育成期高能飼喂下開產與未開產蛋雞肝臟轉錄組差異分析
2022-05-18 09:12:50王星果竇套存胡玉萍李永峰王克華
西北農業學報 2022年1期
曲 亮,王星果,郭 軍,竇套存,胡玉萍,馬 猛,李永峰,王克華
(江蘇省家禽科學研究所,江蘇揚州 225125)
蛋雞的生產性能直接與其養殖效益掛鉤,一直是蛋雞行業的重要研究課題[1],而能量水平對生產性能的影響是其中一個重要的研究方向,也越來越受到人們的重視[2-5],其中包括對育成期蛋雞進行能量調控,檢測其對生產性能的影響。研究表明育成期高能飼喂能明顯提高蛋雞的生產性能,如使用不同能量的飼料飼喂育成蛋雞的研究中發現,高能飼喂能顯著提高體質量、體尺、產蛋率和屠宰性能[6-10]。江蘇省家禽科學研究所蛋雞研究室(本研究室)前期研究了不同能量水平飼料對育成期蛋雞的影響,發現能量水平越高,蛋雞體質量越大,開產日齡越早[11]。
肝臟是重要的代謝器官,在雞等家禽中更是物質代謝特別是脂質代謝的主要器官,對育成期的能量調控肯定起著關鍵作用[12]。同時,產蛋與肝臟也有十分密切的關系[13]。由于雞蛋中卵黃的30%由脂質構成,而這些脂質均來源于肝臟。肝臟中合成的大部分脂質以脂蛋白的形式運輸出肝臟,經血液循環至卵泡,被卵母細胞吸收儲存,形成卵黃[14]。因此,肝臟脂質代謝的快慢影響雞蛋卵黃形成的速度,從而影響產蛋。
近年來,轉錄組測序逐漸用于研究各種生物過程[15-16]。已有一些轉錄組測序在雞肝臟中的研究報道,如對盧氏綠殼蛋雞產蛋前期和高峰期的肝臟、雌激素處理蛋雞的肝臟進行轉錄組測序研究,均發現基因參與肝臟代謝[17-18],本研究室成員也運用轉錄組測序發現LNA-122、GH處理的雞肝臟細胞中若干基因參與代謝過程,尤其是脂質代謝[19-20]。本研究室后期對高能飼喂的育成期蛋雞進行肝臟轉錄組測序,發現與低能組相比,有188個基因差異表達,對差異表達基因的功能和信號通路分析發現,脂質代謝相關過程富集的基因較多,且顯著富集的信號通路都和脂質代謝相關[21]。然而在高能組中的雞仍有開產早晚之分,有必要對其進行深入分析。本研究選取高能飼喂育成期蛋雞,并對其中開產與未開產雞的肝臟進行轉錄組測序,分析比較其表達譜,探明在高能飲食狀態下肝臟中影響開產的基因,為提高產蛋打下基礎。
1 材料與方法
1.1 試驗動物飼養
選用江蘇省家禽科學研究所(本單位)培育的C3蛋雞品系,8周齡前常規飼養,滿8周齡時選取體質量相近母雞80只,分為8個重復,每個重復10只。9~18周齡,定量飼喂代謝能水平為12.14 MJ/kg的高能飼糧,18周齡后自由采食標準營養水平的飼糧。整個試驗過程參照NRC(1994)標準配制試驗日糧,分9~18周齡和19周齡后兩個階段配制,配以粉料飼喂。9~16周齡在育成雞舍四層階梯籠內飼養(5只/籠),17周齡起在產蛋雞舍三層階梯籠內飼養(1只/籠)。整個試驗期間,所有雞只可以自由飲水,并執行常規光照和免疫等程序。
1.2 RNA-seq高通量測序
以20周齡作為開產早晚的界限,開產雞均為19周齡開產,未開產雞均在20周齡末仍未開產,20周齡末每個重復選取1只開產雞和1只未開產雞,稱體質量后各取最接近平均值的3只,未開產雞分別為N1、N2、N3,開產雞分別為R1、R2、R3,屠宰并取出肝臟組織,放入液氮中保存。
使用TRIzol試劑提取肝臟組織總RNA,使用瓊脂糖電泳和Agilent 2100檢測RNA質量。利用rRNA去除試劑盒去除rRNA,并通過離子打斷的方式將RNA打斷成200~300 bp的片段。RNA經過反轉錄形成cDNA,再經PCR富集構建cDNA文庫,該文庫通過熒光定量檢測其大小和濃度,上機至Illumina Nextseq500平臺進行 測序。
1.3 序列分析
下機得到原始數據,經過reads數、Q20、Q30等初步統計,經堿基質量分布檢測后去除接頭,進行質量過濾,得到clean reads和clean data。這些數據使用Tophat2與雞參考基因組進行比對,統計比對到外顯子、內含子和基因間區的reads。比對結果使用RSeQC進行插入片段長度分布、reads重復率等質控分析。
比對到各基因的reads使用HTSeq統計,得到各基因的原始表達量,開產雞和未開產雞的表達量經過RPKM標準化后,再使用DESeq分析其差異性,按照表達量倍數差異(fold change)> 2、差異顯著性P<0.05篩選出差異表達基因。挑選上調基因和下調基因各2個,使用熒光定量PCR(qRT-PCR)進行表達量驗證,以ACTB作為內參,引物信息見表1。對差異表達基因使用超幾何分布進行GO功能富集分析和KEGG信號通路富集分析。

表1 qRT-PCR引物Table 1 Primers for real-time qRT-PCR
2 結果與分析
2.1 蛋雞肝臟的測序質量
未開產與開產蛋雞的肝臟經過RNA-seq測序,得到轉錄組原始序列。表2為測序數據的初步分析,可見,6個樣品測得的原始序列經質量過濾后,純凈序列仍超過80 000 000個,占原始序列的99%以上,說明低質量的序列少,測序效果較好。開產雞與未開產雞的原始序列和純凈序列數量均相當,說明兩組測序質量相近。
將各樣品的純凈序列與雞參考基因組比對,由統計比對上的序列(表 3)可見,6個樣品能比對到基因組的序列均占純凈序列的80%左右,且絕大部分是基因序列,約90%,外顯子序列在基因序列中所占比例也很高,均占80%左右。以上結果表明6個樣品測序結果均較好,得到的序列基本為有效的基因序列,可進行后續研究。
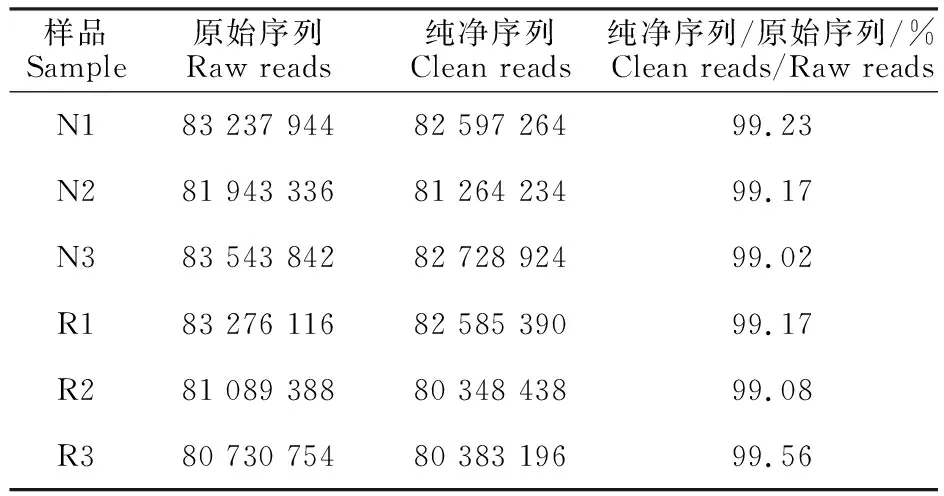
表2 測序數據初步分析Table 2 Preliminary analysis of RNA-seq data
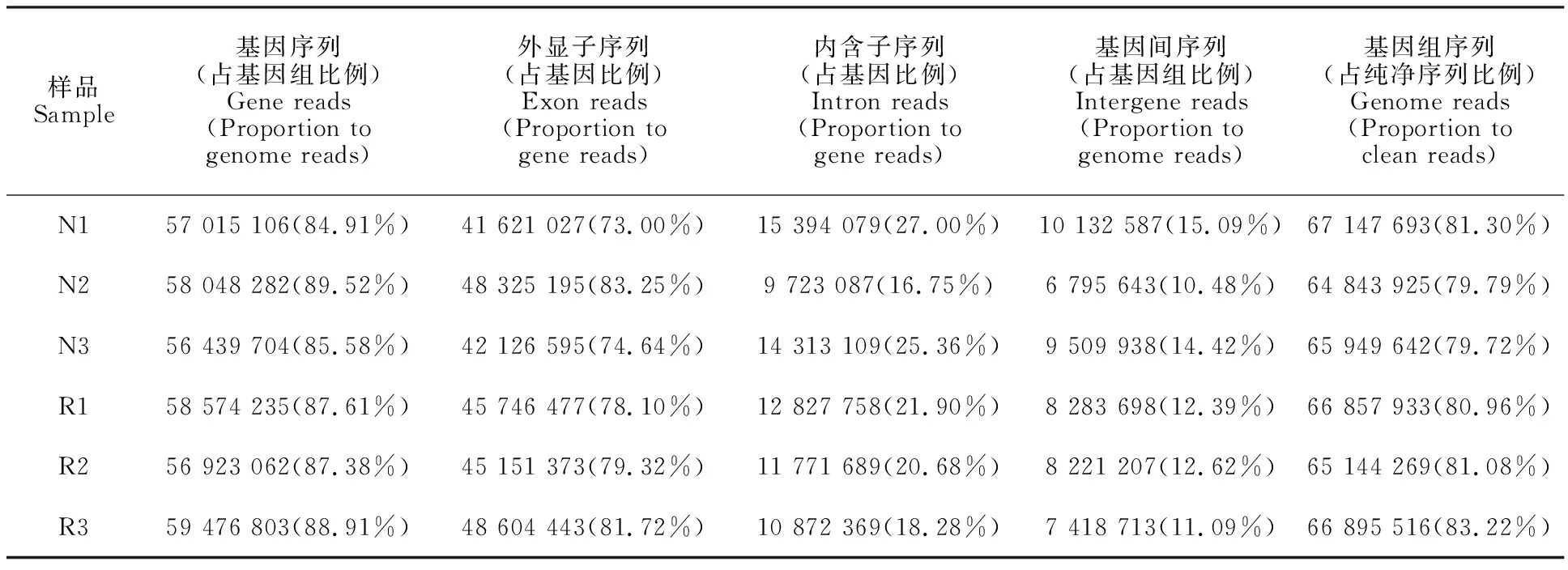
表3 基因鑒定Table 3 Identification of gene
2.2 肝臟基因差異表達分析
對鑒定出的基因進行RPKM表達量分析,并對開產與未開產樣品的基因表達量進行比較分析,篩選出差異表達基因(圖1),共篩選出494個差異表達基因(P<0.05),相比未開產雞,開產雞有285個基因上調,209個基因下調。對差異表達基因進行聚類分析(圖2),結果可見,基因大多匯聚在同一大類,說明它們具有相關的功能。表達差異最大的20個基因(10個上調基因,10個下調基因)見表4,可以看出,未開產雞有7個基因未檢出表達,開產雞有5個基因未檢出表達,上調和下調差異為10~3 000倍,提示它們可能對開產性狀具有特殊的重要作用。
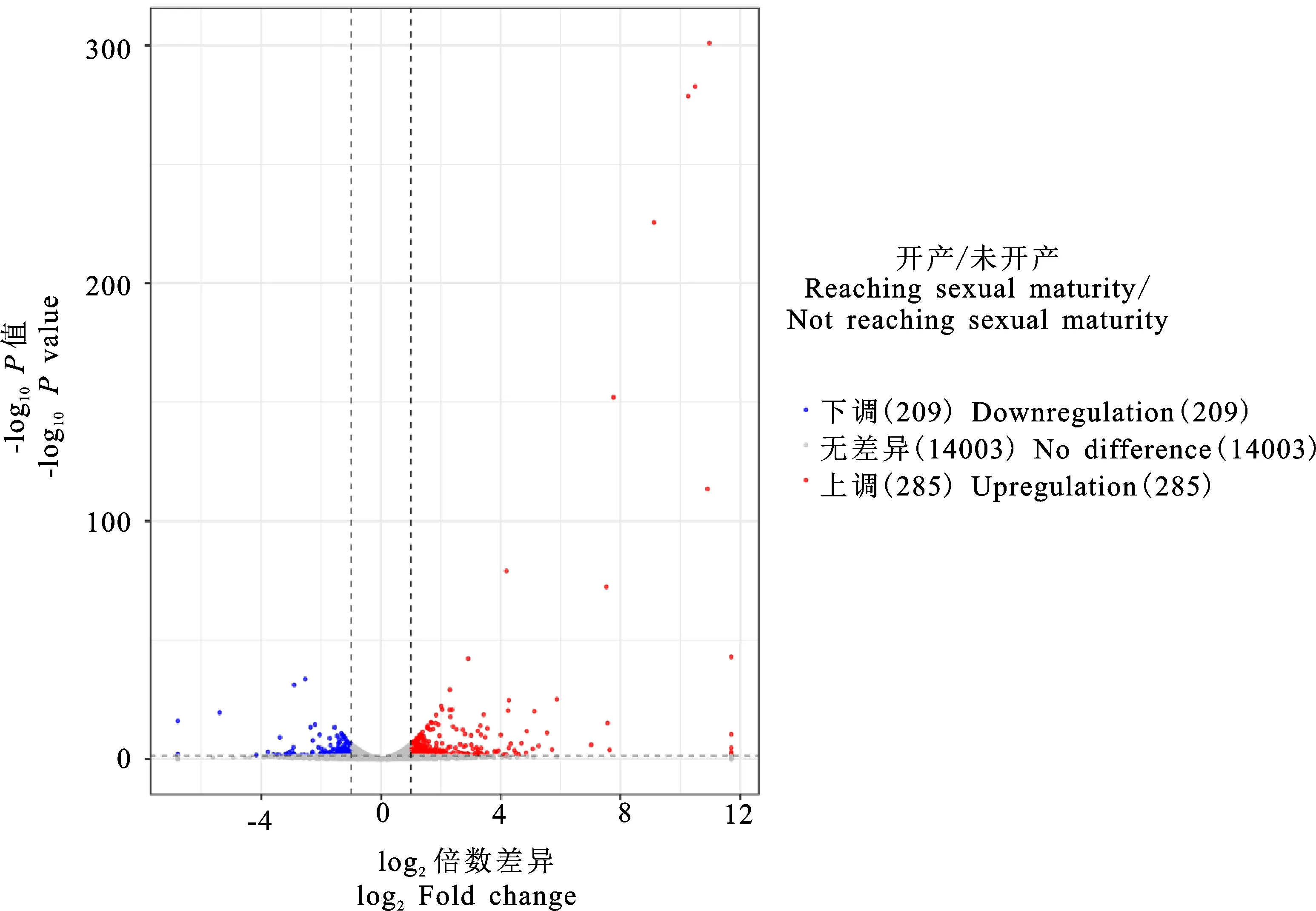
圖1 差異表達基因火山圖Fig.1 Volcano map of differentially expressed genes
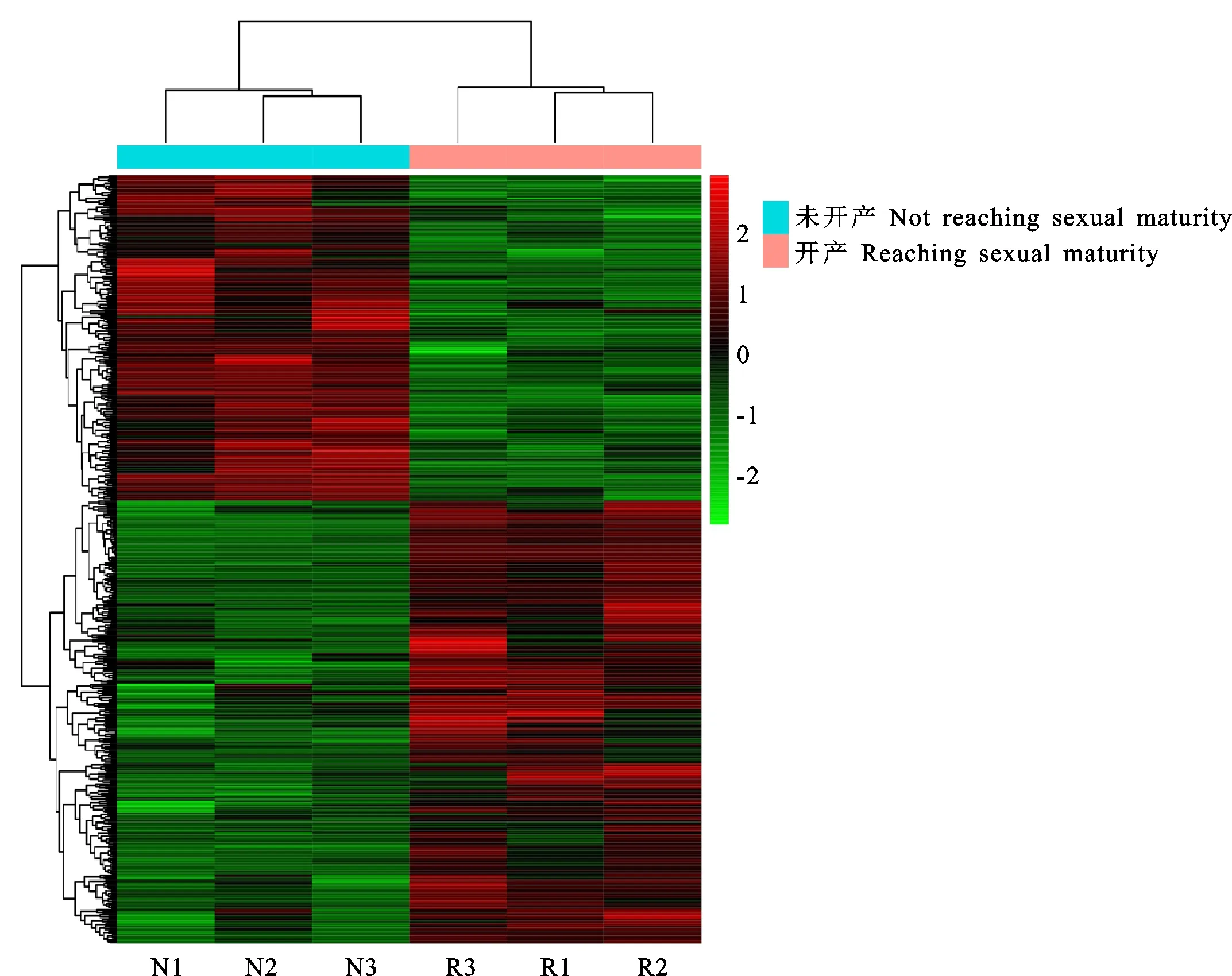
紅色為高表達,綠色為低表達 Red means high expression,green means low expression
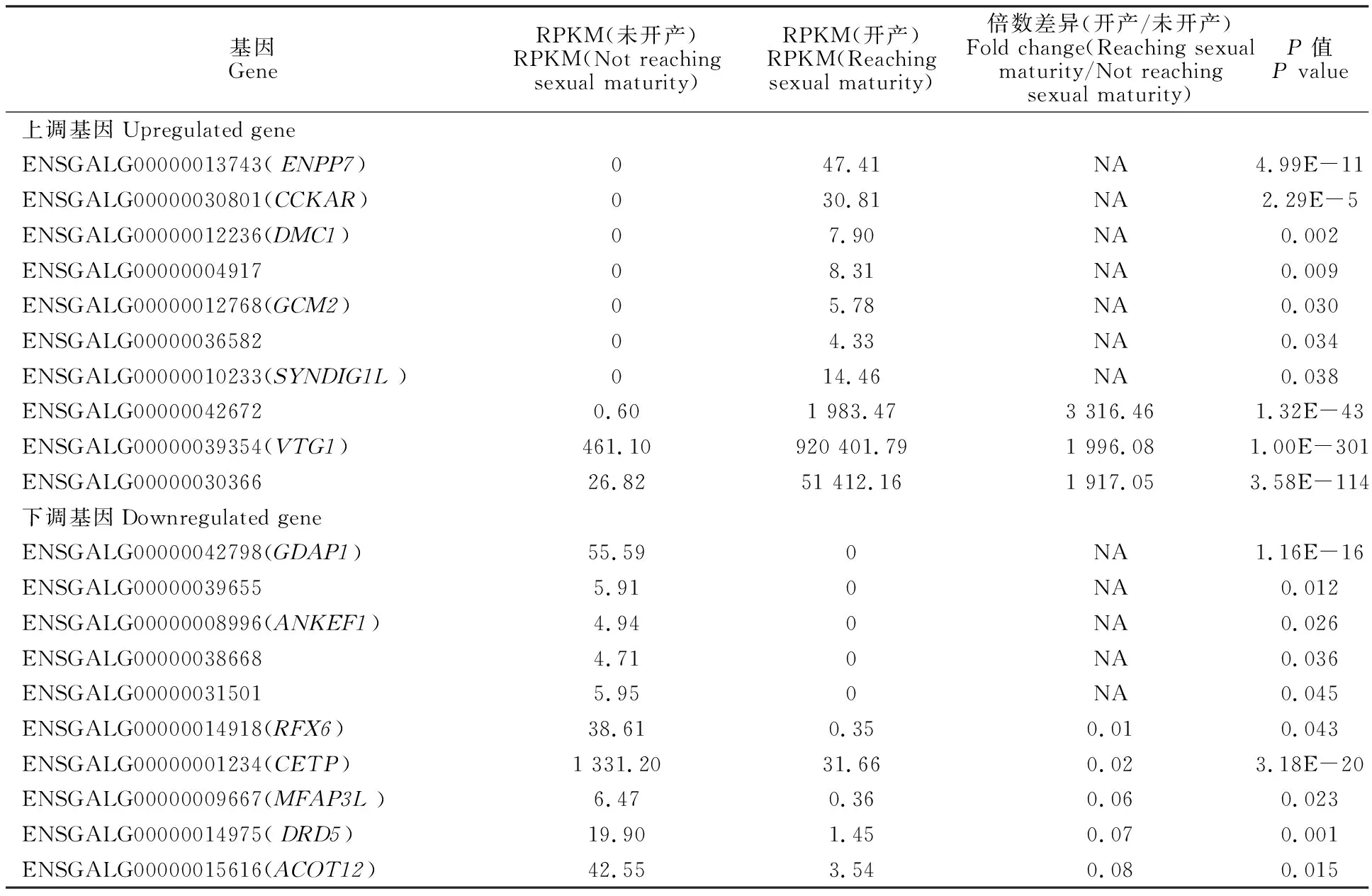
表4 表達差異最顯著的20個基因Table 4 Top 20 differentially expressed genes
挑選部分差異表達基因進行qRT-PCR驗證(圖3),結果可見,4個基因表達量的qRT-PCR結果與RNA-seq結果基本一致,說明差異表達基因分析結果較可靠。
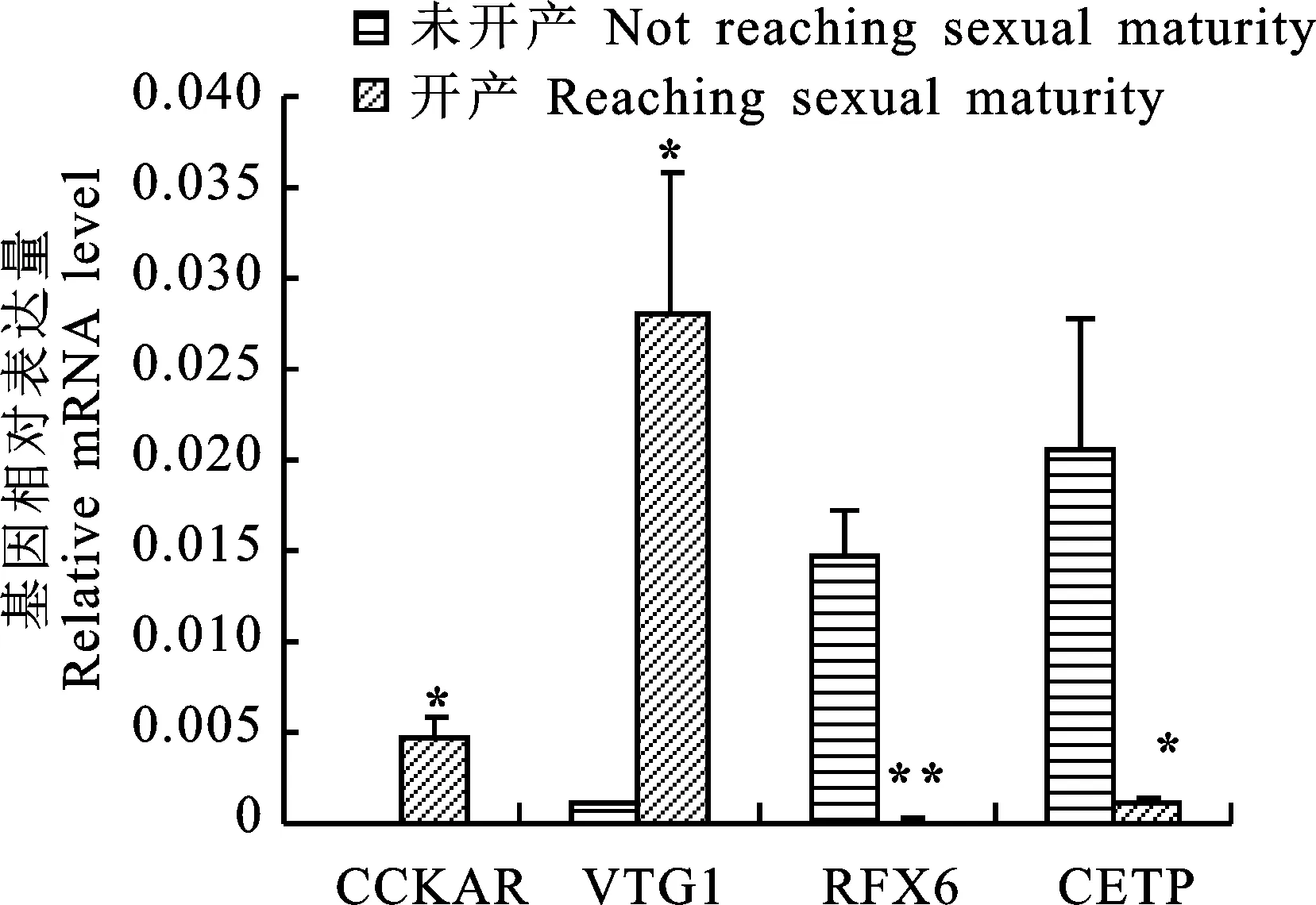
*.P<0.05; **.P<0.01
2.3 肝臟差異表達基因的功能分析
2.3.1 GO功能富集分析 對開產雞和未開產雞的差異表達基因進行GO功能富集分析,發現有60個顯著富集的GO功能(FDR<0.05),其中44個富集在生物過程,11個富集在細胞組分、5個富集在分子功能。圖4顯示富集最顯著的20個GO功能,除少數細胞組分的GO高度富集外,大部分均為脂質代謝相關功能,說明脂質代謝對開產很重要。這些GO功能富集的基因多則53個,少則5個。
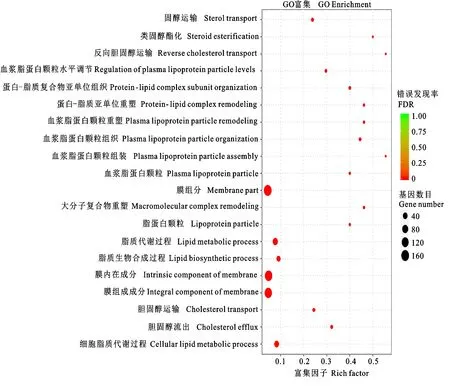
圖4 差異表達基因GO富集分析:富集最顯著的20個GO功能Fig.4 GO enrichment analysis of differentially expressed genes: top 20 enriched GO
2.3.2 KEGG信號通路富集分析 對開產雞和未開產雞的差異表達基因進行KEGG信號通路富集分析,發現僅有1個顯著富集的KEGG信號通路(FDR<0.05),即非不飽和脂肪酸生物合成(Biosynthesis of unsaturated fatty acids)(表5)。此為脂質代謝相關的信號通路,包含基因FADS1、FADS2、ACOT7、ACAA1和ELOVL6,說明非不飽和脂肪酸生物合成信號通路與開產有密切關系。

表5 差異表達基因KEGG富集分析Table 5 KEGG enrichment analysis of differentially expressed genes
3 討論與結論
高通量測序技術發展迅速,現在較常用的RNA-seq數據量一般都是千萬級別的原始序列[22-23],本研究RNA-seq所得原始序列就在千萬級別。同時,各組測序數據能比對上基因組的序列均在80%左右,而其中比對上基因的序列均超過80%,結果表明轉錄組測序所得數據質量較高,后期分析結果更準確,更可信。
對差異表達基因進行RT-qPCR驗證,證明轉錄組測序結果比較可靠。進一步分析后,發現在差異最大的20個基因中,有7個(ENPP7、CCKAR、DMC1、GCM2、SYNDIG1L、ENSGALG00000004917、ENSGALG00000036582)和 5個(GDAP1、ANKEF1、ENSGALG00000039655、ENSGALG00000038668、ENSGALG00000031501)基因分別在低能組和高能組未檢測出表達,ENPP7與腫瘤風險相關[24],CCKAR與生長和體質量相關[25],DMC1與減數分裂相關[26],GCM2與維持甲狀旁腺功能相關[27],SYNDIG1L與哺乳動物乳頭數相關[28],GDAP1與線粒體分裂等相關[29],其他基因尚無功能研究報道。這些基因在肝臟對能量攝入量的影響中雖然發揮了重要作用,但是機理尚不明確,需要進行更深入的研究。
對差異表達基因進行GO富集分析,發現與脂質代謝相關的功能在富集程度最高的20個功能中最多,共有16個GO,說明脂質代謝過程是對開產影響最大的生物過程。李云雷等[30]對北京油雞開產前和開產后腹脂與性發育的關系研究發現,開產前腹脂質量與卵巢質量和卵巢指數呈中等正相關,表明育成期性腺發育一定程度上受脂肪沉積的影響;開產后腹脂質量與卵巢質量、卵巢指數和輸卵管質量均呈中等程度負相關,且低腹脂率組的開產率高于高腹脂率組,其開產一致性較好,腹脂沉積過量導致北京油雞性成熟和開產延遲。對盧氏雞青年雞及產蛋雞肝臟轉錄組的研究發現,雞肝臟脂質轉運相關基因MTTPL影響蛋雞性成熟[31]。另外,對白來航雞產蛋前后的肝臟轉錄組測序研究也發現RNF186等與脂質代謝相關的差異表達基因,提示它們影響蛋雞的產蛋性能[32],這些研究均表明脂質代謝及其相關基因對蛋雞產蛋有重要影響。
對差異表達基因進行KEGG富集分析,發現唯一顯著富集的通路非不飽和脂肪酸生物合成也與脂質代謝相關,通路中差異表達基因FADS1、FADS2是一類脂肪酸脫氫酶基因,參與多種多不飽和脂肪酸的合成,表達量與脂肪酸組成顯著相關[33-34]。ACOT7是脂酰輔酶A硫脂酶7,能將脂酰輔酶A水解成游離脂肪酸,參與調節游離脂肪酸含量[35]。ACAA1是乙酰輔酶A酰基轉移酶1,催化β-氧化途徑的最后一步,參與脂肪酸的延伸和降解[36]。ELOVL6是長連脂肪酸延伸酶6,能催化延長C12-C16的飽和或單不飽和脂肪酸,調節脂肪酸組成總體平衡[37]。而李慧鋒等[32]對高產雞產蛋前后肝臟差異表達基因進行KEGG分析,發現富集的通路及相關基因主要與脂質代謝和氨基酸代謝相關,由此可見,脂質代謝相關信號通路確實影響蛋雞開產。
綜上所述,育成期高能飼喂的蛋雞中,與肝臟脂質代謝相關的一些基因差異表達會影響肝脂代謝,最終影響開產時間。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
