凍融對肌原纖維蛋白溶出豬肉糜體系蛋白質變性及品質的影響
2021-10-22 00:21:58吳興閣曾茂茂何志勇王召君秦昉張志剛陳潔
食品與發酵工業 2021年19期
吳興閣,曾茂茂,何志勇,王召君,秦昉,張志剛,陳潔*
1(食品科學與技術國家重點實驗室(江南大學),江蘇 無錫,214122) 2(廈門銀祥集團有限公司 肉食品安全生產技術國家重點實驗室,福建 廈門,361100)
中國是肉類生產及消費大國,隨著經濟的發展,肉制品的生產與消費呈持續增長態勢。凍藏是肉類食品原料及產品最常用和有效的保藏方式,但在實際生產、貯藏、運輸和銷售的過程中,由于冷鏈系統的不完善甚至缺失,肉及肉制品反復經歷凍融,導致品質劣變。肉糜作為肉丸、肉餡、肉腸等眾多肉制品的主要原料,占比在50%以上,年需求量約530萬t,且逐年增加,但肉糜制品在凍藏過程的反復凍融易導致其品質下降[1-4]。
肉類在凍藏及反復凍融過程中可發生大量的理化反應,如脂肪氧化生成丙二醛等小分子醛類物質,蛋白發生氧化導致羰基增加和巰基減少等[5-7],從而引起肉制品風味、色澤和質構等品質的劣化[8-9]。余小領等[10]研究常規冷凍凍藏對豬肉保水性影響時發現,隨著凍藏時間的增加,豬肉保水性下降;LIU等[11]研究氧化對豬背最長肌性質的影響時發現,隨著氧化的加深,肌肉的持水性下降;MIGNINO等[12]發現使用冷凍肉制作的肉制品,其凝膠性能降低。這些性質的下降都與肌原纖維蛋白有關,肌原纖維蛋白質不僅是肉制品中重要的營養組分,還決定著肉制品的理化性質。肌原纖維蛋白約占豬肉總蛋白質的50%~60%,是一種重要的鹽溶性蛋白質。凍融過程中的氧化和冷凍變性會使肌原纖維蛋白結構發生變化[13]。ZHANG等[14]研究發現,隨著凍融次數的增加,未加鹽豬肉糜的肌原纖維蛋白Ca2+-ATP酶活性下降,導致其持水性下降,表面疏水性增加;ARSLAN[15]研究發現,凍藏使肌肉蛋白乳化能力和乳化穩定性下降;XIA等[16]研究凍融后的豬背最長肌發現,肌原纖維蛋白變性和聚集使其凝膠性質下降。肌紅蛋白賦予了肉制品誘人的色澤,但隨著氧化的加深,肉色會呈現令消費者難以接受的深褐色。同時,脂肪的氧化也會對肉制品的色澤產生一定的影響[17]。肌原纖維蛋白在NaCl 濃度>0.5 mol/L時可以充分溶解[18-19],工業上制作一些肉糜時會加入鹽和冰水并且經過斬拌,此時肌原纖維蛋白已經充分溶出,由此形成的體系即為肌原纖維蛋白溶出的肉糜體系。然而,目前研究大多針對原料肉或加鹽未經斬拌的肉餡等,此時的肌原纖維蛋白仍被包裹在肌束膜中,并沒有溶出,很少有研究報道肌原纖維蛋白在加工過程中已經溶出的肉糜體系。
本文通過考察不同凍融次數對肉糜肌原纖維蛋白變性的影響,以及肌紅蛋白的含量與不同價態肌紅蛋白的比例變化情況,分析不同凍融次數肉餡蒸煮后的感官品質,旨在研究蛋白質變性對肉糜體系質構等感官品質的影響,為工業生產此類冷凍肉餡原料的品質保持和提升提供依據。
1 材料與方法
1.1 材料與儀器
豬背最長肌肉(壹號土豬,11個月,公豬,屠宰后24 h內),無錫華潤萬家超市;L-亮氨酸標品,美國SIGMA公司;其余試劑均為分析純。
CK22絞肉機,意大利La Minerva公司;CM-14斬拌機,西班牙MAINCA公司;VF620真空灌腸機,德國漢德曼公司;AL-104精密電子天平,梅特勒-托利多有限公司;Sigma3K 15離心機,德國SIGMA公司;IKA T18 digital均漿機,德國IKA公司;Spectra Max 190全波長酶標儀,美國Molecular Devices公司;MesoMR23-060V-I低場核磁共振儀,中國紐邁公司;UltraScan Pro1166高精度分光測色儀,美國Hunterlab公司;Q2000 差示掃描量熱儀,美國TA儀器公司;SCC61型萬能蒸烤箱,德國RATIONAL公司;TA.XT plus質構儀,英國Stable Micro Systems公司。
1.2 實驗方法
1.2.1 肉糜樣品的制備
取新鮮豬背最長肌,去除可見肥肉和結締組織,加入鹽、水等,經過斬拌制作成肌原纖維蛋白充分溶出的肉餡[18-19]。將制作好的肉餡用灌腸機灌入直徑6 cm的PE膜腸衣中,每根500 g。處理好的預調理肉餡放入-18 ℃冰箱冷凍36 h,4 ℃解凍12 h,此為1次凍融。以此類推,將肉餡凍融0~7次。肉餡的配方及工藝由廈門銀祥集團有限公司肉食品安全生產技術國家重點實驗室提供。
基本配方:以豬背最長肌肉1 000 g計,純堿6.5 g,小蘇打10 g,食鹽30 g,味精10 g,生抽20 g,雞精10 g,冰水550 g。
整個工藝控制肉的溫度在4~6 ℃。工藝流程:
原料肉處理(豬背最長肌剔除可見脂肪和結締組織,切成2 cm×2 cm×2 cm方塊)→絞肉機絞碎(8 mm孔板)→加入鹽和20%冰水中速1 500 r/min斬拌20 s→加入純堿和小蘇打,20%冰水中速1 500 r/min斬拌20 s→加入剩下調味料和冰水高速3 500 r/min斬拌2 min→灌腸→冷藏
1.2.2 肉糜的解凍與蒸煮損失
肉糜在解凍前和解凍后質量分別為m0、m1,按公式(1)計算解凍損失率:
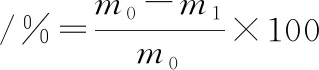
(1)
將上述經過不同凍融次數的肉糜用不銹鋼圓柱型模具(d=2.72 cm,h=0.84 cm)制成圓柱體后稱其質量(m3),放入萬能蒸烤箱中,采用蒸煮模式,85 ℃蒸煮20 min后用濾紙擦干表面的水分后稱其質量為m2)。蒸煮損失率按公式(2)計算[10]:
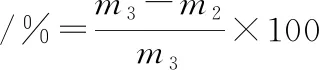
(2)
1.2.3 肉糜蛋白溶解度的測定
參照貢漢坤等[20]的方法,測量蛋白濃度。取0.5 g上述經過不同凍融次數的肉糜樣品,加入20 mL 0.05 mol/L的磷酸鹽緩沖液(含0.5 mol/L NaCl,pH 7.2),勻漿60 s后離心(10 000×g,10 min)。取上清液測量蛋白濃度,使用雙縮脲法。
1.2.4 肉糜低場核磁共振分析
稱取15 g肉糜樣品用保鮮膜包住放入核磁管中,采用Carr-Purcell-Meiboom-Gill(CPMG)脈沖程序進行掃描,設置參數為采樣間隔時間TW=4 500 ms,回波個數Echo Count=3 000,累加次數NS=8。
1.2.5 肉糜肌原纖維蛋白的提取
肌原纖維蛋白的提取參照PARK等[21]的方法并稍作修改,將經過不同凍融次數的豬肉糜樣品置于5倍體積的僵直液[0.1 mol/L NaCl,2 mmol/L MgCl2,1 mmol/L乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA),10 mmol/L K2HPO4,pH 7.0]中均質1 min,然后4 ℃ 2 000×g離心10 min,棄去上清液,此過程重復3次。向離心后的沉淀加入4倍體積0.1 mol/L NaCl均質30 s,離心(4 ℃,2 000×g,10 min),重復3次。最后一次勻漿液調節pH 6.25,用4層紗布過濾后離心(4 ℃,2 000×g,10 min),棄去上清液,所得沉淀即是肌原纖維蛋白。以上所有操作均在4 ℃下完成。提取后的蛋白膏置于冰盒中保存,48 h內用完,蛋白的濃度采用雙縮脲法測定。
1.2.6 肌原纖維蛋白羰基值的測定
羰基值的測定參考LEVINE等[22]的方法并稍有改變,用2,4-二硝基苯肼(2,4-dinitrophenyl hydrazine,DNPH)衍生后比色測定。取上述所提取的蛋白,用緩沖液稀釋至20 mg/mL,取200 μL與2 mL 10 mmol/L DNPH混勻,室溫下避光反應1 h,每隔10 min渦旋振蕩1次。加入2 mL質量分數20% TCA中止反應,10 000×g離心5 min,沉淀用4 mLV(乙醇)∶V(乙酸乙酯)=1∶1溶液洗滌,重復3次后沉淀用N2吹干,加入1.5 mL 6 mol/mL的鹽酸胍(溶解在20 mmol/L KH2PO4),37 ℃反應30 min后離心(10 000×g,10 min)。上清液在370和280 nm處比色。羰基的含量(nmol/mg)按公式(3)計算:
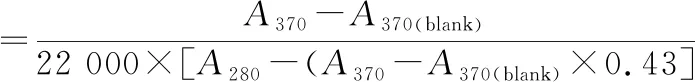
(3)
1.2.7 肌原纖維蛋白自由氨基值的測定
自由氨基值采用李春強[23]的方法并稍加改動,將上述肌原纖維蛋白溶液用磷酸鹽緩沖液稀釋至4 mg/mL,取200 μL溶解在0.2 mol/L 的磷酸鹽緩沖液[含質量分數1%十二烷基硫酸鈉(sodium dodecyl sulfate,SDS),pH 8.2]中,加入1 mL質量分數0.01%三硝基苯磺酸(3-nitrobenzenesulfonic acid,TNBS)于37 ℃避光60 min。用2.0 mL 0.1 mol/L Na2SO3終止反應,室溫放置15 min冷卻后在420 nm處測吸光值。自由氨基的含量根據L-亮氨酸做標準曲線確定。
1.2.8 肌原纖維蛋白巰基和二硫鍵的測定
巰基值測定采用LIU等[24]的方法,用pH 6.25,25 mmol/L的磷酸鹽緩沖液,其中含0.6 mol/L的NaCl,將肌原纖維蛋白樣品稀釋到約2 mg/mL,取0.5 mL上述稀釋后的肌原纖維蛋白樣品加入棕色的離心管中,空白樣品用緩沖液代替。加入2 mL pH 7.4 0.1 mol/L磷酸鹽緩沖液,其中含有8 mol/L尿素,3% SDS,混合均勻。加入0.5 mL 10 mmol/L的5,5′-二硫代雙(2-硝基苯甲酸) (5,5-dithiobis-2-nitrobenzoic acid,DTNB),室溫下反應15 min,在412 nm處比色,摩爾消光系數為13 600 L/(mol·cm)。巰基的含量(nmol/mg)按照公式(4)計算:
(4)
二硫鍵的測定采用THANNHAUSER等[25]的方法,用pH 6.0,25 mmol/L的磷酸鹽緩沖液(含0.6 mol/L的NaCl)將肌原纖維蛋白稀釋到約5 mg/mL,取0.1 mL上述稀釋后的肌原纖維蛋白溶液,加入1.5 mL現配的disodium 2-nitro-5-thiosulfobenzoate(NTSB)溶液,室溫下反應25 min后于412 nm處比色,空白組加入0.1 mL的磷酸鹽緩沖液。二硫鍵含量(nmol/g)按照公式(5)計算:
(5)
1.2.9 DSC測定
肌原纖維的熱變性溫度用差示掃描量熱儀(differential scanning calorimetry,DSC)測量,在專用的坩堝中稱取肉糜約15 mg,以空坩堝作為參照進行測量。在25 ℃下平衡1 min,升溫速率5 ℃/min,從25 ℃掃描到95 ℃。使用Universal Analysis 2000軟件計算樣品的變性溫度和焓變值。
1.2.10 肌紅蛋白含量的變化
參考陳騁[26]的方法,解凍后取2.0 g樣品加入20 mL的緩沖液,10 000 r/min勻漿30 s,然后4 ℃、5 000×g離心15 min,上清液用濾紙過濾后在576 nm處測量吸光值,摩爾消光系數為12.8 L/(mmol·cm)。
1.2.11 氧合肌紅蛋白、高鐵肌紅蛋白含量測定
參考KIM等[27]的方法。稱取解凍后樣品5.0 g,加入25 mL 磷酸緩沖液(40 mmol/L,pH 6.8,4 ℃),1 000 r/min 勻漿25 s 后冰浴靜置1 h,然后離心(3 000×g,25 min,4 ℃),上清液經濾紙過濾后于450~650 nm進行掃描,分析肌紅蛋白吸收光譜的變化,同時記錄525、545、565、572 nm波長下的吸光值,按照 KRZYWICKI[28]的方法計算氧合肌紅蛋白(oxymyoglobin,OMb)和高鐵肌紅蛋白(metmyoglobin,MMb)的相對含量,分別按公式(6)(7)計算:
OMb/%=(0.882R1-1.267R2+0.809R3-0.361)×100
(6)
MMb/%=(-2.514R1+0.777R2+0.800R3+1.098)×100
(7)
式中:R1、R2、R3分別為 572、565、545與 525 nm 吸光值的比值。
1.2.12 顏色的測定
使用高精度分光測色儀(光源D65,視角10°)觀察凍融不同次數的肉糜的顏色(亮度值L*,紅度值a*和黃度值b*),使用反射模式,測試前使用標準的白板和黑板對儀器進行校準。對于每塊樣品隨機選取5個位置進行測量,然后取平均值。
1.2.13 肉糜全質構測定
肉糜制品85 ℃蒸煮20 min,然后用模具切成直徑為2.72 cm,高為0.84 cm的圓柱體。使用質構儀應用質構剖面分析法對其硬度、彈性、內聚性和咀嚼性進行分析。使用P/36R型探頭,設置測前速度為3.0 mm/s,測試速度為5.0 mm/s,測試后速度為5.0 mm/s,壓縮比為60%,觸發類型自動,觸發力10 g。
1.2.14 肉餡的感官評價
取30 g凍融不同次數的肉餡樣品,解凍后放入烤箱中,100 ℃蒸20 min。將樣品打亂隨機編號,放入相同的容器中進行感官評價。感官評價小組由15名食品專業的研究生組成,分別從肉餡的色澤、氣味、口感、硬度、彈性和內部結構6個方面進行評價,感官最高得分為10分,最低為1分。感官評價標準如表1所示。

表1 肉餡感官評價標準Table 1 Criteria for sensory evaluation of minced meat
1.3 數據處理
除感官評價外,所有實驗均重復3次,結果用平均值±標準偏差表示,感官評價由15名感官評價員打分的平均值表示。平均值和方差使用Microsoft Excel計算,顯著性分析使用Statistix 9.0中的Linear Models進行分析,采用LSD全配對多重比較方法進行差異顯著性分析(P<0.05,差異顯著)。使用IBM SPSS Statistics 22進行相關性分析(*表示P<0.05水平上相關,**表示P<0.01水平上相關),SIMCA+13.0軟件用于主成分分析,使用Origin 8.5軟件作圖。
2 結果與分析
2.1 凍融對豬肉糜水分和蛋白溶解度的影響
如表2所示,肉糜解凍后并未出現汁液流失和質量減輕,低場核磁共振分析顯示,解凍后的豬肉糜中的水分有2種分布形態,分別是T2b=1~10 ms,反映與大分子緊密結合的水,T21=10~100 ms,反映被束縛在蛋白結構內部的水。隨著凍融次數的增加,結合水的出峰時間顯著減小,比例顯著下降(P<0.05),不易移動水的出峰時間和比例均未發生顯著的變化(P>0.05),并未發現自由水,可能是因為肉糜斬拌的過程中肌原纖維蛋白溶出,形成蛋白網狀結構,肉糜中的水分被束縛在其間隙之中,這與解凍過程中未出現解凍損失的結果一致。豬肉糜的蒸煮損失率隨著凍融次數的增加顯著上升(P<0.05),未經過凍融的豬肉糜的蒸煮損失最小為6.67%,而經過1次凍融后蒸煮損失上升至7.19%。經過7次凍融后,豬肉糜的蒸煮損失比未凍融時上升了30.28%。蒸煮損失和豬肉糜加熱過程中形成的蛋白凝膠有著密切的關系,凝膠結構越緊密,保留的水分越多。未經凍融豬肉糜中的肌原纖維蛋白保持天然的構象,在加熱的過程中,可以更好地形成蛋白間的聚集物,從而形成均一致密的凝膠網絡結構,更好地保留豬肉糜中的水分。經凍融后的豬肉糜,由于蛋白存在冷凍和氧化變性,蛋白質結構打開,或形成不溶的蛋白質聚集體,導致凝膠結構無序粗糙,持水能力下降。同時,凍融過程中脂肪氧化產生丙二醛,與肌原纖維蛋白相互作用形成復合物,使蛋白變性,降低蛋白的溶解性[29-30],從而使加熱后蛋白持水能力下降。
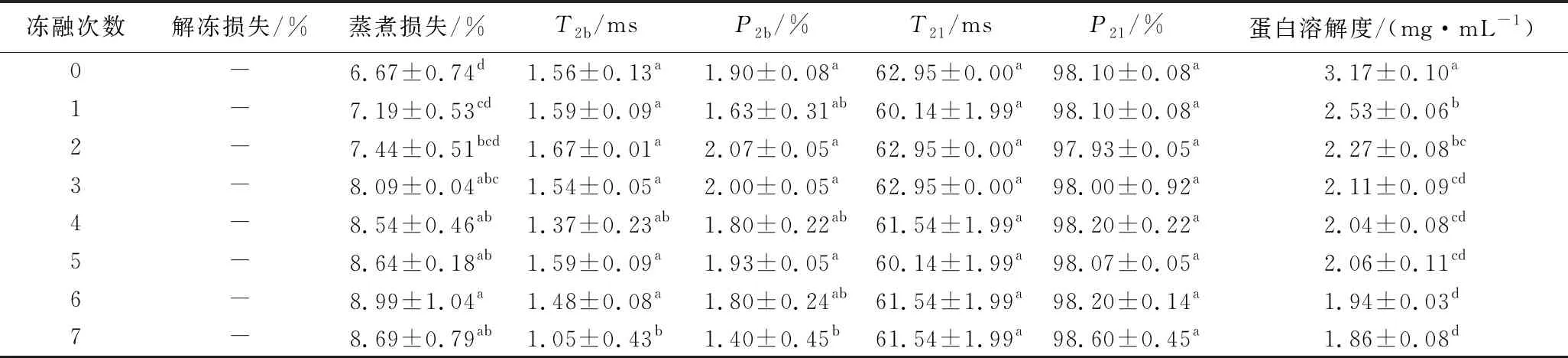
表2 不同凍融次數豬肉糜水分和蛋白溶解度的變化Table 2 Changes of moisture and protein solubility of minced pork with different freeze-thaw cycles
如表2所示,與未凍融組相比,在凍融1次之后肉糜蛋白的溶解度下降了20.19%(P<0.05),隨著凍融次數的增加,肉糜蛋白的溶解度仍然持續地降低,在凍融7次的時候降至最低(1.86 mg/mL)。這可能是因為在第1次凍融的時候,由于肌原纖維蛋白溶出,蛋白質發生了冷凍變性,分子結構發生改變,使包埋在蛋白質分子內部的疏水基團暴露,蛋白質的溶解度發生顯著地下降。隨著凍融次數地增加,蛋白質分子進一步發生冷凍變性,同時發生氧化變性,蛋白質分子間氫鍵和表面電荷發生改變,氧化生成聚集物,溶解度進一步下降。
2.2 凍融對肌原纖維蛋白羰基值的影響
羰基是蛋白質氧化的標志性產物。如圖1所示,隨著凍融次數的增加,羰基值呈現逐漸增加的趨勢,從未凍融的羰基含量2.51 nmol/mg 蛋白上升至5.14 nmol/mg 蛋白,說明凍融的過程中氧化持續存在。未凍藏樣品的羰基值略高于其他研究中未經斬拌的肉制品[21-23],這可能是因為在斬拌的過程中大量氧氣混入肉糜體系,加速了肌原纖維蛋白的氧化。凍融1次和2次時,羰基值緩慢地上升,但不顯著(P>0.05);凍融3~5次時,肌原纖維蛋白羰基值顯著地上升(P<0.05);凍融5~7次肌原纖維蛋白羰基值變化不顯著(P>0.05)。研究表明,新生成的羰基會與蛋白質中的親核物質,如ε-氨基,發生反應,使得后期羰基含量變化不顯著[31]。
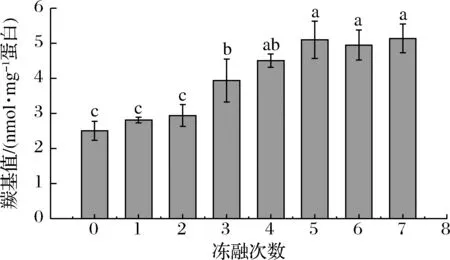
圖1 不同凍融次數豬肉糜蛋白羰基含量的變化Fig.1 Changes in carbonyl content of minced pork protein with different freeze-thaw cycles注:組間不同字母表示差異顯著(P<0.05)(下同)
2.3 凍融對肌原纖維蛋白自由氨基值的影響
圖2反映了不同凍融次數肉糜肌原纖維蛋白自由氨基含量的變化。未凍融前自由氨基的含量最低為42.82 nmol/mg 蛋白,在凍融1次和2次的時候顯著地升高(P<0.05),最高升至55.29 nmol/mg 蛋白,在凍融5~7次的時候,自由氨基的含量變化不顯著(P>0.05)。這說明凍融前期肌原纖維蛋白結構展開,更多的自由氨基暴露。隨著凍融次數的增加,氧化的加深使得蛋白聚集自由氨基含量減少,同時生成的羰基也可以和自由氨基反應,使其減少[27]。
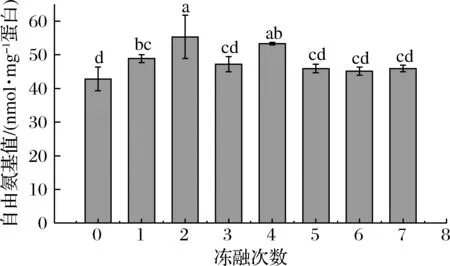
圖2 不同凍融次數豬肉餡自由氨基含量的變化Fig.2 Changes of free amino content of minced pork protein with different freeze-thaw cycles
2.4 凍融對肌原纖維蛋白巰基和二硫鍵的影響
肌原纖維蛋白中含有大量的巰基,如圖3所示,隨著凍融次數的增加巰基含量顯著減小(P<0.05)。未經過凍融的肌原纖維蛋白巰基含量為104.85 μmol/g 蛋白,第7次凍融后含量下降至62.55 μmol/mg 蛋白,下降了40.34%。二硫鍵的含量從1.42 μmol/g 上升至9.03 μmol/g。說明凍融的過程中肉糜肌原纖維蛋白巰基被氧化生成分子內和分子間的二硫鍵,這也是肌原纖維蛋白氧化聚集的主要途徑[32]。

圖3 不同凍融次數豬肉糜蛋白巰基和二硫鍵含量的變化Fig.3 Changes of sulfhydryl and disulfide bond content of minced pork protein with different freeze-thaw cycles
XU等[33]研究鰱魚肌動球蛋白凝膠形成的過程,猜測二硫鍵的增加可能是因為巰基基團的氧化,但本文中巰基的下降速率遠高于二硫鍵的生成速率,說明肌原纖維蛋白氧化的過程中除了生成二硫鍵以外,還生成了其他的含硫衍生物。由于肌原纖維蛋白加熱過程中巰基可生成二硫鍵,促使肌原纖維蛋白凝膠的形成[34]。凍融的過程中由于氧化聚集,巰基含量減少,二硫鍵含量增加,使得肌原纖維蛋白凝膠強度變弱,肉糜制品的彈性、脆度下降,感官品質下降。
2.5 凍融對蛋白熱變性的影響
由表3可知,豬肉糜有2個吸熱峰,分別在59.02~59.48 ℃和71.37~72.00 ℃,隨著凍融次數的增加,肉糜吸熱峰的位置并沒有發生顯著的偏移(P>0.05)。豬肉糜的總焓變隨著凍融次數的增加呈現逐漸下降的趨勢(P<0.05),未凍融組的總焓變為0.38 J/g,經過7次凍融之后的總焓變為0.25 J/g,下降了34.21%。ZHANG等[35]發現新鮮的豬肉樣品有3個明顯的吸熱峰,分別在53、64和77 ℃,且總焓變隨著冷凍解凍減小。這3個吸熱峰分別對應肌球蛋白重鏈、肌球蛋白和肌動蛋白的變性溫度,而本文中測得的豬肉糜樣品只有2個吸熱峰,且吸熱峰的溫度降低。這可能是因為斬拌肉糜的過程中NaCl的加入使得豬肉中肌原纖維蛋白溶出,熱穩定性降低。
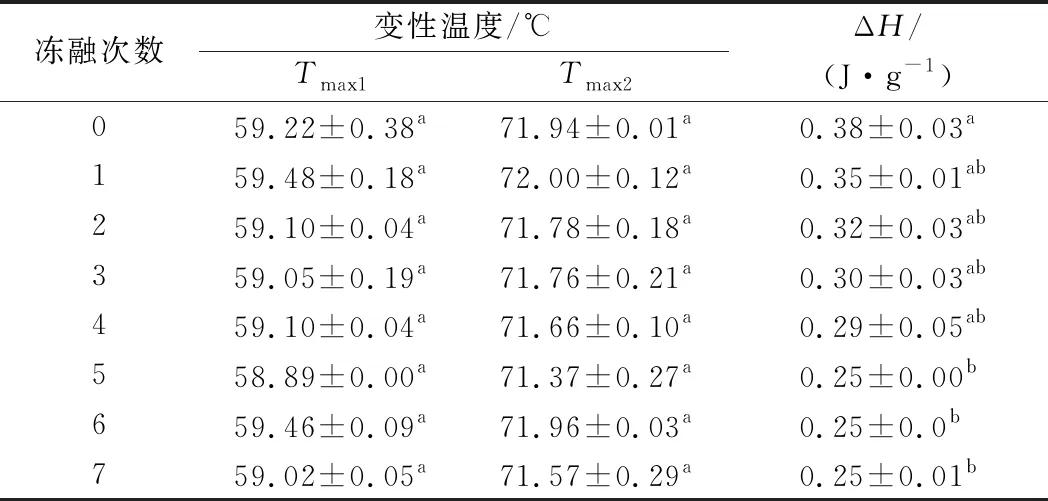
表3 不同凍融次數對豬肉糜蛋白變性溫度和總焓變的影響Table 3 Effect of denaturation temperature and total enthalpy change of minced pork proteins in different freeze-thaw cycles
因為鹽離子與肌原纖維蛋白發生相互作用,導致蛋白鏈伸展或者去折疊結構等空間結構發生變化,使蛋白質之間相互作用減弱,蛋白質的變性溫度減小[36]。由于Na+和Cl-通過水合作用形成水分子層,減弱蛋白質與水之間的相互作用,導致蛋白質親水性降低,從而使得蛋白質變性溫度減小。孟祥忍等[37]研究發現,隨著食鹽添加量的增加,雞肉糜變性溫度逐漸降低,這也與本文的研究結果一致。
2.6 凍融對肌紅蛋白含量變化的影響
血紅蛋白、肌紅蛋白和細胞色素賦予肌肉誘人的紅色,在動物被宰殺放血后,大量的血紅蛋白流失,此時肌紅蛋白是肉及肉制品呈現紅色的主要原因。圖4是凍融過程中肌紅蛋白含量的變化,未經過凍融的肉糜肌紅蛋白的含量最高為59.38 μmol/g,在1次凍融之后即顯著下降(P<0.05)至53.01 μmol/g,隨后逐漸降低,第7次凍融后含量下降至46.92 μmol/g。由此可知,第1次解凍后肉糜中的肌紅蛋白便出現了氧化,使其含量減少。隨著凍融次數的增加,肉糜的氧化加劇。脂肪氧化過程中產生的自由基也會攻擊肌紅蛋白血紅素輔基,生成Fe3+,同時自由基會影響高鐵肌紅蛋白還原酶活力,使生成的Fe3+不能及時還原,肉色劣化,肉糜的肌紅蛋白含量呈現逐漸降低的趨勢。

圖4 不同凍融次數豬肉糜肌紅蛋白含量的變化Fig.4 Changes of myoglobin content in minced pork protein with different freeze-thaw cycles
2.7 凍融對氧合肌紅蛋白、高鐵肌紅蛋白含量的影響
除了肌紅蛋白含量決定了肉制品的色澤之外,肌紅蛋白的價態也是影響顏色的另一個重要因素。圖5 顯示了氧合肌紅蛋白和高鐵肌紅蛋白隨凍融次數比例的變化,與空白組相比,第1次凍融之后肉糜的氧合肌紅蛋白比例顯著上升,高鐵肌紅蛋白比例顯著下降(P<0.05);凍融2、3次的時候變化不明顯;凍融4~7次時,氧合肌紅蛋白的比例顯著地下降,高鐵肌紅蛋白的比例顯著地上升(P<0.05)。新鮮肉中肌紅蛋白以暗紫色的脫氧肌紅蛋白形式存在,當接觸O2時,O2迅速和脫氧肌紅蛋白上的鐵離子配位鍵結合形成氧合肌紅蛋白,氧合肌紅蛋白的比例上升,肉制品呈現誘人的鮮紅色。但隨著凍融次數的增加,氧化加深,氧合肌紅蛋白和脫氧肌紅蛋白中的Fe2+被氧化成Fe3+,形成高鐵肌紅蛋白,肉制品顏色劣變成褐色。凍融初期高鐵肌紅蛋白還原酶可將生成的Fe3+還原,從而抑制了顏色的劣變,但隨著氧化的加深,還原系統被破壞,高鐵肌紅蛋白的比例上升,肉糜顏色加深劣變。
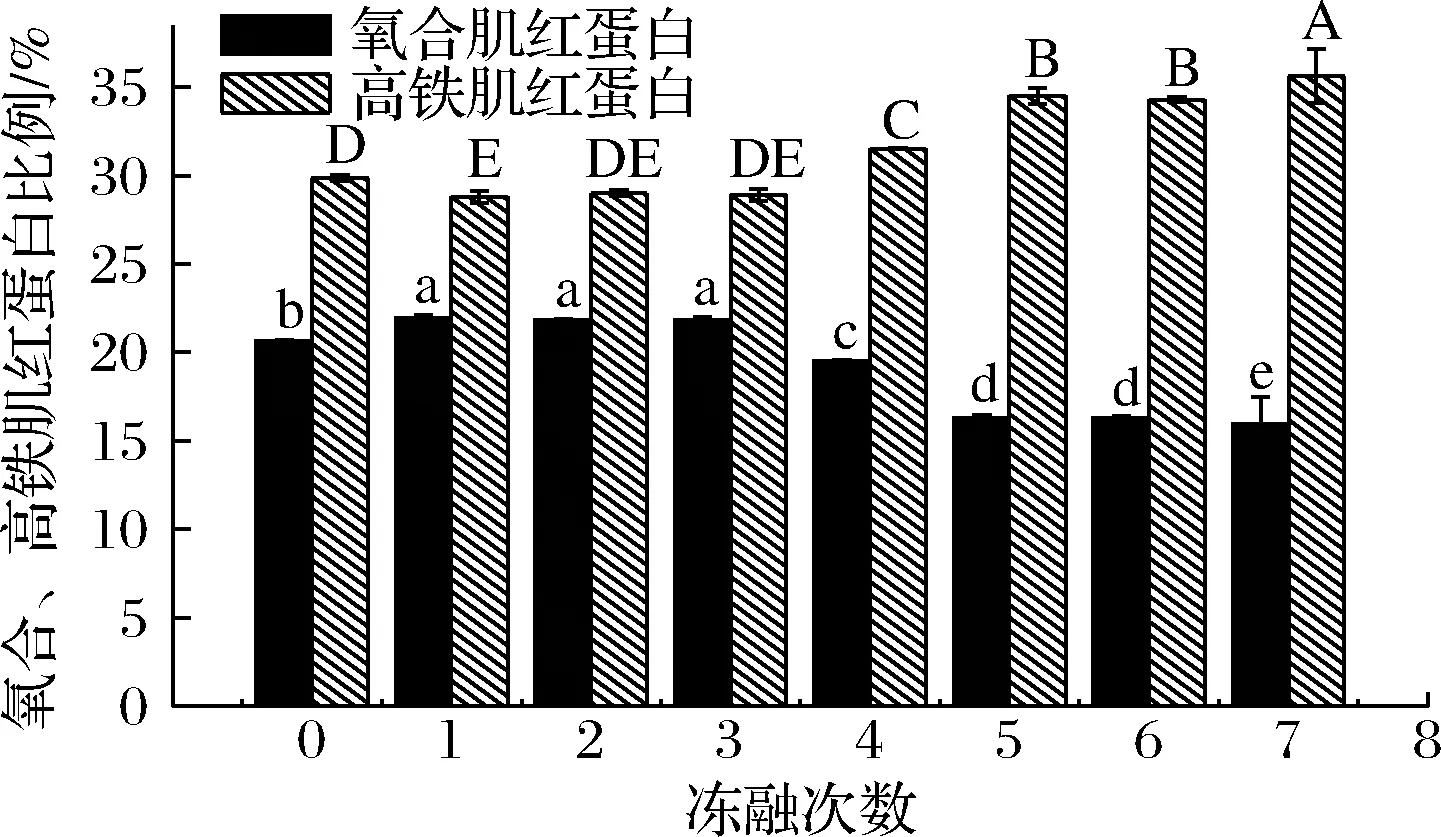
圖5 不同凍融次數豬肉糜氧合肌紅蛋白和高鐵肌紅蛋白比例的變化Fig.5 Changes of the ratio of OMb and MMb in minced pork with different freeze-thaw cycles
2.8 凍融對肉餡顏色的影響
表4顯示不同凍融次數肉糜L*、a*和b*值的變化情況。新鮮的肉糜(未經過凍融的)a*值最高,凍融1次后顯著下降(P<0.05),隨后變化不顯著。這是由于肉在斬拌過程中大量的O2混入,肌紅蛋白上的鐵離子與O2結合生成鮮紅的氧合肌紅蛋白,使得a*值升高。L*值和b*值主要和脂肪氧化有關[38],L*值、b*值變化均不顯著,可能是因為原料采用豬背最長肌肉,脂肪含量低。凍融1次后L*值有一定的上升,這可能是因為解凍之后部分水在肉糜的表面,使得肉糜的L*值升高。
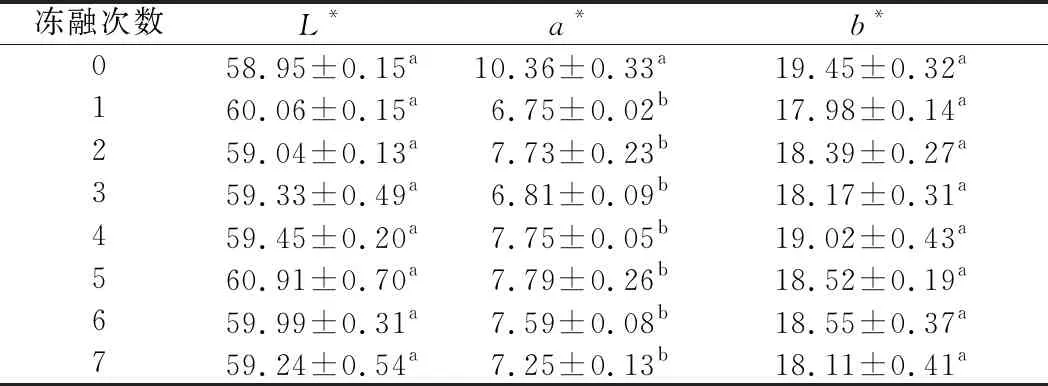
表4 不同凍融次數豬肉糜顏色的變化Table 4 Color changes of minced pork with different freeze-thaw cycles
2.9 凍融對肉餡質構的影響
如表5所示,隨著凍融次數的增加,豬肉糜的質構發生了顯著的變化。其中未經過凍融的豬肉糜樣品硬度為7 756 g,經7次凍融后,硬度降至最低(6 827 g)。彈性、內聚性和咀嚼性凍融7次后,分別下降了6.0%、7.4%和21.0%(P<0.05)。內聚性是咀嚼肉糜時,肉糜內部緊密相連抵抗受損的性質,它反映了肉糜內部結合程度的大小,凍融過程中肉糜內部凝膠結構由于受到冰晶的破環,或者蛋白質氧化聚集使得內部結構變得松散[39],內聚性降低。XIA等[16]研究指出,凍融循環降低肉糜的凝膠性質和乳化能力,凍融循環次數越多,下降越明顯。咀嚼性是硬度、內聚性和回復性的乘積,它綜合地反映了肉糜的感官品質。由表5可知,未經凍融的肉糜的咀嚼性最好,凍融1~4次后發生劣變,凍融5~7次后咀嚼性下降顯著(P<0.05),說明凍融5次后肉糜感官品質下降明顯,這也與感官評價結果相近。

表5 不同凍融次數豬肉糜質構的變化Table 5 Changes in texture of pork mince with different freeze-thaw cycles
2.10 凍融對肉餡感官品質的影響
由表6可知,未凍融的肉餡色澤感官評分最高,隨后感官評分降低,這說明在1次凍融后肉糜的顏色發生了肉眼可辨的變化,隨凍融次數的增加,雖然肉糜的顏色仍然在劣變,但肉眼幾乎分辨不出。新鮮肉糜與凍融后的肉糜在氣味上沒有顯著差異(P>0.05),而口感卻發生了變化,未經過凍融的口感得分最高為7.273分,而凍融7次后降為6.82分。同時質地方面,硬度、彈性和內部結構的感官得分均顯著降低,硬度從19.09 g下降到16.14 g(P<0.05),彈性從19.32 mm下降到16.14 mm(P<0.05),內部結構從15.46下降到13.09(P<0.05)。感官評價的結果與質構結果相近,質構結果也顯示隨著凍融次數的增加,肉糜在硬度、彈性和內聚性方面下降。
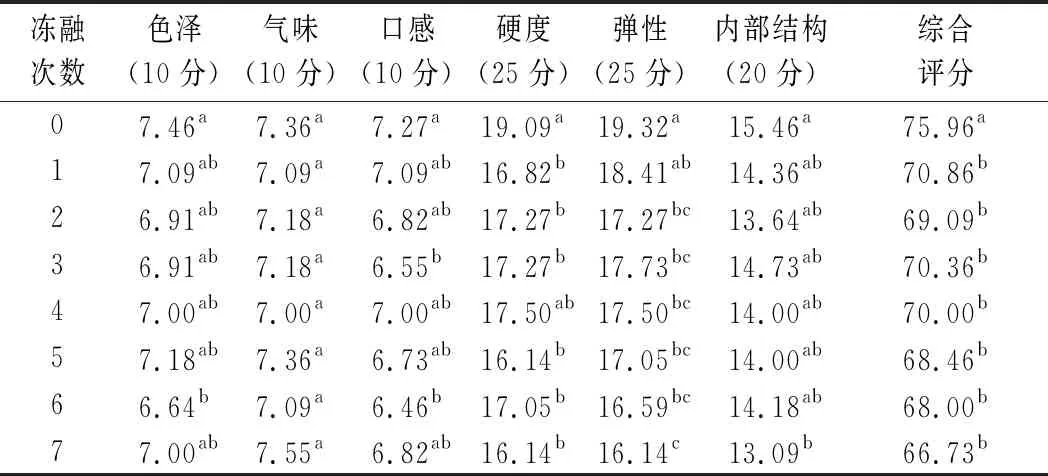
表6 不同凍融次數豬肉糜感官品質評價得分Table 6 Sensory quality evaluation scores of minced pork with different freeze-thaw cycles
肉糜的質地和加熱過程中肉糜形成的蛋白凝膠結構有著密切的關系,蛋白凝膠形成的越緊密,肉糜宏觀上表現出越好的質地。但是在凍藏的過程中由于肌原纖維蛋白冷凍和氧化變性[16,40],導致肌原纖維蛋白所形成的凝膠強度也下降。
2.11 不同凍融次數豬肉糜各指標相關性及主成分分析
如表7所示,蒸煮損失與蛋白質氧化指標(羰基值),蛋白質結構變化(巰基和二硫鍵)以及蛋白質的變性程度指標(總焓變)都呈現極強的相關性(P<0.01),意味著在凍融的過程中蛋白質變性是導致蒸煮后肉糜持水性變化的關鍵因素。蛋白質變性程度越大,形成肉糜結構越粗糙,肉糜的保水能力越差。質構指標和蛋白質變性指標都有不同程度的相關性,其中巰基和二硫鍵與肉糜的硬度、彈性和內聚性都呈現了顯著相關性(P<0.05),和肉糜的咀嚼性和感官評價得分呈現極強相關性(P<0.01),總焓變和咀嚼性以及感官評分也呈現極強相關性(P<0.01),說明豬肉糜在凍融的過程中由于冷凍或者氧化變性,促使蛋白質結構發生改變,從而加熱過程中凝膠網絡結構劣化,表現出肉糜的質構隨著凍融次數的增加劣化,感官評分降低。所以說蛋白質變性指標一定程度上可以反應肉糜體系蒸煮后的感官品質。
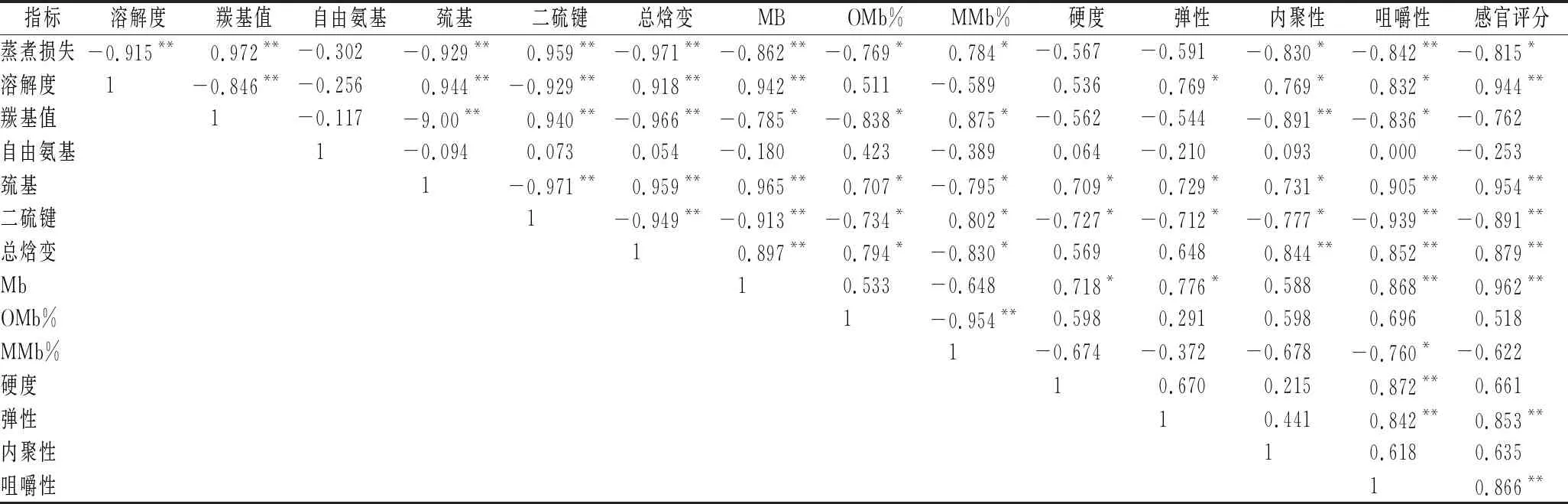
表7 不同凍融次數豬肉糜各個指標之間的相關性Table 7 Correlation among various indexes of minced pork with different freeze-thaw cycles
如圖6所示,對凍融7次的豬肉糜各項指標做主成分分析,第1主成分(PC1)的貢獻率為74.0%,第2主成分(PC2)的貢獻率為12.9%,累加貢獻率為86.9%,可充分反應不同凍融次數肉糜各指標的信息。
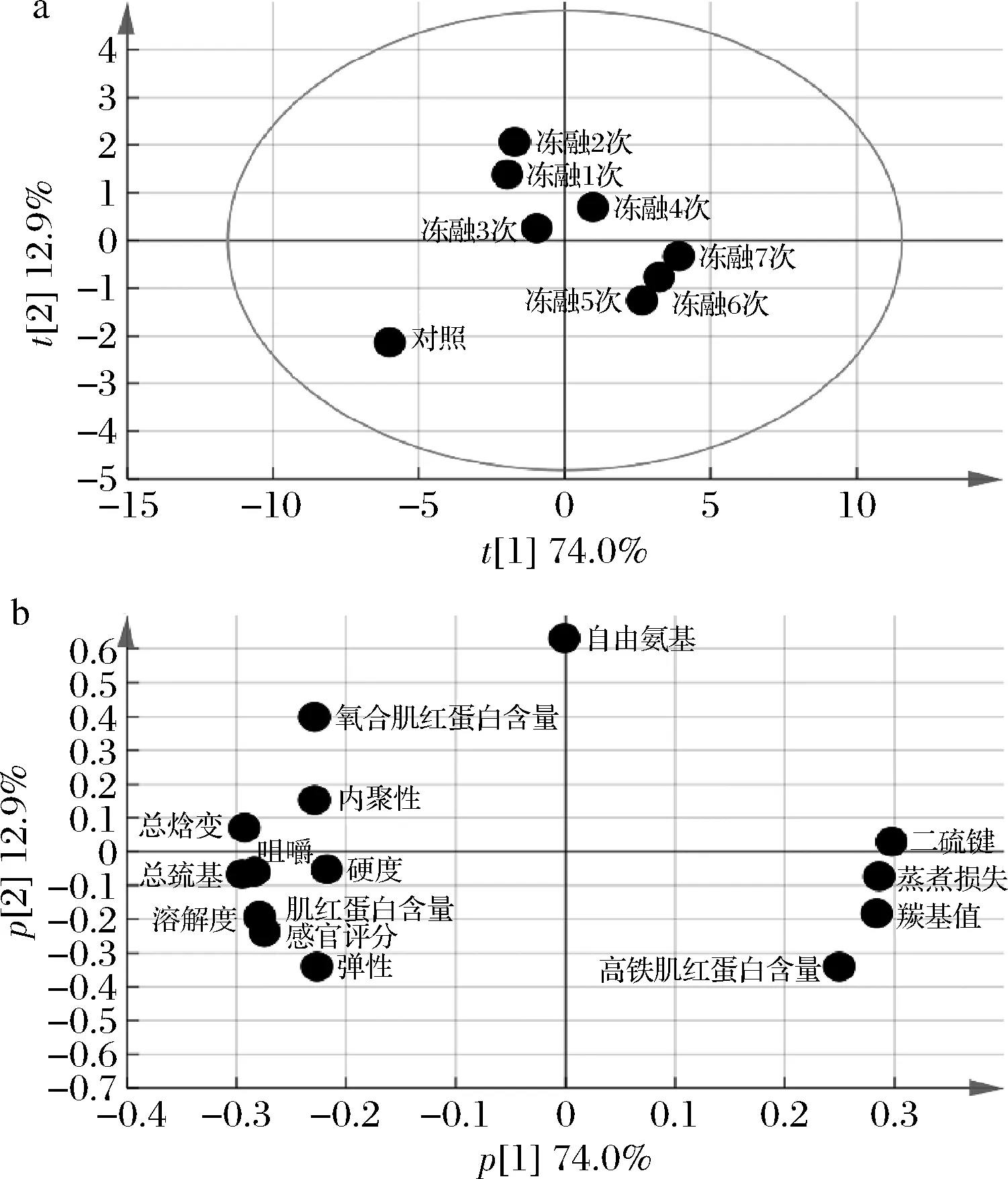
圖6 不同凍融次數豬肉糜各指標的主成分分析Fig.6 PCA of various indexes of minced pork with different freeze-thaw cycles
由得分圖6-a可知,肉糜凍融1~3次和凍融4~7次分別位于縱軸的兩側,說明在凍融4次時肉糜品質發生了顯著變化(P<0.05),在凍融5~7次時樣品點比較聚集,說明此階段變化程度減小。綜合載荷圖6-b分析,隨凍融次數的增加,樣品點沿PC1的方向向右的移動是二硫鍵、羰基值、高鐵肌紅蛋白和蒸煮損失升高造成的,而對照組在PC1方向左方和PC2方向下方主要是因為巰基、溶解度、硬度和彈性等指標值較高,所以說在凍融的過程中肉糜品質在持續的劣變。
3 結論
凍融會導致肌原纖維蛋白溶出的豬肉糜體系蛋白質變性,感官品質下降。具體表現在隨著凍融次數的增加肌原纖維蛋白羰基值和二硫鍵含量顯著地上升(P<0.05),巰基含量顯著地下降,肌紅蛋白含量顯著地下降,氧合肌紅蛋白比例呈現先升高后降低的趨勢,高鐵肌紅蛋白則相反。豬肉糜的硬度、彈性、內聚性等質構特征均下降,感官評分降低。相關性分析表明,蛋白質變性和肉糜的質構以及感官評價存在強相關性,所以一定程度上可用蛋白質變性指標來表征豬肉糜的品質,主成分分析顯示,在凍融4次后,豬肉糜的品質發生了顯著地劣變(P<0.05),所以實際生產和銷售過程中應避免發生超過3次以上的凍融,此研究為工業生產冷凍豬肉糜產品品質的保持和提升提供一定的理論依據。
