組織放置在PDA平皿,每個平皿放置3塊病組織,每個葉片3次重復,30個葉片共90個處理。之后將樣品于(25±2) ℃條件下培養,當樣品邊緣生長出菌絲時及時挑取邊緣菌絲尖端轉移至新的PDA平皿中進行菌株純化,直至獲得純化菌株。獲取真菌菌株后根據菌落、菌絲和孢子特征進行合并,確定主要種類后統計不同真菌的分離率。
分離率=某一真菌分離個數/所有真菌分離個數×100%
1.2.2 病原菌的致病性測定 隨機選取分離得到的相同種真菌菌株5株按照柯赫氏法進行離體葉片的回接。將20年生閩楠健康葉經75%酒精表面消毒后,用無菌昆蟲針輕輕刺傷表面,用打孔器截取直徑為0.5 cm真菌菌餅置于刺傷部位,用濕脫脂棉保濕,25℃恒溫培養10 d,觀察不同真菌是否引起閩楠葉片葉斑的癥狀。選擇回接后產生明顯葉斑病的葉片,由發病的病健交界處,利用組織分離法再次分離病原菌,利用形態學方法比較再分離菌株與原接種菌株形態特征是否一致,以上每個真菌離體回接20片葉。
1.2.3 病原菌的鑒定
1) 形態學鑒定。將回接引起閩楠葉斑的病原菌接種至PDA平板上,于25℃暗培養5 d,培養過程中觀察菌落顏色、形狀、氣生菌絲的疏密程度等形態特征,并采用插片法獲取病原菌菌絲和孢子,觀察記錄菌絲和孢子的分支及大小,參照陸家云的方法進行病原菌的形態學鑒定[10]。
2) 分子學鑒定。在PDA平板上活化閩楠葉斑病病原菌,利用接種鉤刮下平板上生長的菌絲體用于病原菌基因組的提取。病原菌基因組提取采用CTAB法,用真菌ITS通用引物ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′)、ITS4(5′-TCCT-CCGCTTATTGATATGC-3′)擴增病原菌的目的基因序列。PCR反應體系(25 μL):10×Buffer 2.5 μL、2.5 mmol/L d NTP 2 μL、Taq 酶0.2 μL、模板 DNA 1 μL、25 μmol/L引物ITS1和ITS4各0.5 μL,加ddH2O至25 μL。PCR 擴增條件:95℃預變性4 min;94℃變性1 min,54℃退火1 min,72℃延伸1 min,共35個循環;最后72℃延伸10 min,4℃保存。擴增產物的純化和序列測定委托重慶擎科生物技術有限公司進行,所得到的核苷酸序列進行在線NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)同源性比對分析,應用MEGA 7.0中的臨近法(Neighbor-joining,NJ)構建病原菌ITS的系統進化樹,系統進化樹構建過程中采用Bootstrap法進行檢驗,重復計算1 000次后確定菌株的分類地位。
1.2.4 不同因素對病原菌菌絲體生長的影響
1) 培養基。供試培養基有水瓊脂(WA)培養基、馬鈴薯葡萄糖瓊脂(PDA)培養基、馬鈴薯蔗糖瓊脂(PSA)培養基、PDA+維生素B6(PDA+VB 6)培養基、PDA+楠木煎汁(Ph.bournei’s leaf agar,PBLA)培養基、孟加拉紅(Rose Bengal Agar)培養基、查彼培養基(Czapek Dox Agar) 、SNA培養基(Synthetic low nutrient Agar) 、燕麥瓊脂培養基(Oatmeal agar medium,OA)。將直徑為5 mm的菌餅接種至不同培養基,25℃暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑,5次重復。
2) 溫度。試驗溫度設15℃、20℃、25℃、30℃和35℃共5個梯度,取直徑為5 mm的菌餅接至PDA平板中央,分別在不同溫度條件下暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑,5次重復。
3) pH。用1 mol/L HCl溶液和1 mol/L NaOH溶液分別調節PDA培養基pH為5.0、6.0、7.0、8.0、9.0共5個梯度,取直徑5 mm 菌餅分別接種于不同pH的PDA培養基平板,25℃條件下暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑,5次重復。
4) 光照。取直徑5 mm菌餅接種至PDA平板中央后,分別置于全黑暗、12 h光暗交替和日光燈連續照射條件下,25℃暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑,5次重復。
5) 碳、氮源。以查彼培養基為基礎培養基,將等量的葡萄糖、麥芽糖、蔗糖、乳糖、可溶性淀粉、果糖為碳源,以不添加碳源為對照;將等量的蛋白胨、尿素、硝酸銨、酵母浸粉、牛肉膏為氮源,以不添加氮源作為空白對照;取直徑5 mm 菌餅分別接種于不同條件的培養基,25℃條件下暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑,5次重復。
6) 菌絲的致死溫度。設40℃、45℃、50℃、55℃和60℃共5個梯度,在無菌環境下,將無菌水加入5 mL的離心管中,取直徑5 mm菌餅放入離心管中并置于不同溫度的水浴鍋中水浴10 min,再接回PDA平板中,25℃條件下暗培養5 d,然后采用“十字交叉法”測定菌落生長直徑。5次重復。
1.3 數據處理
采用SPSS 20.0對數據進行統計分析,用最小顯著差數(LSD)法進行差異顯著性檢驗。
2 結果與分析
2.1 病原菌及其致病性
從閩楠患病葉片品中共分離得215個真菌菌株,經形態學初步檢測合并菌株后確定為15種。分離后確定的菌株按回接試驗結果顯示,菌株CB-2和CB-3均可產生葉斑病癥狀(圖1B)。其中,菌株CB-2和CB-3在30片病葉樣品中均有出現,其分離率分別為78.0%和86.0%。觀察閩楠葉斑病癥狀發現,病害初期在侵染點周邊呈黑點狀,后期逐漸擴大形成黑褐色不規則病斑,病斑邊上常產生密集的黑色小點,潮濕環境下發病部位常密布病原菌絲,與野外自然發病癥狀一致(圖1A)。對離體回接葉片進行病原菌的再分離,經形態學對比,同樣與菌株CB-2和CB-3一致。經柯赫氏法驗證,可確定菌株CB-2和CB-3為閩楠葉斑病致病菌。

注:A為野外發病病樣,B為離體回接致病癥狀。
2.2 病原菌的鑒定
2.2.1 形態學 菌株CB-2在PDA平板上的菌落生長呈圓形(圖2-A),初期為白色,培養5~6 d后變為鼠灰色,菌絲蓬松,有黑色小點,背面為黃綠色,近圓形,氣生菌絲濃密(圖2-B)。菌落生長速率12.4 mm/d。其分生孢子為無色,單胞,呈圓柱形,兩端鈍圓,大小為(15.0~25.0)μm×(4.0~7.5)μm(圖2C~D)。根據《植物病原真菌學》的方法初步鑒定菌株CB-2為膠孢炭疽菌(Colletotrichumgloesporioides)。菌株CB-3在PDA平板上菌落為白色絮狀,氣生菌絲濃密,背部淡黃色,具輪紋,生長較快。20 d 后菌絲表面形成黑色墨汁狀黏液,排列松散不規則,即為分生孢子堆,分生孢子梗短,圓柱形,無色。成熟分生孢子紡錘形,直或略彎,具4隔膜5細胞,大小(18.4~22.5)μm×(5.0~7.5)μm,初步鑒定菌株CB-3為小孢擬盤多毛孢(Pestalotiopsismicrospora)[9]。
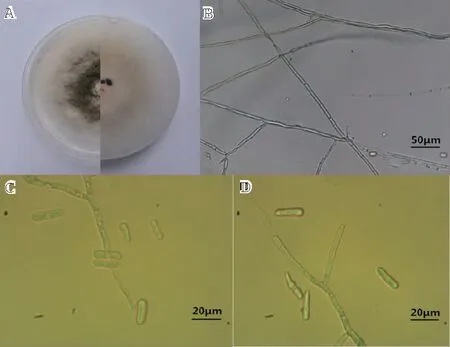
注:A為菌株CB-2的菌落,B~D為菌株CB-2的孢子及菌絲。
2.2.2 rDNA-ITS序列分析 以菌株CB-2總DNA為模板,ITS1/ITS4為引物擴增得到長度542 bp的序列,GenBank接收號為MK728960。該菌株序列經NCBI在線BLAST比,與GenBank號為MH700457.1、MF379341.1的膠孢炭疽菌(Colletotrichumgloeosporioides)的堿基序列覆蓋度達100%,序列相似度高達99%。用MEGA7.0 NJ法構建系統發育樹后發現其形成明顯的分枝,其中菌株CB-2與不同菌株來源的膠孢炭疽菌聚類為同一分枝(圖3),遺傳距離最近,結合形態學特征確定菌株CB-2為膠孢炭疽菌(Colletotrichumgloeosporioides)。
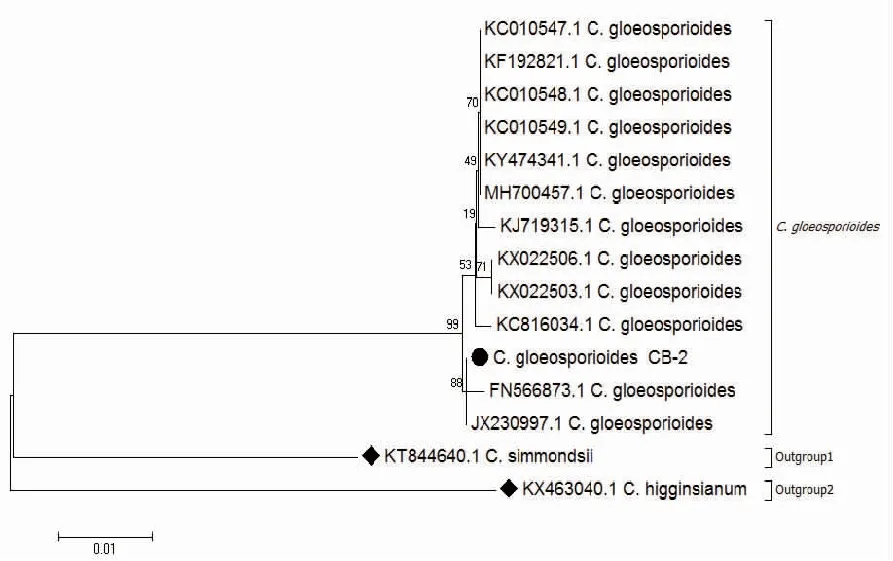
注:●為菌株CB-2,◆為外源菌株。
2.3 不同因素對膠孢炭疽菌菌絲體生長的影響
2.3.1 培養基 從圖4可知,PSA培養基最利于膠孢炭疽菌菌落的生長,培養5 d后的菌落直徑達9 cm,且菌絲最為茂密。在燕麥培養基上菌落生長速度僅次于PSA培養基,但在燕麥培養基上菌絲生長太過稀薄。在PDA+楠木葉煎汁瓊脂培養基上菌落直徑位于燕麥培養基之后。在察氏、SNA、PDA+維生素B6培養基中菌落生長較慢,在PDA、虎紅培養基上菌落直徑顯著降低,在對照組WA培養基上基本不生長。PSA培養基菌落直徑顯著大于其余培養基,WA培養基菌落直徑顯著小于其余培養基,其余基培養基間菌落直徑差異顯著或不顯著。
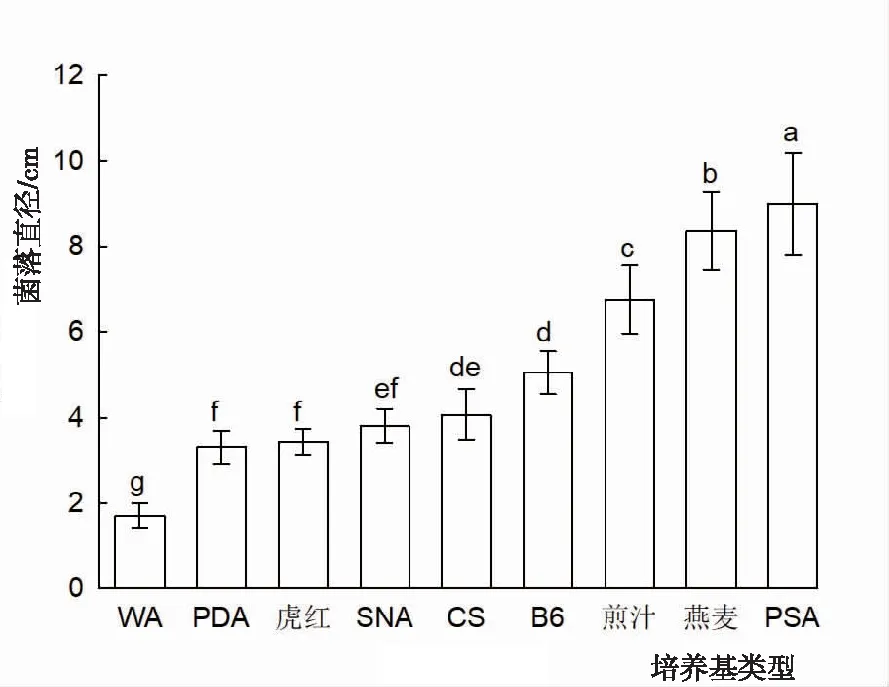
圖4 不同培養基閩楠葉斑病病原菌菌絲的生長狀況
2.3.2 溫度、pH與光照 從圖5看出,不同溫度、pH和光照下閩楠葉斑病病原菌菌落的生長狀況。溫度:病菌菌落在15~35℃均可生長,以25℃時菌絲的生長速度最快,各溫度下菌絲的生長速度依次為25℃>30℃>20℃>15℃>35℃。其中,25℃時菌絲的生長速度顯著大于除30℃外的其余溫度處理,35℃菌絲的生長速度顯著小于其余溫度處理。pH:為9.0時菌落直徑最大,各pH下菌落的生長速度依次為pH 9.0>pH 7.0>pH 8.0>pH 5.0>pH 6.0,pH 9.0與pH 7.0菌落直徑顯著大于其余pH處理,但二者間差異不顯著,其余pH處理間差異不顯著,表明該菌株不適宜在過酸或過堿的環境下生長。光照:全光照條件下菌落生長最好,菌落直徑為9 cm;光暗交替次之,菌落直徑為4.6 cm;全黑暗條件最差,菌落直徑為3.9 cm;全光照顯著大于光暗交替和全黑暗,光暗交替和全黑暗間差異不顯著。此外,在全黑暗情況下,菌絲生長擴散性不強,生長比較密集,趨于塊狀生長。
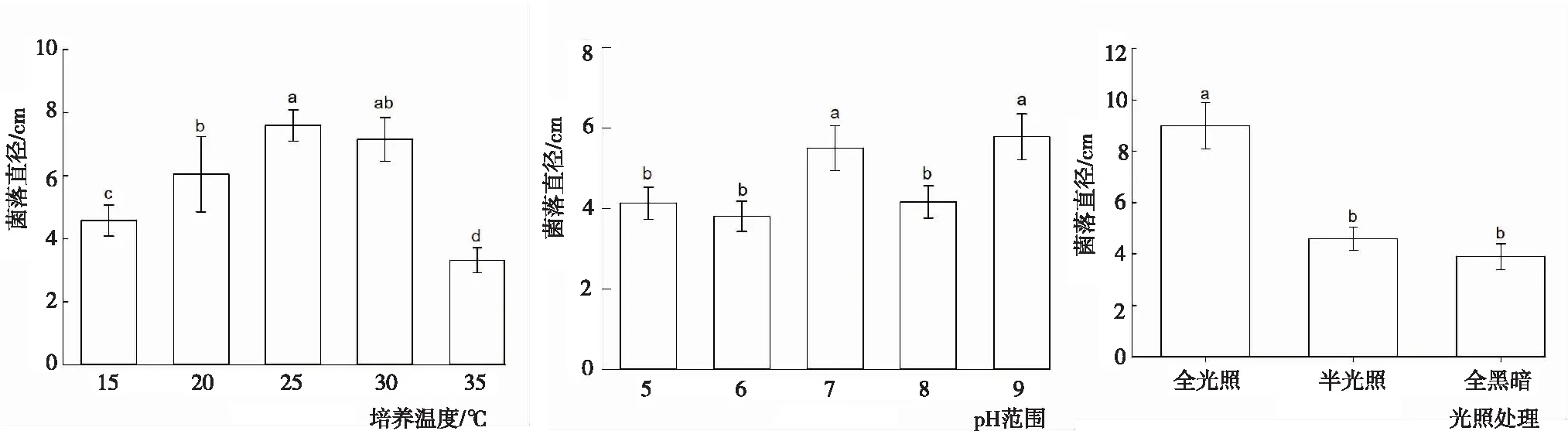
圖5 不同溫度、pH和光照下閩楠葉斑病病原菌菌絲的生長狀況
2.3.3 碳源與氮源 從圖6可知,不同碳源和氮源下閩楠葉斑病病原菌菌落生長的變化。碳源:各處理菌株均可生長,但菌落直徑存在差異。菌落直徑以麥芽糖最大,為4.90 cm;對照最小,為1.6 cm;各處理依次為麥芽糖>葡萄糖>蔗糖>果糖>可溶性淀粉>乳糖>對照。其中,麥芽糖顯著大于除葡萄糖和蔗糖外的其余處理,對照顯著小于其余處理,其余處理間差異顯著或不顯著。氮源:各處理菌株均可生長,但菌落直徑存在差異。菌落直徑以酵母浸粉最大,為4.3 cm;尿素最小,為0.5 cm;各處理依次為酵母浸粉>硝酸銨>蛋白胨>牛肉膏>對照>尿素。酵母浸粉顯著大于其余處理,尿素顯著小于除對照外的其余處理,其余處理間差異顯著或不顯著。
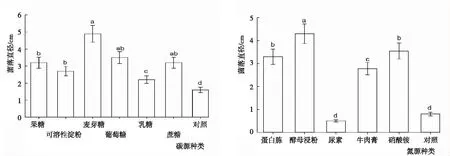
圖6 不同碳源與氮源下閩楠葉斑病病原菌菌絲的生長狀況
2.3.4 膠孢炭疽菌菌絲致死溫度 不同溫度水浴處理結果表明,菌絲塊在35~45℃水浴處理10 min后,置于PDA平板上培養,均可正常長出菌絲,50℃、55℃和60℃處理10 min,培養10 d后仍未觀察到菌絲生長。可見,膠孢炭疽菌菌絲的致死溫度為50℃下10 min。
3 結論與討論
閩楠是我國特有樹種,該樹種的生理特性、苗木繁育、群落生態、遺傳多樣性和野外資源等已有研究報道[11-14],但對閩楠病害的研究報道極少。陳全助等[9]研究結果表明,引起閩楠葉斑病的病原菌為小孢擬盤多毛孢(Pestalotiopsismicrospora)。研究結果表明,除P.microspora可引起閩楠葉斑病外,膠孢炭疽菌(C.gloeosporioides)也可引起閩楠葉斑病的發生。C.gloeosporioides寄主范圍廣泛,可侵染山楂、蘋果、桂花、決明子和柿樹等多種樹種,引起相關病害[15-19],C.gloeosporioides侵染閩楠葉片引起葉斑病為國內首次報道。
研究結果表明,C.gloeosporioidesCB-2的生物學特性與張翊等對C.gloeosporioides的生物學特性研究結果不完全一致[20],這可能與菌株來源不同或菌株所處環境差異有關。C.gloeosporioidesCB-2在25℃時更利于菌株的生長,當溫度繼續升高或下降菌株生長均呈下降趨勢。推測閩楠葉斑病的發生多在25℃左右,因此,在這個溫度對應的季節之前應做好該病害的防控工作。全光照更利于菌株的生長,結合閩楠自身生長特性為陰性樹種,因此,要盡量避免閩楠幼苗栽植在光照較強的地域,或在幼苗栽植過程中做好遮陰處理,即可滿足閩楠自身生長特性所需,也可減少閩楠葉斑病的發生。另外,結合C.gloeosporioidesCB-2的其他生物學特性及閩楠生長特性,在其幼苗栽植及管護過程中要注意合理的播種密度,保持苗木通風,適度噴灌,控制幼苗期光照時長,做好遮陰處理,在溫度達25℃前做好病害的防控工作。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
