利用葡萄枝屑、桑枝屑栽培白參菌、秀珍菇的研究
2021-11-23 04:13:14方志榮
四川農業科技 2021年10期
方志榮,林 靜,清 源,姜 英
(1.西昌學院資源與環境學院,四川 西昌 615013;2.西昌學院攀西特色作物研究與利用四川省重點實驗室,四川 西昌 615013)
白參菌(Schizophyllumcommuner)別名裂褶菌、白花、白參,真菌植物門真菌裂褶菌(Schizophyllqaeae)的一種食藥兼用的珍稀菇菌,是中溫型木腐菌,能很好地分解纖維素和木質素,并且它能夠在含有葡萄糖、纖維素、半纖維素和木質素的基質上良好生長[1]。秀珍菇(Pleurotusgeesteranus)別名鳳尾菇,在分類學上屬于真菌門、擔子菌綱、傘菌目、側耳科、側耳屬真菌,口感鮮美,營養豐富,有“菇中極品”的美譽,是目前人工栽培具有商業價值的珍貴菌種[2],生長所需的營養物質均要從栽培料中獲取。栽培白參菌和秀珍菇通常利用棉籽殼或者高粱殼、玉米芯、雜木屑和各種農作物秸稈作為主料[3-4]。2021年種植食用菌的木屑、玉米芯價格上漲,食用菌種植成本增加,急需尋找新的替代品。
涼山州立體氣候明顯,適合多種經濟作物的生長。如寧南縣2016年桑園面積達14666.7 hm2,西昌市2019年葡萄種植面積逾6666 hm2,隨著鄉村振興的推進,種植面積將進一步擴大,隨之產生大量的枝屑廢棄物。目前大部分枝屑都是直接還田,少部分作為燃料供農戶自用。雖然枝屑還田短期內在一定程度上可以改良土壤的物理性質與土壤內部的生態環境,增加土壤微生物的多樣性,提高耕地的生產力。但由于廢枝屑結構復雜,碳/氮比高,不易降解,從長遠角度看,將影響土壤墑情和耕作。此外,未腐爛的枝屑將大量的致病菌和蟲卵帶入土中,造成致病菌和蟲卵大量繁衍,給葡萄樹和桑樹的生長帶來不利的影響。如果合理利用這些廢枝屑,不但可以降低種植食用菌的成本,也能提高廢棄物的利用率,變廢為寶,實現資源的合理利用。有關測定表明,葡萄枝屑含粗蛋白5.57%,粗纖維64.38%,粗灰分3.81%,有機質77.35%,也含有豐富的鉀、鈣等礦質元素[5];桑枝屑含粗蛋白5.44%,纖維素51.88%,半纖維素23.02%,木質素18.18%,粗灰分1.57%,同時也含有鉀、鈣、鎂等礦質元素[6]。大量研究表明可采用葡萄枝屑和桑枝屑作為基質栽培食用菌[7-13]。利用葡萄枝屑栽培白玉菇[7],平菇[4],杏鮑菇[8-9]、香菇[10];利用桑枝屑生產猴頭菇[11]、平菇[12]、香菇[13]利都已獲得成功。但是目前利用葡萄枝屑或者桑枝屑栽培白參菌和秀珍菇的報道尚少,本研究以葡萄枝和桑枝為栽培主料,研究不同培養料中白參菌、秀珍菇胞外酶活性、生物學轉化率及營養成分的差異,以期為生產上利用葡萄枝和桑枝栽培白參菌和秀珍菇提供參考。
1 材料與方法
1.1 試驗材料
白參菌菌種、秀珍菇菌種購于貴州習水縣食用菌研究所,葡萄枝屑和桑枝屑購于冕寧縣農戶。
1.2 試驗設計
試驗選擇農戶種植秀珍菇最常用的基質配方作為對照(CK),用葡萄枝屑和桑枝屑替代玉米芯,3種處理的基質配方如下:配方1:葡萄枝屑50%+高粱殼30%+麥麩16%+石灰1%+石膏2%+蔗糖1%;配方2:桑枝屑50%+高粱殼30%+麥麩16%+石灰1%+石膏2%+蔗糖1%;配方3(CK):玉米芯50%+高粱殼30%+麥麩16%+石灰1%+石膏2%+蔗糖1%。每個配方栽培100袋。胞外酶和營養成分測定每次從不同配方的選取3袋,鮮菇產量選取10袋測定。
1.3 菌棒制作及栽培管理方法
制作菌棒時,按照上述配方比例稱取原材料,拌料均勻,采用16cm×32cm的聚乙烯袋裝袋,每袋500g(干料)。按常規方法進行菌絲培養及出菇管理。
1.4 生物學相關指標及酶活性測定
1.4.1 營養生長階段相關指標測定 采樣:每一個配方隨機選取3袋樣品,測定營養生長期菌絲體CMC-Na酶、半纖維素酶、漆酶、淀粉酶、過氧化物酶的活性。
粗酶液的提取:20g發菌料加蒸餾水100mL,25℃下浸提4h,過濾后,4000轉/min離心10min,上清液即為粗酶液。另取20g發菌料于80℃的鼓風干燥箱中烘干至恒重,采用上述方法提取上清液作為對照[14]。
CMC-Na酶活性的測定采用DNS比色法[15]。CMC-Na酶的活力單位為:在50℃的條件下反應1min產生相當于1μg分子葡萄糖的還原糖的酶量。
半纖維素酶的測定采用DNS比色法[16]。半纖維素酶的活力單位為:在50℃的條件下反應1min產生相當于1μg分子葡萄糖的還原糖的酶量。
漆酶的測定采用鄰甲苯胺氧化法[14]。定義漆酶活力單位:1g干物質中的酶含量單位時間內使OD600值改變0.01定義為一個活力單位。
淀粉酶的測定采用DNS比色法[17-18],定義淀粉酶活力單位:1g干物質與底物反應1min釋放1μg葡萄糖定義為一個活力單位。
過氧化物酶的測定采用愈創木酚法[19],定義過氧化物酶活性單位:1g干物質中的酶含量單位時間內使OD470值改變0.01定義為一個活力單位。
1.4.2 生殖生長階段相關指標測定 生殖生長階段胞外酶活性的測定:按照上述方法采集發菌料,測定第一茬菇和第二茬菇菌絲體和子實體階段胞外酶CMC-Na酶、半纖維素酶、漆酶、淀粉酶、過氧化物酶的活性。
1.4.3 子實體產量和營養成分測定 待白參菌和秀珍菇子實體七八分成熟采收。記錄每個配方下白參菌和秀珍菇的單袋第一茬菇鮮菇產量和第二茬菇鮮菇產量,同時計算不同配方第一茬菇和第二茬菇的生物學效率,采集第一茬菇和第二茬菇進行干物質、總糖和粗蛋白的分析。
生物學效率(%)=鮮菇重(g)/干料重(g)×100%[20]
每個配方采集10個子實體,記錄子實體的鮮重,將其放入鼓風干燥箱中105℃烘10min,后在80℃的條件下烘干至恒重,測其干物質含量。將烘干后的子實體粉碎,過10目篩,材料用于測定粗蛋白和總糖含量。粗蛋白含量的測定:采用納氏比色法[21]。總糖的測定:采用蒽酮比色法[22]。
1.5 數據統計與分析
利用SPSS 23.0對胞外酶活性和子實體營養成分進行方差分析,并用LSD進行差異顯著性檢驗。
2 結果與分析
2.1 不同配方對白參菌和秀珍菇各個生長階段胞外酶活性的影響
在白參菌、秀珍菇的生長發育過程中,菌絲體會產生羧甲基纖維素酶、半纖維素酶、漆酶、過氧化物酶、淀粉酶等分泌到菌絲體外將培養料中的纖維素、半纖維素、木質素、淀粉、果膠等大分子物質水解成小分子物質供菌絲體、子實體吸收和利用。
不同配方對白參菌第一茬菇不同生長階段胞外酶活性的影響見表1。羧甲基纖維酶是一類將纖維素分解為葡萄糖的酶,各處理在菌絲體、原基、子實體生長階段羧甲基纖維素酶的活性大小的順序為:子實體階段>原基階段>菌絲體階段。CK羧甲基纖維素酶的活性顯著高于配方1和配方2,配方1羧甲基纖維素酶的活性顯著高于配方2。半纖維素酶可將半纖維素分解為五碳糖、六碳糖和糖醛酸,各處理在菌絲體、原基、子實體生長階段羧甲基纖維素酶的活性大小的順序為:菌絲體階段>原基階段>子實體階段。CK半纖維素酶的活性顯著高于配方1和配方2,配方1半纖素酶的活性顯著高于配方2。漆酶和過氧化酶是木質素分解相關的酶,可將木質素分解為復雜的小分子化合物,各處理漆酶活性的大小表現為配方3(CK) >配方1>配方2。在原基生長階段,漆酶的活性達到最高值,在子實體生長階段漆酶活性下降。各處理漆酶活性的大小表現為配方3(CK) >配方1≈配方2。在原基生長階段,過氧化物酶的活性達到最高值,在子實體生長階段漆酶活性下降。淀粉酶可將小分子的碳水化合物分解為葡萄糖,各處理在菌絲體、原基、子實體生長階段淀粉酶活性大小的順序為:菌絲體階段>原基階段>子實體階段。CK淀粉酶的活性顯著高于配方1和配方2,配方1半纖素酶的活性顯著高于配方2。
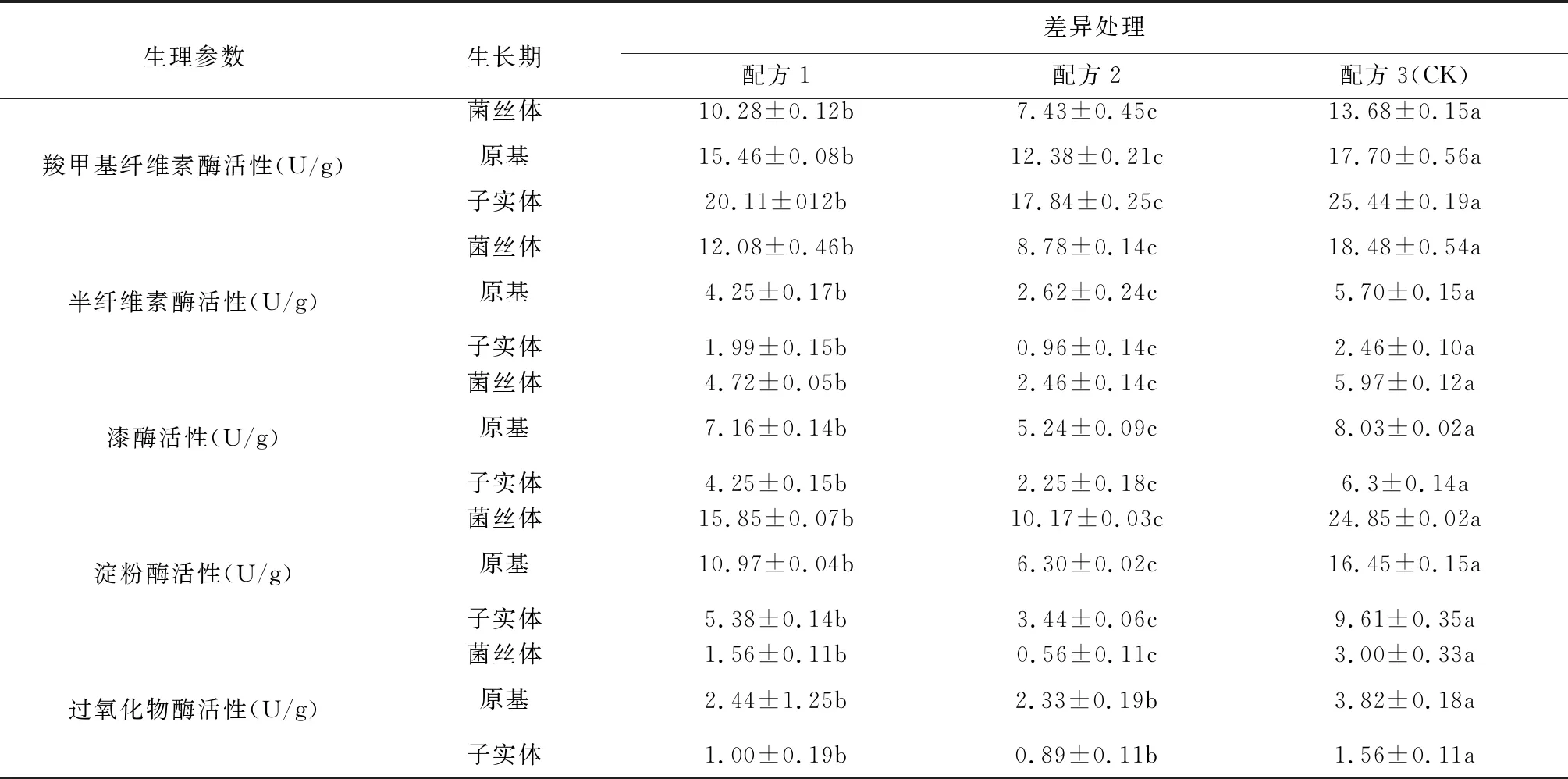
表1 不同配方對白參菌第一茬菇不同生長階段胞外酶活性的影響
不同配方對秀珍菇第一茬菇不同生長階段胞外酶活性的影響見表2。各處理在菌絲體、原基、子實體生長階段羧甲基纖維素酶的活性大小的順序為:子實體階段>原基階段>菌絲體階段。CK羧甲基纖維素酶的活性顯著高于配方1和配方2,配方1羧甲基纖維素酶的活性顯著高于配方2。各處理在菌絲體、原基、子實體生長階段羧甲基纖維素酶的活性大小的順序為:菌絲體階段>原基階段>子實體階段。CK和配方2半纖維素酶的活性顯著差異,二者顯著高于配方1。各處理在菌絲體、原基、子實體生長階段羧甲基纖維素酶的活性大小的順序為:菌絲體階段>原基階段>子實體階段。各處理漆酶活性的大小表現為配方3(CK) >配方2>配方1。配方1菌絲體、原基、子實體生長階段過氧化物酶無明顯的變化,配方2、CK菌絲體和原基階段過氧化物酶無明顯變化,但在子實體階段過氧化物酶達到最大值,CK的過氧化物酶活性在子實體階段顯著高于配方2。各處理淀粉酶活性的大小表現為配方3(CK) >配方2>配方1。在原基階段淀粉酶的活性最低,在子實體階段淀粉酶活性有所上升,但活性仍低于菌絲體階段。
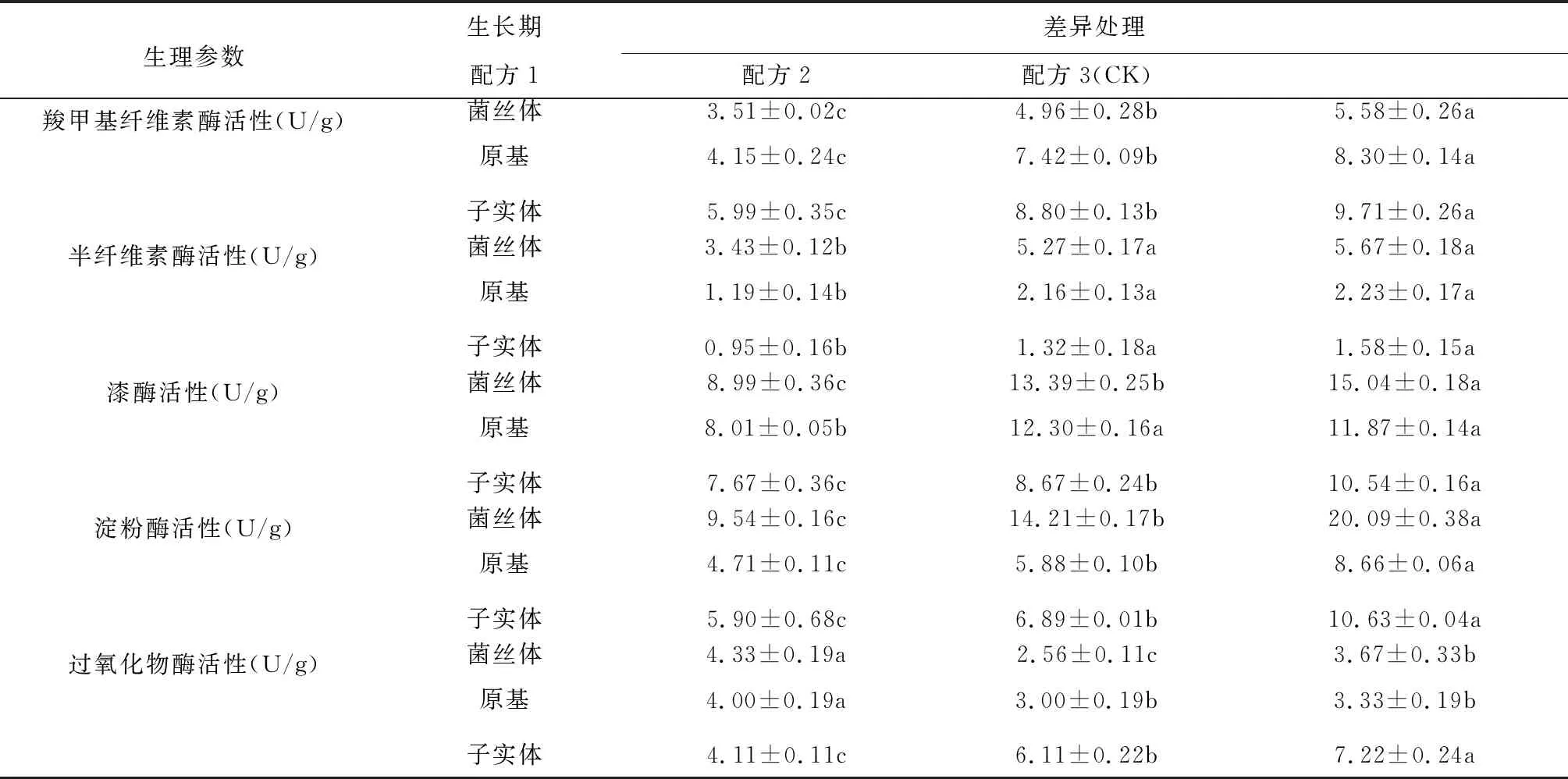
表2 不同配方對秀珍菇第一茬菇不同生長階段胞外酶活性的影響
2.2 不同配方對白參菌和秀珍菇生物學效率和營養成分的影響
由表3可知,在不同栽培配方下,白參菌和秀珍菇生長所需時間都不同。在3個配方中,配方3白參菌的菌絲滿袋時間、形成原基時間、現蕾時間和第一次采收時間最短,其次是配方1。配方3秀珍菇的菌絲滿袋時間、形成原基時間、現蕾時間和第一次采收時間最短,其次是配方2。結果表明:利用葡萄枝屑作為主料栽培白參菌生長時間比利用桑枝屑作為主料栽培的更短,但是利用桑枝屑作為主料栽培秀珍菇生長時間比利用葡萄枝屑作為主料栽培的更短。
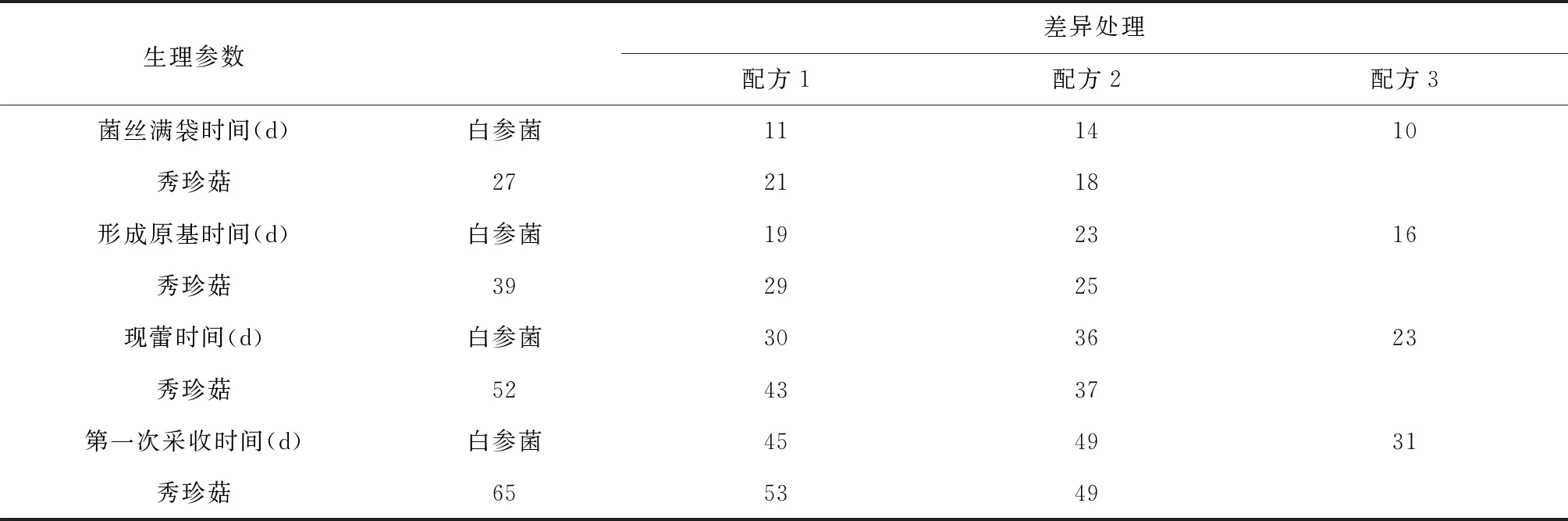
表3 不同配方對白參菌和秀珍菇生長時間的影響
由表4可知,白參菌第一茬菇、第二茬的鮮菇產量、生物學效率、干物質、總糖和粗蛋白含量在不同配方中均具有顯著差異。在3個配方中,配方1的第一茬菇的產量和生物學效率高于配方2,干物質和總糖含量最高的均是配方1,配方1的粗蛋白含量高于顯著配方2,但低于配方3(CK)。配方1的第二茬菇的產量和生物學效率也高于配方2,僅次于配方3(CK),第二茬菇配方1的總糖含量最高,粗蛋白含量也顯著高于配方2,但低于配方3(CK)。結果表明:利用葡萄枝屑種植的白參菌鮮菇產量、生物學效率和粗蛋白顯著高于桑枝屑種植的白參菌。
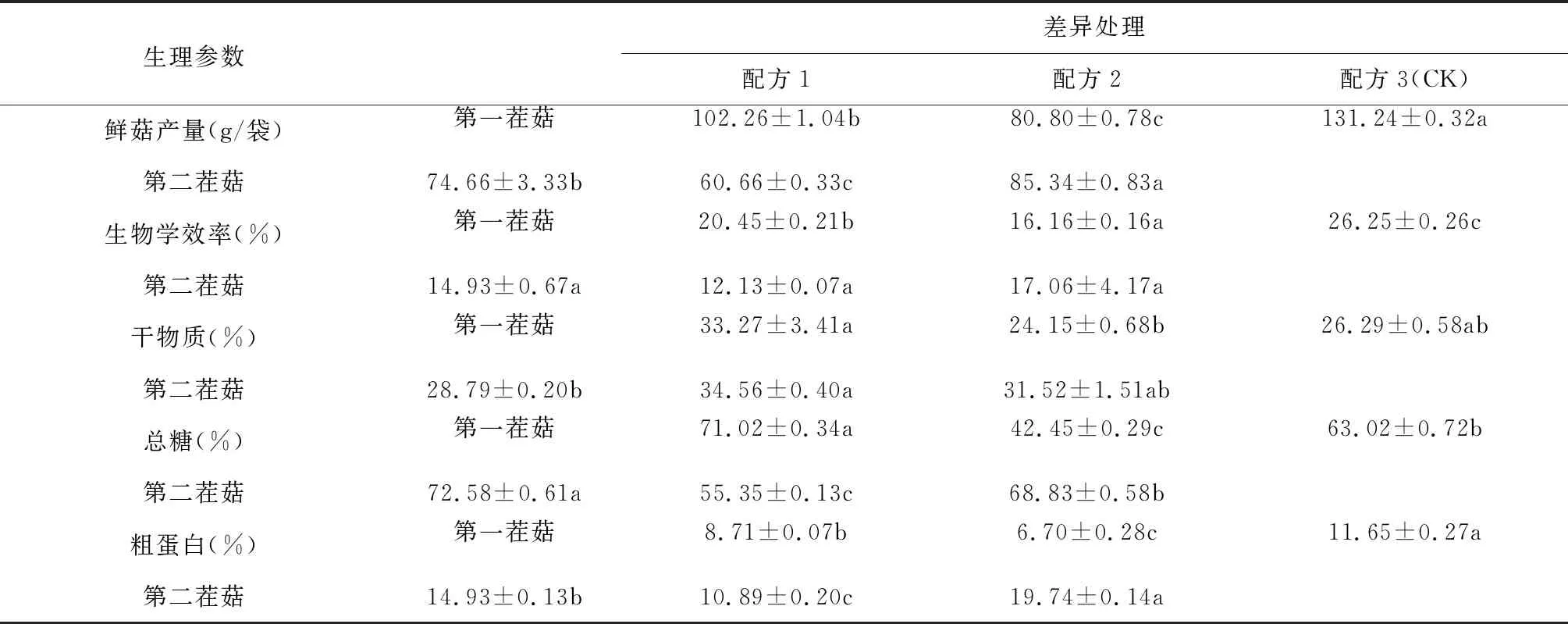
表4 不同配方對白參菌生物學效率和營養成分的影響
在不同配方中秀珍菇第一茬菇、第二茬菇的鮮菇產量、生物學效率、干物質、總糖、粗蛋白含量具有顯著差異(見表5)。在3個配方中,配方2的第一茬菇的鮮菇產量和生物學效率高于顯著配方1,低于配方3(CK)。配方2的第二茬菇鮮菇產量和生物學效率在3個配方中是最高的。干物質含量最高的是配方2,其次是配方3,但是配方2與配方3差異不顯著,配方3總糖含量顯著高于配方1和配方2,配方2的粗蛋白含量最高。第二茬菇配方2的鮮菇產量和生物學效率由于其它2個配方,干物質含量低于其它2個配方,總糖和粗蛋白含量由于其它2個配方。結果表明:利用桑枝屑種植的秀珍菇的鮮菇產量、生物學效率和粗蛋白顯著高于葡萄屑種植的秀珍菇。
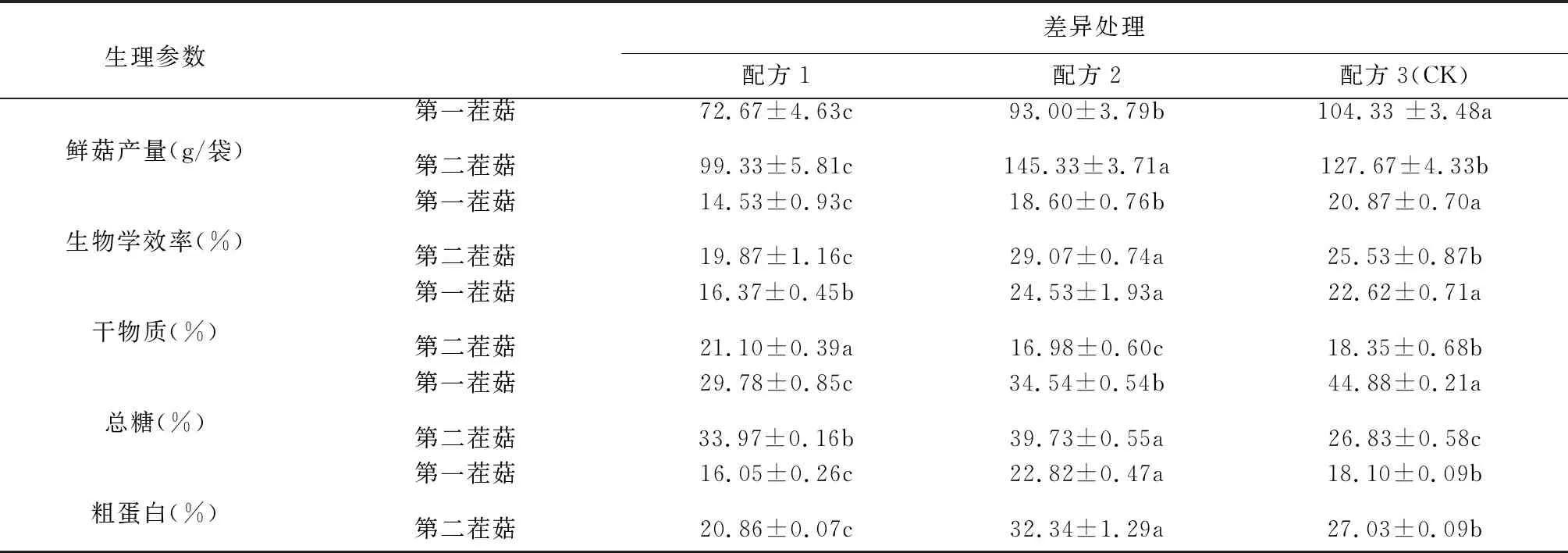
表5 不同配方對秀珍菇生物學效率和營養成分的影響
3 結果與討論
不同配方栽培白參菌和秀珍菇的結果表明,由于配方的差異導致胞外酶活性、生物學效率、粗蛋白等出現差異。研究表明,食用菌胞外酶活性的大小影響其自身的分解能力[23]。同時也有研究表明,不同的栽培配方,由于生長環境營養物質的不同,對酶活性大小影響較大,但對變化趨勢影響較小[19]。葡萄枝屑和桑枝屑含有豐富的纖維素和半纖維素等大分子營養物質,但是不能直接被食用菌吸收利用,只有通過其自身分泌的相應胞外酶將其大分子物質分解成葡萄糖小分子物質,供給食用菌生長所需要的碳源,由此可見,胞外酶的活性大小在食用菌各個生長階段發揮著至關重要的作用。同時有研究結果說明,秀珍菇的纖維素酶活性、半纖維素酶活性、漆酶活性、淀粉酶活性和過氧化物酶活性的提高能夠促進秀珍菇的生長發育[13]。張曉昱[24]通過研究香菇胞外酶活性與其自身生產發育的關系得出,香菇胞外酶活性相對高一點的其產量相對較高,營養成分相對較好。因此,在本研究中配方1、配方2、配方3同時種植白參菌和秀珍菇,配方3(CK)胞外酶的活性顯著高于配方1和配方2,因而對纖維素、半纖維素、木質素等大分子的分解更為徹底,表現為更高的生物學效率。配方1白參菌各個生長階段的胞外酶活性大體上要高于配方2,因而白參菌的生物學效率配方1高于配方2。配方2秀珍菇各個生長階段胞外酶活性大體上要高于配方1,因而秀珍菇的生物學效率配方2高于配方1。由此說明,在其它輔料相同的情況下,利用葡萄枝屑為主料栽培白參菌的效果優于桑枝;利用桑枝屑為主料的栽培料栽培秀珍菇的效果優于葡萄枝屑。
白參菌在碳氮比為10:1~100:1的條件下均能生長,最適生長C/N為40:1[1],葡萄枝屑C/N為56.08[5]與40:1相接近,所以利用以葡萄枝屑為主料栽培白參菌是可行的。秀珍菇適宜生長的C/N比為20-30:1[25],桑枝屑的C/N比在23:1左右[6],因此利用以桑枝屑為主料的栽培料栽培秀珍菇是可行的。忻龍祚等[26]利用葡萄枝屑栽培平菇,發現平菇產量高,生物學效率大,且葡萄枝屑可以提高平菇中鋅、硼等微量元素的含量,賴氨酸的含量也有所提高,可以滿足人體對氨基酸的需求。陳嬌嬌等[27]利用葡萄枝屑栽培姬菇也表現出生物學效率和產量高,且肉質厚實。研究表明利用桑枝屑栽培平菇,產量高[12]。徐建俊[13]通過桑枝屑香菇與雜木屑香菇營養成分比較得出,生物學效率比雜木屑高1.7%,粗蛋白是雜木屑香菇的1.03倍。曹澤彬等[28]研究桑枝屑生產香菇發現第1、2潮菇的產量與對照對比較高,且粗蛋白、粗纖維和多糖含量也較高,粗脂肪含量較低。由表4和表5,配方1和配方2同時與配方3(CK)對比,白參菌的鮮菇產量、生物學效率、干物質、總糖和粗蛋白含量在配方1中高于配方2,然而秀珍菇的鮮菇產量、生物學效率、總糖含量在配方2中高于配方1。由此說明配方1與配方2相比較,配方1種植出的白參菌產量更高和營養成分更好,然而配方2種植出的秀珍產量更高和營養成分更好。
雖然配方3白參菌和秀珍菇表現最佳,但配方1和配方2種植的白參菌和秀珍菇仍有較高的生物學效率。本試驗的研究結果表明:生產上可用葡萄枝屑代替玉米芯種植白參菌,可用桑枝屑代替玉米芯種植秀珍菇。
