佛手和代代酸橙授粉對香櫞果形的花粉直感效應研究
2022-04-13 05:14:56劉金蓮王小青徐勁劍梅煜琳廖芳蕾陳文榮郭衛東
浙江師范大學學報(自然科學版) 2022年2期
關鍵詞:實驗
劉金蓮, 王小青, 徐勁劍, 梅煜琳, 廖芳蕾,2, 陳文榮,2, 郭衛東,2
(1.浙江師范大學 化學與生命科學學院,浙江 金華 321004;2.浙江省特色經濟植物生物技術重點實驗室,浙江 金華 321004)
0 引 言
果形是園藝作物的重要生理性狀,是食用價值和觀賞價值的外在表現指標之一[1],果形建成不僅影響果實的外觀形態,也與果實質量、果實品質等密切相關[2],研究果形的建成對改變果形、提高果實品質有重要意義.花粉直感效應對許多園藝作物的果形產生顯著的影響[3-4],且花粉中的遺傳因子會對果形的發育起到重要調控作用[5-6].
目前果形發育的研究發現,番茄中SUN,CNR等基因分別在果實發育的不同階段調控果形建成[7].SUN,YABBY基因主要在果形建成初期,即花芽分化和發育期中起至關重要的調控作用[7-8].實驗證明,SUN作為正向調控因子編碼影響果實伸長的基因[9-10],能夠有效控制番茄果形指數[7].SUN在授粉后開始啟動[11].KAN2是心皮發育相關基因,該基因啟動表示心皮發育在進行[12].CNR是控制胎座發育的基因,PavCNR12可控制甜櫻桃果形,而與其高度相似的PbCNR6也能控制梨果形[13-14].CRC能夠抑制花的分生組織發育,并且調控雌蕊發育[15].柑橘是世界上產量最高的果樹,也是我國第一大水果,其果形相關基因研究領域存在較多空白,對花粉影響果形發育的機制和果形遺傳方面的研究相對較少.
佛手、香櫞同為蕓香科柑橘屬物種,但在果形上卻存在極大差異[16].香櫞(CitrusmedicaL.)果形一般呈現為橢圓形、近圓形或兩端狹的紡錘形[17],表面粗糙.佛手(Citrusmedicavar. L.sarcodactylisSwingle)是香櫞的變種,果形頂端呈指狀開裂[18].研究果形需要對果實外觀進行準確描述,通常會采用果形指數作為參考,即測量果實橫縱徑.果形指數是衡量蘋果外觀質量的主要指標,通常果形指數越接近1,表明該果實越端正[19].已有研究表明,梨[20]、獼猴桃[21]等外形規則的果實,在生長發育過程中,果形指數會發生變化,且同一物種不同品種間果形指數同樣也存在差異[22].而對于佛手等不規則形態的果實,則需測量果實最長橫徑和最寬縱徑,對果實進行跟蹤測量.
本實驗室前期研究發現,佛手和香櫞果形差異從花芽發育時期開始.在佛手和香櫞開花當天花和葉的組織中,檢測到SUN家族基因及YABBY家族基因在雌蕊上的表達差異[18],推測上述家族其他基因也可能會參與雌蕊到果實的形態建成.通過預實驗已檢測CNR,CRC,SUN,KAN2,YABBY2,YABBY5等基因在香櫞和佛手的花芽發育階段存在表達差異,因此,本文將以上述基因為參考繼續探討果形差異和基因表達差異的相關性.代代酸橙(C.aurantiumvar.daidaiTanaka)的果形在經過園藝栽培后有諸多形態,本文選擇以果頂有較深的放射溝直達果基,果實表面有凹凸縱向紋路的代代酸橙為研究材料.研究以佛手、香櫞和代代酸橙這3種花粉為父本,以香櫞為母本進行授粉,對香櫞果形發育的形態學數據追蹤觀測,在各授粉組香櫞的雌蕊的mRNA檢測上述5種果形建成相關基因的表達.通過研究佛手、代代酸橙花粉對香櫞果形的影響,以期了解柑橘屬花粉直感對果形發育的影響及可能的作用機理,為蕓香科植物果形研究提供新的思路.
1 材料與方法
1.1 實驗材料
選擇生長狀況一致且無病蟲害的三年生香櫞共10株,作為本次實驗的母本材料,對其進行適當修剪,每株保留相同數目的花芽,進行相同的水肥管理.收集開花當天的佛手、香櫞及代代酸橙開放花的新鮮花粉作為本次實驗的父本材料.實驗于2018年2月—2018年12月在大棚內進行.
1.2 實驗方法
1.2.1 組別設置
在大小合適的3組香櫞花序上用鑷子除去多余的雄花和已開的雌花,保留同樣數目的雌花,并對其進行前期套袋處理和分組掛牌標記,防止其他花粉對實驗產生干擾.
每個花粉授粉組選擇3株香櫞為母本,設1,2,3株為香櫞花粉授粉組,4,5,6株為佛手花粉授粉組,7,8,9株為代代酸橙花粉授粉組,其中第3,6,9株用作觀察,其余植株用作取樣;第10株香櫞不作處理,用作空白組的取樣拍照等.
1.2.2 花粉的采集凍存和授粉套袋
在佛手、香櫞、代代酸橙開花的當天,用鑷子剝離花藥,置于26~28 ℃條件下24 h,待花粉自然散粉后,收集成熟的花粉輕柔放入離心管中,在離心管上標明取樣日期及花粉種類.取樣結束后迅速將花粉放入冰盒中,再放入-80 ℃的冰箱中低溫凍存.
對前期去雄套袋處理的花芽進行授粉,將佛手、香櫞和代代酸橙花粉分別授粉香櫞柱頭(花粉在柱頭上均勻涂抹3次,盡量使每個柱頭上的花粉數量一致),授粉處理完畢后重新套袋,并進行掛牌標記.
1.2.3 果形觀察及生長數據追蹤記錄
定期觀察并拍照記錄3組花粉授粉組果實形態的變化,用游標卡尺分別測量果實的橫徑和縱徑,果形指數為縱徑和橫徑的比值.觀察果實的顏色變化、突起、褶皺的有無等形態特征,及時記錄香櫞果實有落果或者生長狀況不佳的情況.
1.2.4 香櫞雌蕊的收集和凍存
參考前期栽培經驗,將香櫞花期分為開花前7天、開花授粉當天及授粉后7天共3個時期進行取樣,并將樣本置于-80 ℃冰箱中冷凍保存.
1.2.5 熒光定量PCR
根據Trizol C Invitrogen試劑盒的方法提取事先凍存的3種花粉和各組花芽樣本的RNA,參照Vazyme公司的HiScript?Reverse Transcriptase反轉錄說明書合成轉錄的特定基因的cDNA第一鏈,β-actin驗證cDNA產物.
使用生物信息學技術設計果形相關基因序列,使用Primer Premier 5.0軟件設計引物,引物序列合成于生工生物工程有限公司(上海).
以各樣品cDNA為模板,β-actin作為內參基因,在模板內分別加入不同目的基因引物,按照SYBR? Premix Ex TaqTM PCR Kit試劑盒(艾德萊,北京)配制反應體系,通過熒光定量PCR儀(型號:ABI PRISM 7000)進行實時熒光定量PCR實驗.實時熒光定量的程序為:95 ℃預變性5 min,95 ℃ 30 s,退火溫度30 s,72 ℃ 30 s,40個循環,反應體系為20 μL,含200 ng cDNA,10 μL SYBRGreen PCR mix,10 μM的上下游引物各0.8 μL,最后用雙蒸水補足.每個基因和每個樣品均進行3個生物學重復,采用2-△△CT方法計算基因相對表達量.
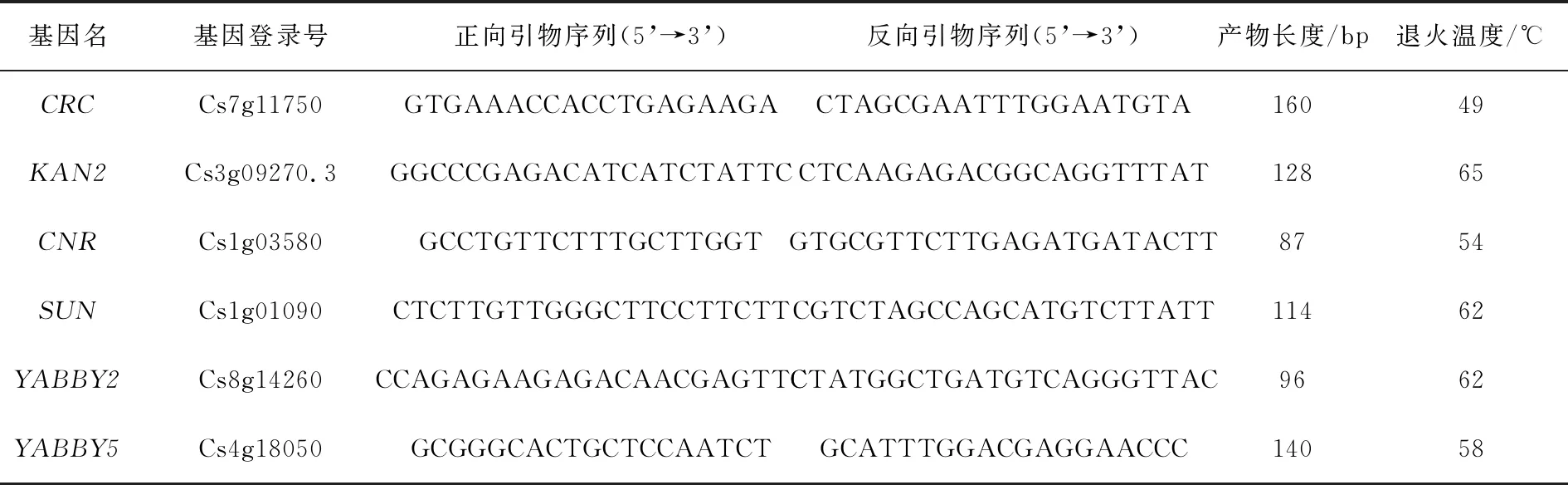
表1 熒光定量 PCR 引物
1.3 統計分析
使用SPSS 19.0進行統計分析,SigmaPlot 14.0繪制圖表,采用Duncan檢驗進行單因素(One-way ANOVA)方差分析(P<0.05).
2 結果與分析
2.1 香櫞、佛手、代代酸橙的果形比較
在果實成熟期,香櫞果形一般呈果頂果臍尖的紡錘形,佛手具有指狀的開裂.代代酸橙的果形具有縱向突起,全果有瘤狀褶皺,但縱向突起不分裂的形態.以上3種果形如圖1所示.在預實驗中發現,與佛手和代代酸橙種在一起的香櫞果皮更加粗糙,因此,在第2年對該香櫞進行去雄套袋處理,觀察不同花粉授粉后的果形是否確實發生了變化.

圖1 香櫞、佛手、代代酸橙果形圖
2.2 不同授粉組香櫞的果形差異
經過2年授粉的統計數據表明,在佛手授粉和代代酸橙授粉組,均有90%以上的果形出現變異.從形態外觀上比較,自交授粉香櫞果形呈現為中間圓,頂端狹的紡錘形,果皮表面較光滑(見圖2a~2b).代代酸橙花粉授粉后的香櫞表面出現縱向輕微褶皺突起現象(見圖2c~2d).佛手花粉授粉后的香櫞相對更加狹長,表面粗糙,果皮不光滑程度大大加深,除了縱向的褶皺突起,突起部分不連續,導致果皮上出現瘤狀突起(見圖2e~2f).果形指數顯示,代代酸橙與佛手授粉的果實果形指數偏小(見圖2g),即果形偏扁平.
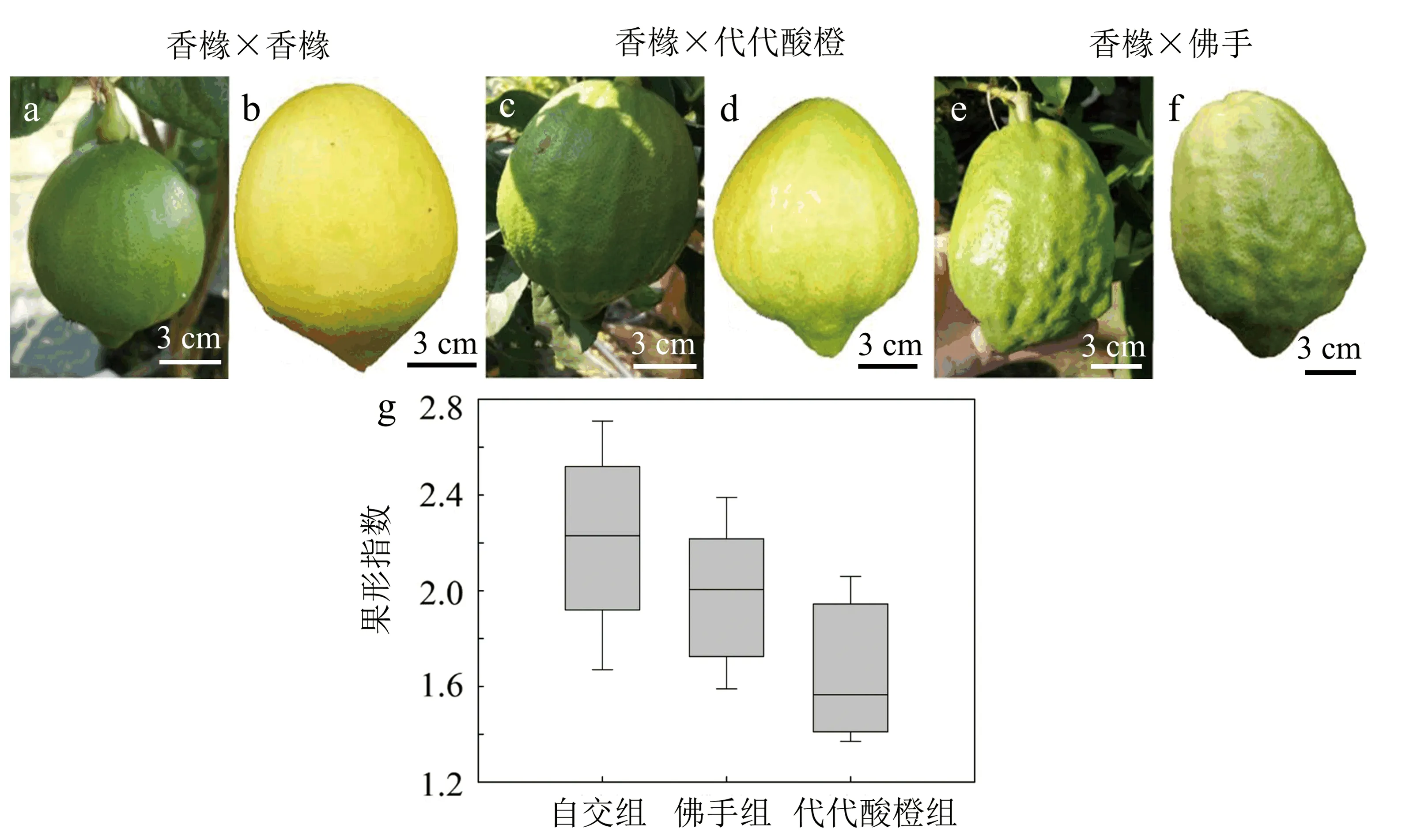
a:香櫞花粉授粉組未成熟香櫞;b:香櫞花粉授粉組成熟香櫞;c:代代酸橙花粉授粉組未成熟香櫞;d:代代酸橙花粉授粉組成熟香櫞;e:佛手花粉授粉組未成熟香櫞;f:佛手花粉授粉組成熟香櫞;g:授粉后的果形指數比較
2.3 果形相關基因在花粉和授粉前后柱頭上的表達分析
在授粉前,對果形相關基因在3種花粉中的相對表達量進行分析(見圖3)后發現,CNR基因在香櫞花粉中的表達量遠高于其他2種花粉,在佛手花粉中表達最低.CRC和YABBY5基因在佛手花粉中表達量最高,在香櫞和代代酸橙花粉中僅有微量表達;SUN基因在代代酸橙花粉中的表達量極低,在香櫞和佛手花粉中表達量稍高,但表達總量仍處較低水平.由于YABBY2僅在香櫞花粉中檢出,在佛手和代代酸橙中無法檢出,因此,對YABBY2基因的相對表達量分析未完成.KNA2在3種花粉中表達量都較低,無法分析.
觀察各基因在不同花粉授粉的雌蕊中相對表達量(見圖4)發現,CNR基因授粉后的表達量在自交組呈上調趨勢,在佛手和代代酸橙授粉后并未上調,尤其在代代酸橙組下調更明顯.CRC基因在佛手授粉當天的表達量有突增現象,各組都在授粉后表達顯著下調.SUN基因在自交組和佛手授粉組中授粉當天顯著上調,自交組授粉后相對授粉當天下調,但高于授粉前;SUN基因在代代授粉后表達量無顯著差異.KAN2基因授粉后的表達量在佛手組變化不顯著,在自交組出現上調趨勢,而在代代酸橙組出現下調趨勢.3個授粉組中的YABBY2基因表達趨勢總體相同,授粉后表達量都呈上調趨勢.YABBY5基因的表達在3個授粉組中均為授粉后下調,且低于授粉前;在代代酸橙授粉組中,在授粉當天顯著增高,其余2組并未有顯著差異.
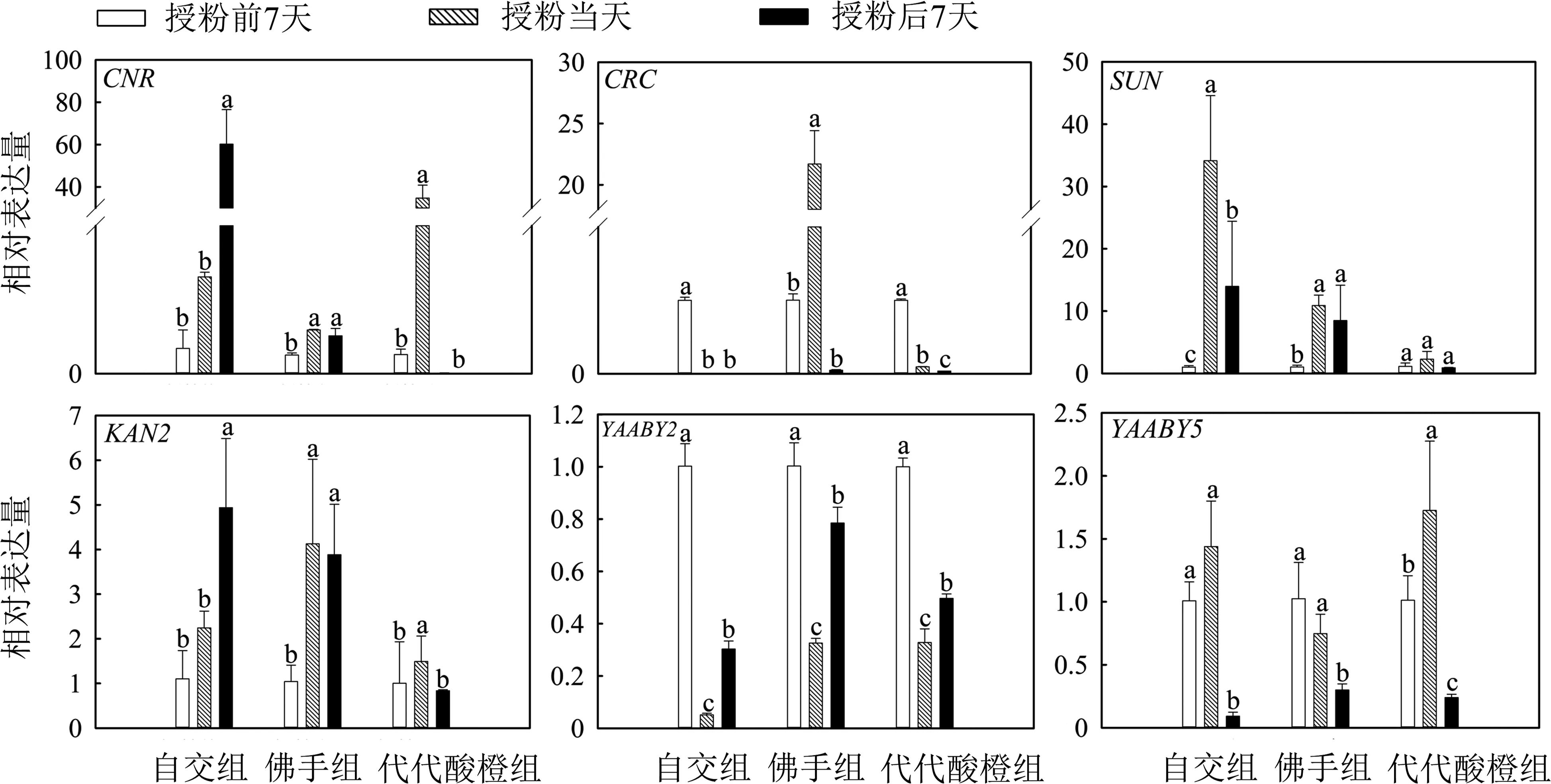
圖4 果形相關基因在不同花粉授粉的香櫞雌蕊相對表達量
3 討論與結論
為了探討花粉直感是否在柑橘屬作物果形建成中發揮作用,對不同花粉授粉后的香櫞果實形態進行了研究.通過比較自交組香櫞花芽不同發育時期后發現,香櫞雌蕊授粉前的發育明顯有一個縱向快速生長階段,而開花授粉后不同外源花粉的介入會影響花芽中這些基因的表達,這可能是不同花粉授粉導致香櫞果形變化的內在分子機理之一.不同外源花粉介入后可能會改變香櫞橫縱軸發育的起始時間點,從而在一定程度上改變果形.各組授粉后的香櫞果實在橫向上出現快速增長,導致果形指數發生變化.佛手花粉能使香櫞果形在橫軸上有明顯的縱裂凸起,使香櫞原本的長圓形果形性狀出現畸變;而代代酸橙花粉會使香櫞果形變得扁平,同時也能輕微加深表皮凹痕.由此可見,花粉直感確實能改變香櫞果形.根據果形相關基因在花芽與花粉的mRNA表達模式分析,可總結出結果,見表2及圖5.
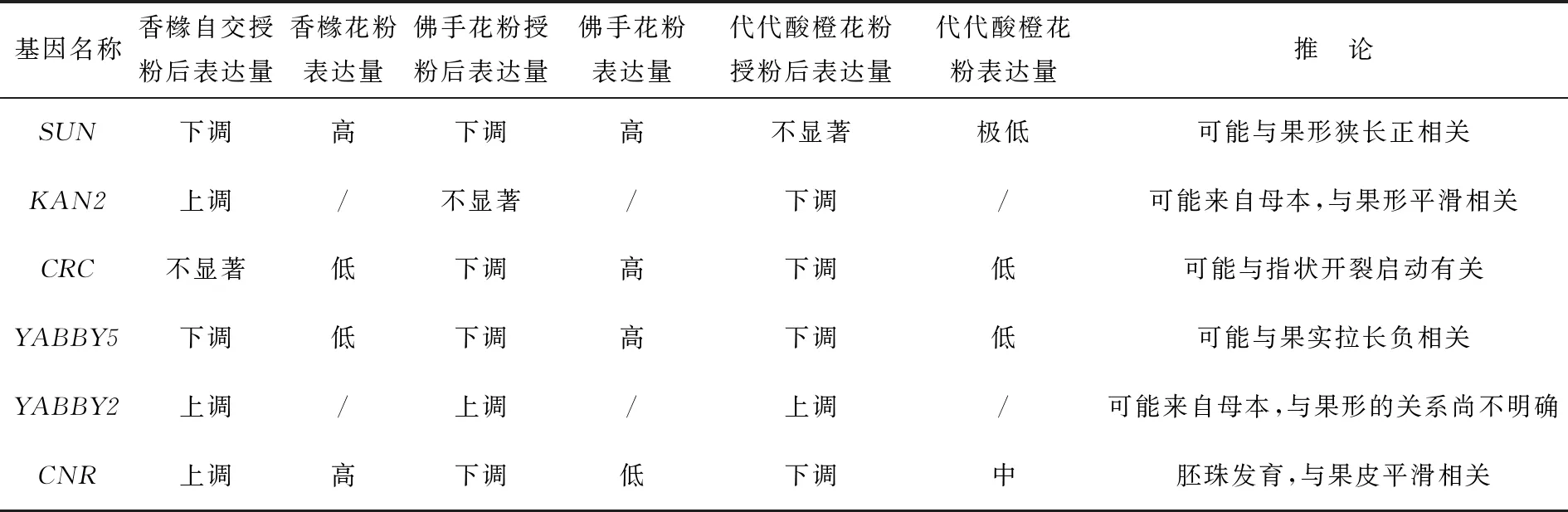
表2 果形相關基因在花芽與花粉的mRNA表達模式分析
已有研究表明,CmsSUN20在佛手和香櫞中的表達有顯著差異,可能是導致佛手果實長度大于寬度的原因[18].SUN基因在佛手、香櫞花粉中的表達量高于代代酸橙,而佛手、香櫞花粉授粉當天SUN表達量上調,代代酸橙授粉后并未有顯著性改變,即花粉中的SUN影響了授粉后的花柱;結合果形特征分析,SUN基因可能與果形狹長呈正相關.KAN2是心皮發育相關基因[12],本實驗在花粉中未檢測到KAN2基因的表達,推測KAN2基因可能來自母本;KAN2在自交組的3個時期表達量一直呈上升趨勢,但在佛手和代代酸橙授粉組中分別呈現授粉后無顯著差異或者下調趨勢,這可能導致佛手和代代酸橙組授粉的果實表皮特征不同.KAN2基因的表達可能與香櫞果實表皮平整相關.本實驗中CRC在佛手花粉中表達量遠遠高于其他2個物種,CRC在佛手授粉后確實引起了表達量的突增,即CRC可能與果形指狀開裂有關.YABBY5在佛手花粉中表達量也高于其他2種花粉,但是香櫞、佛手花粉的授粉都未顯著改變YABBY5在授粉當天雌蕊中的表達,僅在代代酸橙授粉當天表達上調,授粉后表達又顯著低于授粉前,推測YABBY5可能與果形拉長負相關.YABBY2在除香櫞外的花粉中未檢測到表達,卻在3組授粉當天顯著下降,授粉后表達上調.上述結果暗示,YABBY2與授粉的花粉種類無關,推測該基因可能來自母本,且與果形的發育關系尚不明確,可能與香櫞果形平滑相關.有報道發現,CNR基因在胚珠中表達[23].本實驗香櫞自交授粉后的CNR表達顯著上調,但佛手與代代酸橙花粉在授粉后并未顯著改變CNR的表達,且在代代酸橙授粉后表達顯著下調;結合佛手胚珠敗育[24]、代代酸橙種子敗育等性狀,本實驗驗證了CNR與胚珠發育相關,推測其與果皮平滑相關.
以果形發育的形態觀察為基礎,根據佛手、香櫞、代代酸橙的果實特點設置6個果形性狀:香櫞設紡錘形與表皮平整;代代酸橙設扁平形與表皮皺紋;佛手設狹長型指狀開裂與表皮縱裂.這些特征可歸納為果皮特征和形態特征(見圖5).同時,總結上述果形基因表達的模式分析(見表2),筆者確定了CRC,SUN,KAN2,CNR,YABBY5這5個基因在香櫞果形發育中發揮的可能作用(見圖5).與對照相比,佛手花粉授粉后香櫞果形的橫軸上出現明顯的縱裂凸起,凸起位置在果皮上隨機分布;代代酸橙花粉授粉后香櫞果形變扁平,并加深了表皮凹痕,這些形態特征可能由SUN,CRC,PbCNR6共同調控,而果皮的光滑或開裂由KAN2,CNR共同調控.SUN,YABBY5基因與果形拉長相關;CRC基因表達上調可能與指狀開裂啟動有關;推測KAN2可能與果實表皮平整相關;CNR基因與胚珠發育有關.但本次實驗尚不明確YABBY2基因與果形的關系,需要繼續開展類似實驗來對其進行探究.挖掘新的果形建成基因,并建立這些基因之間的關系,是未來果實形態建成的研究趨勢之一[25].后續研究將會擴大實驗樣本規模和基因數量,繼續發掘新的果形相關基因,并嘗試尋找不同果形相關基因之間的可能關聯.
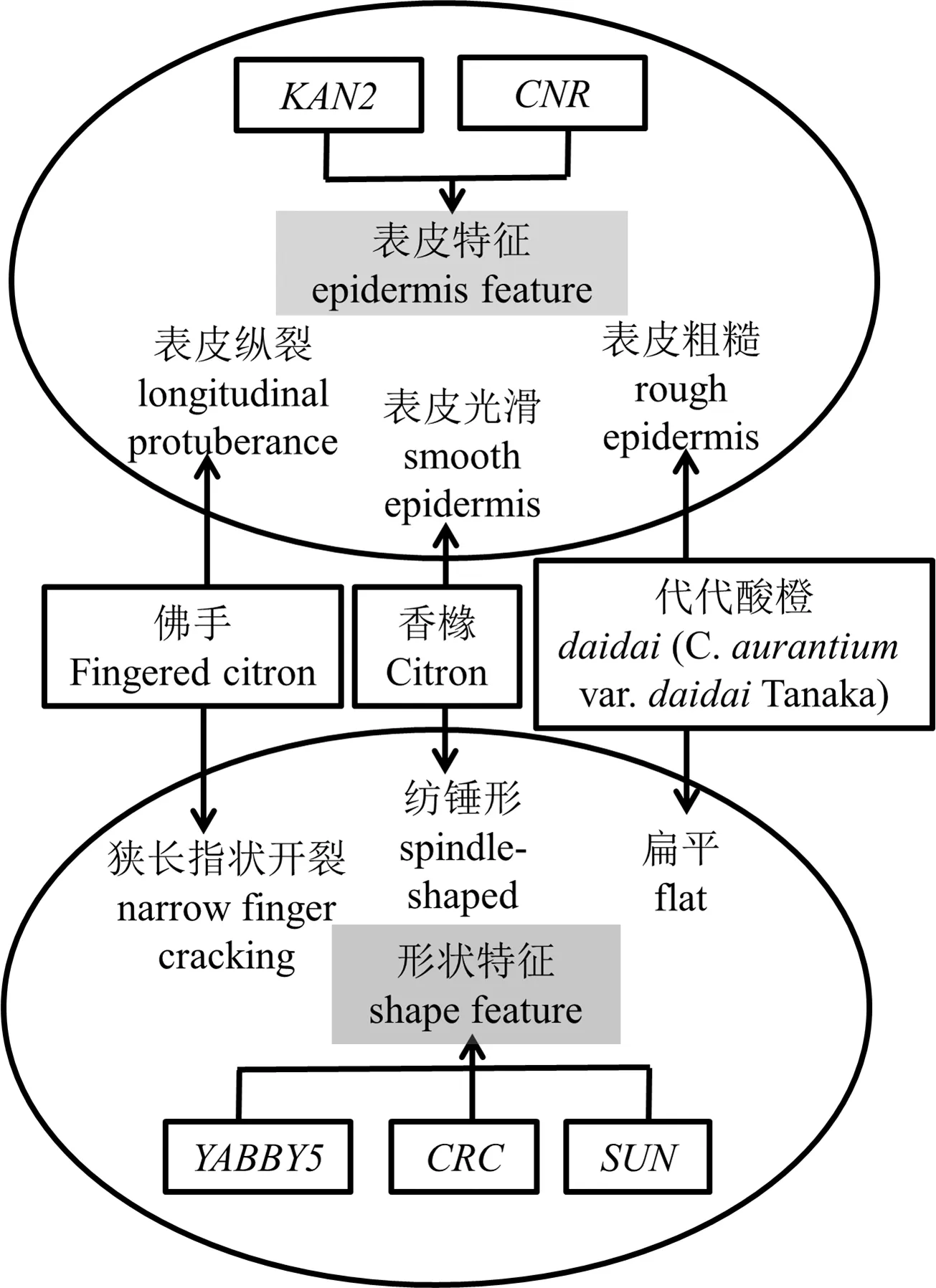
圖5 果形特征和基因關聯圖
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
