大鼠不同時期高脂飲食對子代糖脂代謝的影響及機制研究
2021-11-15 10:53:28魏銘詹迪李柱錫汪歡玉邢影羅小平
中國當代兒科雜志 2021年11期
魏銘 詹迪 李柱錫 汪歡玉 邢影 羅小平
(華中科技大學同濟醫學院附屬同濟醫院兒科,湖北武漢430030)
“健康與疾病發育起源”學說認為:宮內不良的營養環境可擾亂發育早期細胞增殖分化過程并對胎兒程序化產生永久的影響,導致成年期代謝性疾病的發生[1]。流行病學和臨床研究表明,圍生期環境如母親營養、激素水平、胎盤功能等可對胎兒的組織、器官發育產生永久性損害,孕期營養過剩可增加子代肥胖和肥胖相關疾病的風險[2-3]。
隨著經濟社會的發展,肥胖已經成為全球公共衛生危機。在世界范圍內,中國肥胖人口數量最多,成人肥胖和超重率為42%,青少年肥胖和超重率為16%[4]。育齡婦女的肥胖率隨著肥胖人口的增多而逐年升高,肥胖導致的妊娠并發癥如妊娠糖尿病、先兆子癇,以及分娩、產后并發癥的風險亦隨之增加[5-6]。妊娠期肥胖在危害孕母健康的同時,還會影響子代的肥胖易感性、胰島素敏感性等,增加子代成年后發生代謝性疾病的風險[7-8]。
研究表明,在妊娠早期、妊娠晚期等不同的發育窗口,母體肥胖對子代肥胖、代謝等有獨立的編程效應[9-10]。為了明確孕前、孕后高脂飲食干預對子代糖脂代謝的影響差異,本研究通過在孕前和/或孕期哺乳期給予高脂飲食構建妊娠期肥胖動物模型,觀察子代糖脂代謝的差異情況。
1 材料與方法
1.1 實驗動物及試劑
SPF級8周齡Sprague-Dawley(SD)大鼠,雌性40只,雄性20只,購于湖南斯萊克景達實驗動物有限公司,許可證號:SCXK(湘)2019-0004。飼養于華中科技大學同濟醫學院附屬同濟醫院動物實驗中心SPF動物房內,環境溫度20~26℃,濕度40%~70%,12 h/12 h晝夜光照,自由飲水,按實驗分組給予相應飲食。
45%高脂飼料(江蘇省協同醫藥生物工程有限責任公司);拜安康血糖儀、血糖試紙(安晟信糖尿病保健美國股份有限公司);大鼠胰島素(insulin,INS)ELISA試劑盒(北京索萊寶科技有限公司);三酰甘油(triglyceride,TG)、總膽固醇(total cholesterol,TC)含量測定試劑盒(南京建成生物工程研究所);蘇木精染液、伊紅染液、油紅O染液(武漢賽維爾生物科技有限公司);RNA提取試劑盒、逆轉錄試劑盒、PCR SYBR Green試劑、β-actin抗體、二抗(上海翌圣生物科技股份有限公司);4%多聚甲醛、蛋白酶抑制劑、RIPA裂解液、上樣緩沖液、ECL曝光液(武漢博士德生物工程有限公司);胰島素受體(insulin receptor,IR)、胰島素受體底物(insulin receptor substrate,IRS)、磷酸化蛋白激酶B(p-AKT)、膽固醇調節元件結合蛋白1c(sterol-regulatory element binding protein 1c,SREBP1c)抗體(武漢愛博泰克生物科技有限公司);絲氨酸磷酸化IRS(p-IRS)抗體(美國Thermo Fisher公司);蛋白激酶B(AKT)、過氧化物酶體增殖物激活受體α(peroxisome proliferatoractivated receptor alpha,PPARα)抗 體(英 國Abcam公司);脂肪酸合酶(fatty acid synthase,FASN)抗體(美國CST公司)。
1.2 實驗分組及妊娠期肥胖動物模型的建立
本研究通過對母鼠的飲食干預構建妊娠期肥胖動物模型[11]。8周齡SD大鼠適應性喂養1周后,將雌性SD大鼠隨機分為兩組(n=20),分別給予正常飲食、高脂飲食6周。雄性SD大鼠保持正常飲食。6周后,按雌雄2∶1合籠,次晨行陰道涂片,顯微鏡觀察到精子視為懷孕。每組剔除2只不孕、假孕的雌性大鼠后,將孕前正常飲食大鼠隨機分為CC組(孕前、孕期哺乳期均正常飲食)和CH組(孕前正常飲食,孕期哺乳期高脂飲食)(n=9);將孕前高脂飲食大鼠隨機分為HC組(孕前高脂飲食,孕期哺乳期正常飲食)和HH組(孕前、孕期哺乳期均高脂飲食)(n=9)。CH組在孕期哺乳期更換為高脂飲食,HC組在孕期哺乳期更換為正常飲食,CC組和HH組在孕期哺乳期繼續延續孕前飲食。母鼠自然分娩后,每窩隨機保留4只雄性仔鼠,進行均一化處理,避免哺乳條件、性別等的不同造成的差異。仔鼠生后3周斷奶,斷奶后全部采取正常飲食。記錄母鼠孕前、孕期體重及仔鼠體重。
1.3 檢測空腹血糖和INS水平
每組各取9只幼年期(3周)、成年期(12周)雄性仔鼠,禁食12 h,尾尖取血,血糖儀檢測空腹血糖(glucose,GLU)水平;然后腹腔內注射10%水合氯醛麻醉,暴露心臟后取血,分離血清,按照說明書要求,ELISA試劑盒檢測空腹INS水平。胰島素抵抗指數(homeostasis model assessment of insulin resistance,HOMA-IR)=[空 腹GLU(mmol/L)×空腹INS(μU/mL)]/22.5。
1.4 糖耐量試驗和胰島素耐量試驗
每組取9只仔鼠行糖耐量試驗(glucose tolerance test,GTT):3周、12周仔鼠禁食過夜,腹腔注射2 g/kg葡萄糖。尾尖取血,血糖儀檢測0、15、30、60、90、120 min血糖值。計算曲線下面積(area under the curve,AUC)。
每組取9只仔鼠行胰島素耐量試驗(insulin tolerance test,ITT):3周、12周仔鼠禁食4 h,腹腔注射0.8 U/kg胰島素,尾尖取血,血糖儀檢測0、15、30、60、90、120 min血糖值。計算AUC。
1.5 檢測肝臟TG和TC水平
每組取9只仔鼠,將采集的肝臟組織樣本研磨后提取上清用于檢測。根據說明書操作,酶標儀測定各孔光密度值,計算各組仔鼠肝臟的TG、TC水平。
1.6 肝臟蘇木精-伊紅染色和油紅O染色
蘇木精-伊紅染色:新鮮的肝臟組織在4%多聚甲醛中固定24 h,石蠟包埋后切片。石蠟切片脫蠟水化,蘇木精-伊紅染色,中性樹膠封片,光學顯微鏡下觀察。
油紅O染色:肝臟冰凍切片后干燥復溫10 min,60%異丙醇漂洗,油紅O染色,中性樹膠封片,光學顯微鏡下觀察。
1.7 RT-PCR檢測肝臟糖脂代謝關鍵基因mRNA水平
每組取9只仔鼠,利用RNA提取試劑盒提取肝臟組織RNA,分光光度計檢測RNA濃度及純度。根據逆轉錄試劑盒說明,冰上配制逆轉錄反應體系。逆轉錄儀將RNA逆轉錄為cDNA后,冰上配制PCR反應體系(10 μL):PCR SYBR Green 5 μL,上下游引物各0.2 μL,cDNA 1 μL,ddH2O 3.6 μL。PCR反應條件:95℃預變性2 min;95℃變性10 s,60℃退火/延伸30 s,40個循環。根據得到的CT值,利用2-ΔΔCT計算分析,得到相對定量的結果。引物序列見表1。

表1 RT-PCR引物序列
1.8 Western blot檢測肝臟糖脂代謝關鍵蛋白水平
每組取3只仔鼠肝臟組織,利用含蛋白酶抑制劑的RIPA裂解液裂解組織并離心取上清,加入上樣緩沖液并沸水浴加熱使蛋白變性。取等體積蛋白加樣,電泳、轉膜。PVDF膜封閉后加入一抗IR、p-IRS、IRS、p-AKT、FASN、SREBP1c(均1∶1 000稀釋),AKT、PPARα(均1∶2 000稀釋),β-actin(1∶5 000稀釋),4℃過夜,TBST洗膜3次,加入二抗(1∶5 000稀釋),室溫孵育1 h,TBST洗膜3次。ECL化學發光試劑浸泡2 min,凝膠成像儀曝光成像,以β-actin為內參,計算目的蛋白的相對表達量。
1.9 統計學分析
采用SPSS 19.0統計軟件對數據進行統計學分析,計量資料以均數±標準差(xˉ±s)表示。重復測量數據比較采用重復測量方差分析;多組間比較采用單因素方差分析,組間兩兩比較采用LSD-t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 不同飲食組母鼠孕前體重及孕期增重變化情況
兩組SD母鼠孕前體重均隨時間推移逐漸增加(P<0.001);孕前各時間點高脂飲食組母鼠體重高于正常飲食組,且孕前3周開始,高脂飲食組與正常飲食組母鼠體重差距逐漸增大(P<0.001);時間因素與分組因素存在交互作用(P<0.001)。見表2。

表2 不同飲食干預組母鼠孕前6周內體重變化比較(xˉ±s,g)
4組SD母鼠孕期體重均隨時間推移逐漸增加(P<0.001);SD母鼠懷孕后3周,HC組、CH組、HH組母鼠孕期增重均較CC組有明顯增加,且HH組母鼠孕期增重最顯著(P<0.001)。時間因素與分組因素存在交互作用(P=0.01)。見表3。
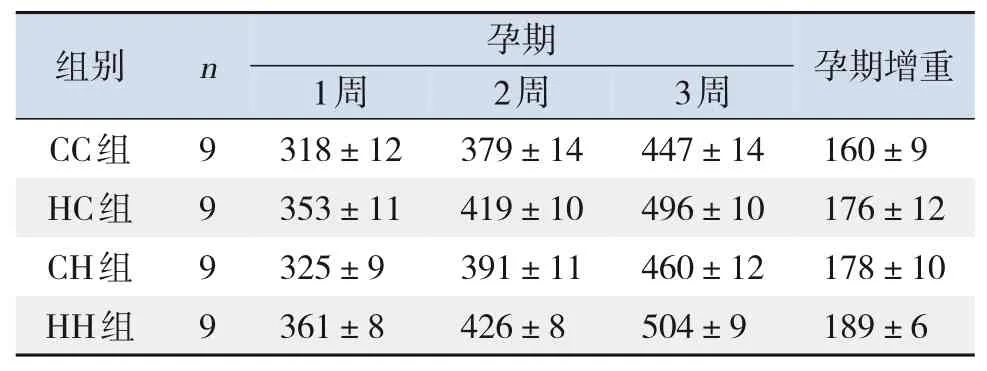
表3 各組母鼠孕期3周內體重變化比較(xˉ±s,g)
高脂飲食大鼠孕前體重增加和孕期增重過多,與正常飲食大鼠比較差異有統計學意義,提示孕前和/或孕期高脂飼料喂養均可成功構建妊娠期肥胖動物模型。
2.2 生后3周仔鼠體重及糖脂代謝情況
仔鼠生后3周時,HC組、CH組、HH組仔鼠體重均較CC組顯著增加,HH組仔鼠體重較HC組、CH組顯著增加(P<0.05),見表4。
糖代謝方面,HH組空腹GLU水平較CC組、HC組顯著增加,空腹INS水平較CC組顯著增加,HOMA-IR、GTT-AUC、ITT-AUC較其他3組均顯著增加(P<0.05),即HH組胰島素抵抗增加,糖耐量、胰島素耐量均降低(表4)。RT-PCR結果提示,HH組糖代謝通路關鍵基因IR、IRS的mRNA水平較CC組顯著降低(P<0.05),AKT的mRNA水平在4組中比較差異無統計學意義(P>0.05)(表5)。Western blot結果提示,HH組IR、IRS蛋白表達水平較其他3組顯著降低,p-IRS蛋白表達水平較其他3組顯著升高(P<0.05);CC組IR蛋白表達水平較HC組、CH組顯著升高,CH組IR蛋白表達水平較HC組顯著降低(P<0.05)。p-AKT、AKT蛋白表達水平在4組間比較差異無統計學意義(P>0.05)。見圖1和表6。
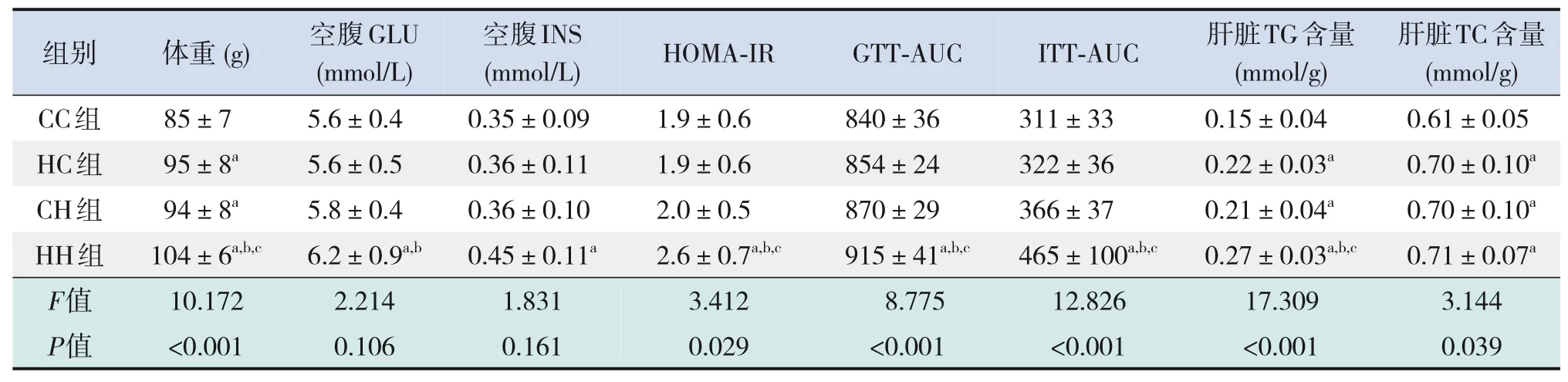
表4 各組生后3周仔鼠體重及糖脂代謝指標水平比較(n=9,xˉ±s)
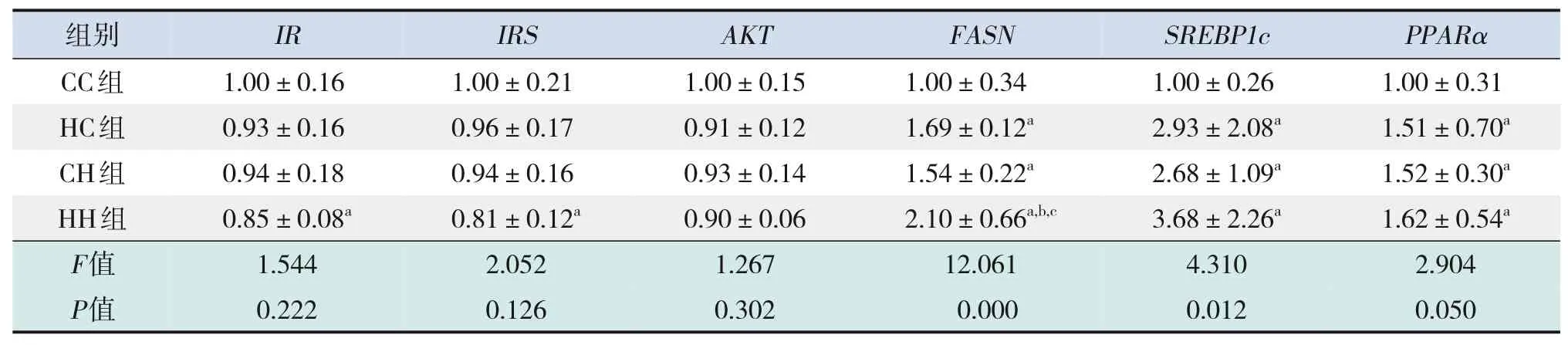
表5 各組生后3周仔鼠糖脂代謝相關基因的mRNA水平比較(n=9,xˉ±s)
脂代謝方面,HC組、CH組、HH組肝臟TG、TC水平均較CC組顯著增加,HH組肝臟TG水平較HC組、CH組顯著升高(P<0.05)(表4)。肝臟蘇木精-伊紅染色及油紅O染色結果提示,HH組肝臟空泡形成及脂質沉積最明顯,其次為HC組、CH組(圖2)。HC組、CH組、HH組肝臟脂質代謝關鍵基因FASN、SREBP1c、PPARα的mRNA水平均較CC組顯著增加,HH組FASN的mRNA水平較HC組、CH組顯著升高(P<0.05)(表5)。HC組、CH組、HH組FASN、SREBP1c、PPARα蛋白表達水平均較CC組顯著升高(P<0.05);HH組FASN蛋白表達水平較HC組、CH組顯著升高,CH組FASN蛋白表達水平較HC組顯著升高(P<0.05)。見圖1和表6。
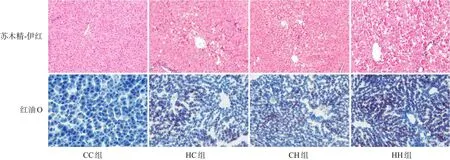
圖2 各組生后3周仔鼠肝臟病理改變(×200) 肝臟蘇木精-伊紅染色提示HH組肝細胞空泡增多,脂肪變性最明顯,其次為HC組、CH組;油紅O染色提示HH組肝臟脂質沉積最嚴重(脂滴被染為紅色),其次為HC組、CH組。
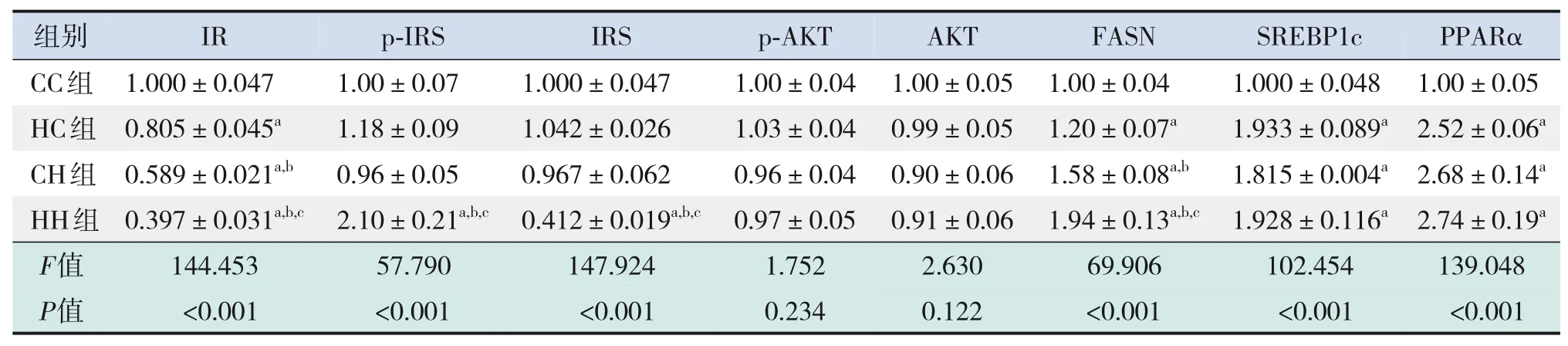
表6 各組生后3周仔鼠肝臟糖脂代謝相關蛋白表達水平比較(n=3,xˉ±s)
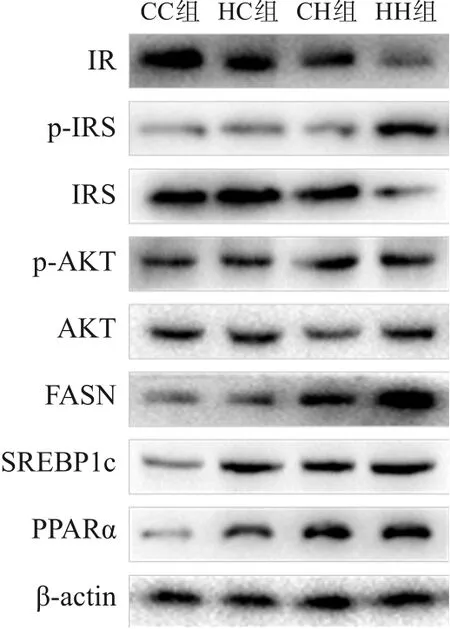
圖1 Western blot檢測各組生后3周仔鼠肝臟糖脂代謝相關蛋白表達電泳圖
2.3 生后12周仔鼠體重及糖脂代謝情況
仔鼠生后12周時,HC組、CH組、HH組仔鼠體重均較CC組顯著增加(P<0.05),見表7。
糖代謝方面,HC組、CH組、HH組空腹GLU及INS水平、HOMA-IR、GTT-AUC、ITT-AUC均較CC組顯著增加(P<0.05);HH組空腹INS水平、HOMA-IR較HC組、CH組顯著增加,GTT-AUC較HC組顯著增加(P<0.05)(表7)。RT-PCR結果提示,HC組、CH組、HH組糖代謝通路關鍵基因IR、IRS、AKT的mRNA水平較CC組顯著降低(P<0.05);HH組IRS的mRNA水平較HC組顯著降低,AKT的mRNA水平較HC組、CH組顯著降低(P<0.05)(表8)。Western blot結果提示,HC組、CH組、HH組IR、IRS、p-AKT蛋白表達水平均較CC組顯著降低,p-IRS蛋白表達水平較CC組顯著升高(P<0.05);AKT蛋白表達水平在4組中比較差異無統計學意義(P>0.05)。見圖3和表9。
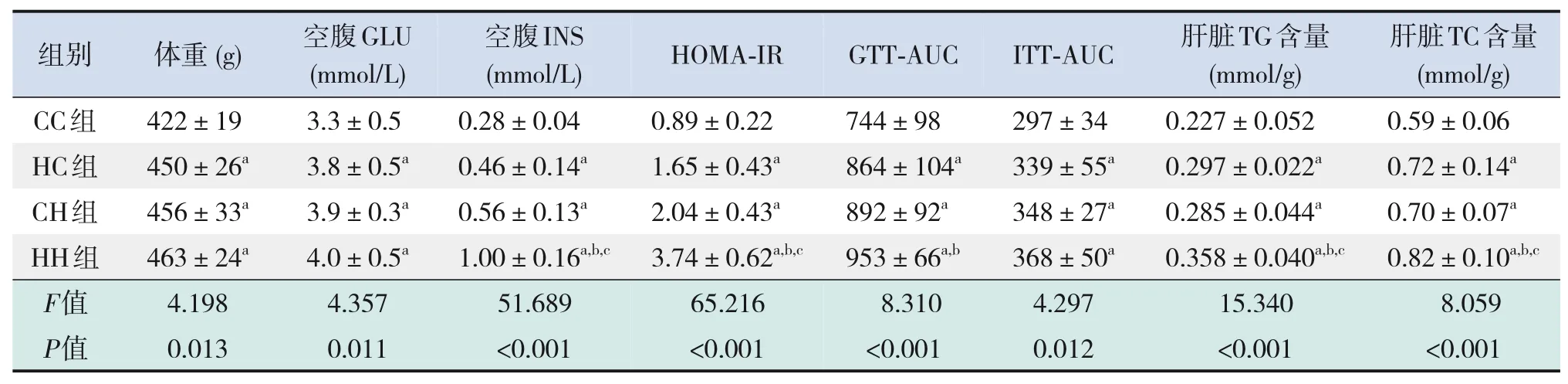
表7 各組生后12周仔鼠體重及糖脂代謝指標水平比較(n=9,xˉ±s)
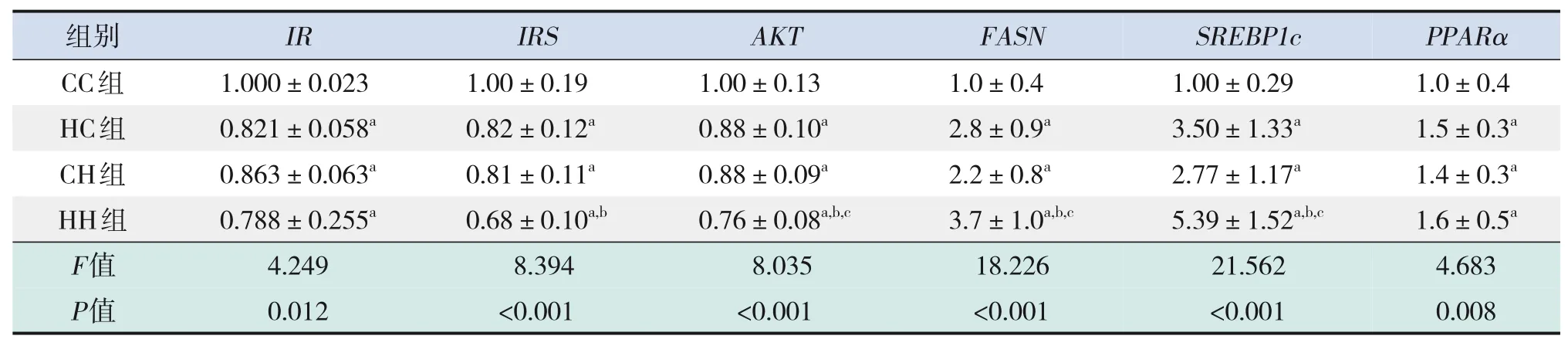
表8 各組生后12周仔鼠肝臟糖脂代謝相關基因的mRNA水平比較(n=9,xˉ±s)
脂代謝方面,HC組、CH組、HH組肝臟TG、TC水平均較CC組顯著增加(P<0.05);HH組肝臟TG、TC水平較HC組、CH組顯著增加(P<0.05)(表7)。肝臟蘇木精-伊紅染色及油紅O染色結果提示,HH組肝臟空泡形成及脂質沉積最明顯,其次為HC組、CH組(圖4)。HC組、CH組、HH組肝臟脂質代謝關鍵基因FASN、SREBP1c、PPARα的mRNA水平較CC組顯著增加(P<0.05);HH組FASN、SREBP1c的mRNA水平較HC組、CH組顯著增加(P<0.05)(表8)。HC組、CH組、HH組FASN、SREBP1c、PPARα蛋白表達水平均較CC組顯著升高,且HH組SREBP1c蛋白表達水平較HC組、CH組顯著升高(P<0.05)。見圖3和表9。
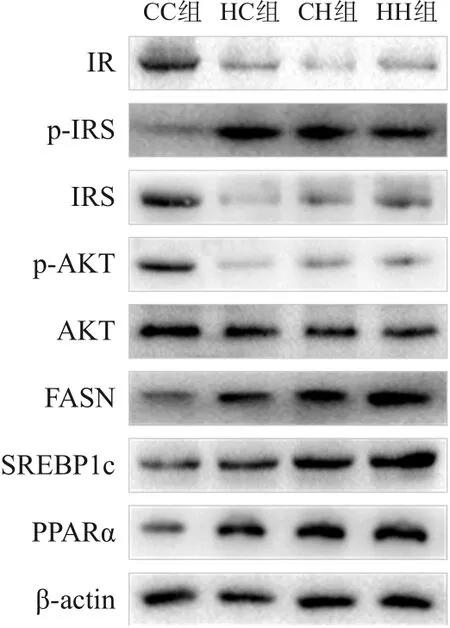
圖3 Western blot檢測各組生后12周仔鼠肝臟糖脂代謝相關蛋白表達電泳圖
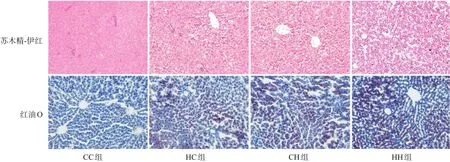
圖4 各組生后12周仔鼠肝臟病理改變(×200) 肝臟蘇木精-伊紅染色提示HH組肝細胞空泡增多,脂肪變性最明顯,其次為HC組、CH組;油紅O染色提示HH組肝臟脂質沉積最嚴重(脂滴被染為紅色),其次為HC組、CH組。
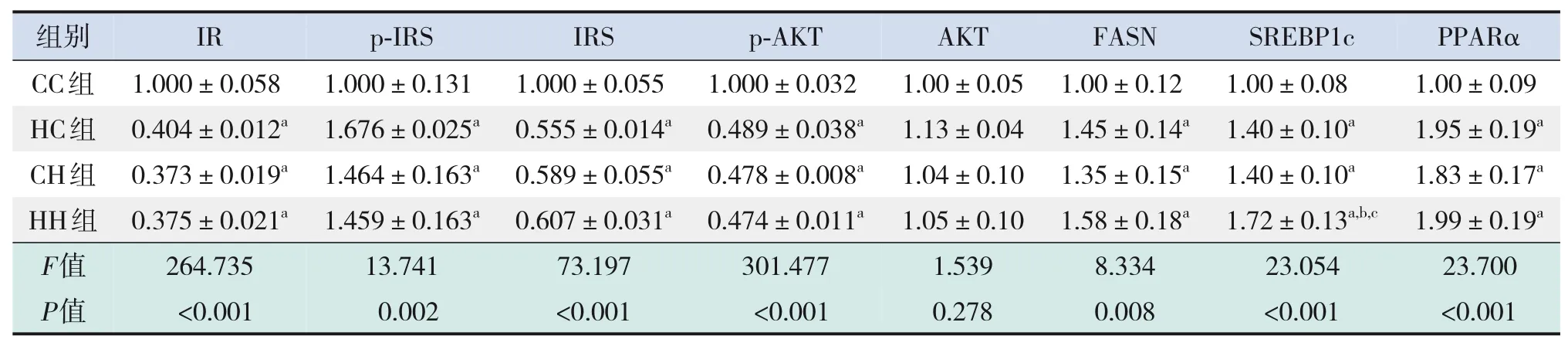
表9 各組生后12周仔鼠肝臟糖脂代謝相關蛋白表達水平比較(n=3,xˉ±s)
3 討論
在代謝研究中,干預的時機不同,可導致后代不同的表型。Zhao等[7]分別在孕期和哺乳期應用高脂飼料喂養ICR小鼠,探究分娩前后母鼠飲食的差異對子代的不同影響,研究發現,哺乳期高脂飲食組的雄性仔鼠生后21 d的肝臟TG含量較孕期高脂飲食組顯著增加。Choi[12]研究了斷奶前后母鼠和子代的飲食調整對子代成年后肥胖、胰島素抵抗等的影響,研究發現,斷奶前母鼠的高脂飲食和斷奶后子代的高脂飲食通過下丘腦促食欲基因的表達增加導致子代肥胖和胰島素抵抗,斷奶后的飲食調整可部分改善此作用。以上研究分別探討了分娩前后、斷奶前后母鼠或子代的高脂飲食對子代的影響,鑒于孕前、孕期、哺乳期母體肥胖對子代代謝的獨立編程作用,本研究重點討論了懷孕前后飲食的不同對子代糖脂代謝的影響。
由于SD大鼠體型適中便于取材,給予SD大鼠高脂飲食易構建出與人類肥胖發生過程相似的肥胖模型,故本研究中使用SD大鼠進行模型的構建。妊娠期肥胖包括孕前肥胖和孕期增重過多[13]。本研究中,孕前給予高脂飲食6周的SD母鼠,較正常飲食的母鼠,孕前體重明顯增加;孕前正常飲食或高脂飲食的母鼠,孕期給予高脂飲食,其孕期增重較正常飲食組顯著增加,說明三種方式均可成功構建妊娠期肥胖動物模型。為闡明懷孕前后的高脂飲食干預對子代糖脂代謝的影響,本研究在孕前、孕期哺乳期對SD大鼠進行高脂飼料喂養,并選取幼年期、成年期的仔鼠進行糖脂代謝研究。研究發現孕前、孕期哺乳期全程高脂飲食對子代糖脂代謝影響最大;僅孕前高脂飲食或孕期哺乳期高脂飲食對子代的影響在本研究中未見差異;孕前高脂飲食的大鼠,即使孕期哺乳期更換為正常飲食,其對子代的不利影響仍然存在;高脂飲食對子代糖脂代謝的影響考慮與糖脂代謝基因的表達改變有關。目前較少有研究強調妊娠不同時期的飲食干預對子代代謝的不同影響,本研究不僅對不同時期高脂飲食對子代的糖脂代謝影響進行了比較,而且選取了不同時期的子代進行動態觀察,本研究為后期進一步進行妊娠期肥胖相關的機制研究提供了重要依據。
本研究中,HC組、CH組子代斷奶時僅表現出脂代謝異常,成年后出現糖代謝異常,提示高脂飲食母鼠子代出現糖代謝異常的時間晚于脂代謝。GTT和ITT分別用于評估葡萄糖耐量和胰島素敏感性,AUC的增加表明葡萄糖耐量受損和胰島素敏感性下降。HOMA-IR亦被用來評估胰島素敏感性,主要預測肝臟胰島素敏感性,而ITT代表全身胰島素敏感性[14-15]。胰島素是體內唯一的降血糖激素,促進糖原生成,減少糖原分解,加速糖酵解,并抑制糖異生。胰島素抵抗是Ⅱ型糖尿病、代謝綜合征等多種慢性病的基礎,其特征是肝臟和骨骼肌等對胰島素的反應減弱[16-17]。在胰島素抵抗初期,胰島β細胞分泌胰島素代償性增加,導致高胰島素血癥,使血糖維持在正常水平。隨著胰島素抵抗的進展,胰島素水平急劇下降,機體對胰島素的反應性減弱,從而出現高血糖[18]。IR/IRS/AKT是經典的胰島素受體信號轉導通路,在糖代謝中發揮重要作用。胰島素激活IR酪氨酸激酶,從而磷酸化IRS,酪氨酸磷酸化的IRS激活AKT,通過下游因子誘導糖原合成并抑制糖異生,從而發揮降血糖的作用[18-19]。IRS總蛋白減少,絲氨酸磷酸化蛋白增加,p-AKT蛋白減少是胰島素抵抗的典型特征[20-21]。本研究中,妊娠期肥胖的子代后期均出現不同程度的糖代謝異常,考慮與胰島素通路基因IR、IRS、AKT的表達改變有關。
本研究中,高脂飲食組子代在斷奶時、成年后均表現出脂代謝異常,表現為肝臟TG、TC含量升高。TG是由長鏈脂肪酸和甘油形成的脂肪分子。FASN利用乙酰輔酶A作為引物,丙二酰輔酶A作為雙碳供體,NADPH作為還原等價物,將碳水化合物轉化為長鏈脂肪酸,是脂肪酸從頭合成中的重要調節酶[22]。SREBP1c是脂肪酸從頭合成的關鍵轉錄因子,通過上調肝臟中參與脂肪酸合成的關鍵基因的mRNA水平,轉錄激活脂肪酸合成所需的一系列酶[23-24]。PPARα是一種在肝臟中高度表達的配體激活的核受體,參與脂肪酸轉運等基因的轉錄調節,在脂質代謝中發揮重要作用[25]。本研究中,高脂飲食組子代斷奶時及成年后肝臟FASN、SREBP1c、PPARα的mRNA及其蛋白表達水平均高于正常飲食組,高脂飲食導致子代肝臟脂質沉積加重,考慮與FASN、SREBP1c、PPARα介導的脂質代謝相關。其深入機制還有待進一步探討,可能與營養過剩的宮內環境誘導的表觀遺傳學調控有關,如DNA甲基化、組蛋白修飾、非編碼RNA等[26]。
本研究中,雖然HC組與CH組相比,不同時期的子代糖脂代謝水平未見顯著性差異,但不能證明孕前高脂飲食與孕期哺乳期高脂飲食相比,其對子代健康的影響無差異。有研究應用自助餐飲食誘導母鼠孕前肥胖,并研究了孕前肥胖和孕期自助餐喂養對胎鼠的獨立影響,結果證明母鼠的肥胖狀態和孕期的自助餐喂養對子代有獨立的影響[27]。該研究應用多樣性自助餐飲食干預Wistar大鼠,與本研究中結果的不一致,可能與動物品系、飼料類型等不同有較大關系。由于本研究僅選取了幼年期、成年期兩個時間點,且每組大鼠數量有限,尚需增加觀察時間,擴大樣本數目加以驗證。
利益沖突聲明:所有作者均聲明不存在利益沖突。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
中國生殖健康(2019年2期)2019-08-23 08:12:10
人大建設(2019年12期)2019-05-21 02:55:32
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
中國全科醫學(2013年36期)2013-01-25 06:20:58
