碎米薺超氧化物歧化酶基因家族成員的鑒定和分析
2022-06-10 10:30:24付偉李林康宇龍瀾楊永康殷紅清向極釬梅磊
熱帶亞熱帶植物學報 2022年3期
付偉, 李林, 康宇, 龍瀾, 楊永康, 殷紅清*, 向極釬,梅磊
(1. 恩施土家族苗族自治州農業科學院,湖北 恩施 445000;2. 湖北省富硒產業技術研究院,湖北 恩施 445000;3. 中國科學院植物研究所,北京 100093)
活性氧自由基(reactive oxygen species, ROS)是植物細胞在需氧代謝過程中產生的具有高度化學反應活性的含氧集團,如超氧陰離子自由基(O2-·)、羥基自由基(·OH)、過氧化氫(H2O2)、單線態氧(1O2)等。當植物遭受脅迫時,抗氧化酶系統受到損傷導致活性氧大量積累,引起一系列生理生化紊亂對植物造成損害,甚至導致不可修復性的代謝功能障礙以及細胞死亡[1]。在長期的進化過程中,植物為了抵御氧化傷害,維持正常的生理代謝,形成了抗氧化酶促清除系統和非酶促清除系統協同作用清除活性氧。而超氧化物歧化酶(superoxide dismutase,SOD)作為植物防御系統的第一道防線對清除氧自由基起關鍵作用,可催化超氧陰離子自由基(O2-·)歧化生成H2O2和O2,再通過過氧化物酶和氧化物酶將H2O2轉化為H2O,從而實現活性氧的清除[2],其活性與植物的生長發育以及抗逆性等有密切相關性[3-5]。
超氧化物歧化酶是一種含金屬的酶,其通過金屬輔因子得失電子清除活性氧自由基。根據SOD 結合的金屬輔因子種類,可分為Cu/Zn-SOD、Fe-SOD、Mn-SOD 和Ni-SOD 等4 類[5-6]。其中,Cu/Zn-SOD主要存在于高等植物的葉綠體和細胞質中,與植物抗逆性有密切關系[7]。在小麥(Triticum aestivum)中,組成型表達的Cu/Zn-SOD響應各種非生物脅迫的程度不同[8]。低溫脅迫下,茶樹(Camellia sinensis)的Cu/Zn-SOD和Mn-SOD基因上調表達,Fe-SOD基因受到抑制[9]。轉基因植株中,Cu/Zn-SOD基因的過表達增強了木薯(Manihot esculenta)[10]耐旱性和甘薯(Dioscorea esculenta)[11]的鹽脅迫耐受性。而Fe-SOD 和Mn-SOD 主要分布于低等植物中,主要涉及植物的抗逆性[12]。在煙草(Nicotiana tabacum)與苜蓿(Medicago sativa)中,Fe-SOD和Mn-SOD明顯增強了植株的抗冷、抗氧化能力[13-14]。Ni-SOD 首次從鏈霉菌(Streptomyces)中發現,目前報道其僅存在于少數原核生物如綠藻和細菌中[15]。Tsang 等[16]通過亞細胞和系統發育分析表明,僅Cu/Zn-SOD、Fe-SOD和Mn-SOD 在植物中共存。
碎米薺(Cardamine hirsuta)為十字花科(Brassicaceae)碎米薺屬碎米薺組植物,分布廣泛,全株可作野菜食用和入藥[17]。有研究表明,碎米薺組有些植物具有較強的硒耐受性,如彎曲碎米薺(C. flexuosa)[18]和堇葉碎米薺(C. violifolia)[17]等。碎米薺全基因組測序的完成,為ChSOD基因家族的全基因組分析奠定了基礎。然而,目前尚未有從全基因組水平上對碎米薺SOD基因功能進行研究的報道。本研究通過生物信息學對碎米薺SOD基因家族進行全基因組鑒定分析,分析其結構、進化關系和染色體定位等信息,為碎米薺抗氧化機制研究提供理論參考。
1 材料和方法
1.1 材料
選取顆粒飽滿的碎米薺種子于4 ℃冰箱春化24 h 后,置于培養皿中黑暗催芽4 d。待種子萌發后,將其移至塑料花盆中(蛭石∶營養土=1∶1), 并在光周期為12 h/12 h、25 ℃下培養15 d,分別收集碎米薺幼苗的根、莖和葉,用液氮快速冷凍后保存于-80 ℃冰箱備用。
1.2 SOD 基因家族成員鑒定
通過數據庫(http://chi.mpipz.mpg.de/assembly.html)下載碎米薺全基因組數據[19],通過Pfam 數據庫(http://pfam.xfam.org/)下載SOD 的隱馬氏模型文件(PF00080、PF00081 和PF02777),并以該文件為搜索條件,利用HMMER 3.0 中的Hmmsearch 程序對碎米薺蛋白質序列進行搜索(E<1×10-5),并去除重復,然后通過NCBI-CDD (http://www.ncbi.nlm.nih.g/stuctu/cdd/wrpsb.cgi)和SMART (http://smart.embl heidelberg.de/)進行鑒定和篩選,最終得到碎米薺SOD 家族蛋白質序列。根據碎米薺基因組注釋文件信息,獲得SOD基因在染色體上的位置分布信息,并利用Map Chart 繪制碎米薺SOD基因在染色體上的定位圖。最后,利用ExPASy (https://web.expasy.org/compute_pi/)分析碎米薺SOD 的分子量、氨基酸數量和等電點。
1.3 系統發育關系和分類
從TAIR 數據庫(https://www.arabidopsis.org/index.jsp)和RGAP 數據庫(http://rice.plantbiology.msu.edu/index.shtml)分別下載擬南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的SOD 氨基酸序列,利用Clustal W 對碎米薺、擬南芥和水稻的SOD 氨基酸序列進行比對,并用MEGA 7.0 軟件通過相鄰連接法(NJ)構建系統進化樹,設置重復次數為1 000,提高進化分支樹結果的可靠性。
1.4 基因結構和基序組成
利用在線網站GSDS (http://gsds.cbi.pku.edu.cn/)分析碎米薺SOD基因家族成員的基因結構;利用在線軟件MEME (http://memesuite.org/)分析碎米薺SOD 氨基酸序列中的保守基序,設置motif 為10;利用Clustal W 對碎米薺SOD 氨基酸序列進行比對,并用MEGA 7.0軟件相鄰接法(NJ)構建系統進化樹,設置重復次數為1 000。最后利用TBtools 繪制碎米薺SOD基因家族成員的系統進化樹、基因組成和基因結構的組合圖。
1.5 順式作用元件分析
提取碎米薺SOD基因編碼序列上游1.5 kb 文件,提交到Plant CARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)對順式作用元件進行識別,并利用在線網站GSDS (http://gsds.cbi.pku.edu.cn/)繪制碎米薺SOD基因家族順式作用元件圖[20]。
1.6 SOD 基因表達分析
取碎米薺根、莖、葉冷凍樣本各50 mg,采用Trizol 法提取總RNA,采用0.8%的瓊脂糖凝膠電泳和Nanodrop 2000 檢測RNA 質量,以確保總RNA 的完整性和濃度。參照SuperscriptⅢ first strand cDNA synthesis Kit (天根生化科技有限公司,北京)說明書進行cDNA 第一鏈合成,置于-20 ℃保存備用。
利用Primer Premier 3.0 進行熒光定量PCR 引物設計(表1),引物由睿博興科生物技術有限公司廣州分公司合成。以GADPH為內參基因,采用Super-Real PreMix Plus (SYBR Green) PCR Kit (天根生化科技有限公司,北京)進行實時熒光定量PCR。擴增體系(20μL):2×SuperReal PreMix Plus 10μL,上下游引物(10μmol/L)各1μL,cDNA 1μL,ddH2O 7μL。反應程序:95 ℃預變性10 min;然后95 ℃ 10 s,60 ℃ 20 s,72 ℃ 30 s,共45 個循環。采用2-ΔΔCT法進行數據分析,并用Graph Pad Prism 8繪制柱狀圖。

表1 qRT-PCR 引物Table 1 qRT-PCR primers
1.7 SOD 的二級、三級結構預測
利用在線網站SOMPA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL (https://swissmodel.expasy.org/)分別對碎米薺SOD 蛋白序列進行二級、三級結構預測。
2 結果和分析
2.1 碎米薺SOD 家族成員
通過HMMER3.0 中的Hmmsearch 程序對碎米薺氨基酸序列進行搜索,同時利用NCBI-CDD 和SMART 進行蛋白結構域鑒定,剔除了不含保守結構域的氨基酸序列,并手動去除重復的氨基酸序列和不完整的氨基酸序列,最終鑒定出10 個碎米薺SOD基因家族成員,不均勻地分布在7 條染色體上,其中在1、4 和6 號染色體上各有2 個,而3、5、7和8 號染色體上各有1 個(圖1)。

圖1 碎米薺SOD 基因的染色體分布Fig. 1 Chromosome location of ChSOD genes
對碎米薺SOD 家族成員進行氨基酸序列和結構分析表明(表2),有4 個ChSOD(CARHR008750、CARHR121080、CARHR085500 和CARHR256990)分布在染色體正義鏈上,其余6 個分布在反義鏈上;氨基酸序列長度為57 (CARHR120080)~324 (CARH R012800);蛋白質分子量為6 419.41~34 659.01 kDa;等電點在4.92~9.60,且酸性蛋白較多。

表2 碎米薺SOD 基因及其編碼蛋白信息Table 2 ChSOD genes and information of their encoding proteins
2.2 系統發育分析
通過Clustal W 對碎米薺10 個SOD 的氨基酸序列進行比對,結果表明(圖2),SOD 的氨基酸序列間同源性較高,且主要集中在C 端,該區域可能是SOD 蛋白行使功能的關鍵區域。
用MEGA 7.0 對碎米薺、擬南芥和水稻SOD 家族成員的系統進化關系進行了比較,并構建了系統發育進化樹。結果表明(圖3),相較于水稻,碎米薺與擬南芥的SOD 蛋白質具有相似的結構而聚在一起,說明碎米薺與擬南芥同源性較高,可通過分析擬南芥相應同源序列來推測碎米薺基因的功能;此外,進化樹還表明,Cu/Zn-SOD 起源于同一分支, 而Mn-SOD、Fe-SOD 起源于另一分支,表明碎米薺Mn-SOD 和Fe-SOD 也可能起源于同一祖先。
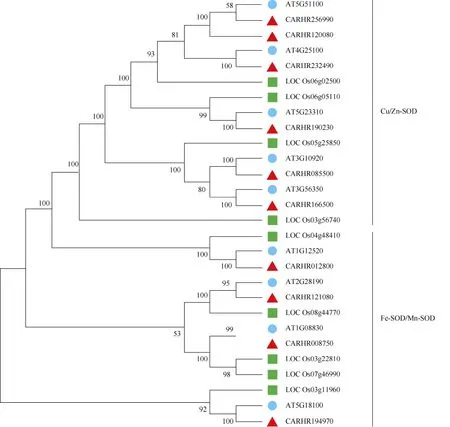
圖3 碎米薺(▲)、擬南芥(●)和水稻(■)中SOD 蛋白的系統進化樹Fig. 3 Phylogenetic tree of SOD in Cardamine hirsuta (▲), Arabidopsis thaliana (●) and Oryza sativa (■)
2.3 SOD 基因結構和基序組成
基因結構分析表明,ChSODs含有2~9個外顯子,其中CARHR256990 含有9 個外顯子,而CARHR 120080 僅有2 個外顯子(圖4)。而對ChSOD 進行保守基序分析表明,同一亞族內的SOD 保守基序的種類與數量大體相同,而不同亞族內的SOD 保守基序的種類和數量差異較大,Motif 3、Motif 5 僅出現在Fe-SOD/Mn-SOD (CARHR012800、CARHR194970、CARHR008750 和CARHR121080)分支中,而其余8種Motif 只存在于Cu/Zn-SOD (CARHR256990、CAR HR120080、CARHR190230、CARHR232490、CARHR 085500 和CARHR166500)分支中。

圖4 碎米薺SOD 家族成員的系統發育關系(A)、基序(B)和基因結構(C)Fig. 4 Phylogenetic tree (A), motiff (B) and gene structure (C) of ChSOD members
2.4 ChSODs 啟動子順式作用元件分析
為了進一步研究ChSODs 在非生物脅迫響應中的潛在機制,提取ChSODs 起始位點上游1.5 kb 序列,提交到Plant CAR 檢測順式作用元件,包括ABA響應元件、干旱誘導響應元件(MBS)、低溫響應元件(LTR)、防御和應激反應元件(TC-rich)和創傷響應元件(WUN-motif)。從圖5 可見,ChSODs 含有2~11個順式作用元件,其中CARHR008750、CARHR01 2800 和CARHR194970 僅有2 個順式作用元件,而CARHR166500 有11 個順式作用元件。此外,有6個ChSODs 基因具有ABA 響應元件,5 個基因具有LTR 響應元件,5 個基因具有TC-rich 響應元件,4個基因具有WUN-motif 響應元件和4 個基因具有MBS 響應元件。因此,推測在響應非生物脅迫時,ChSODs 對ABA 和低溫脅迫響應更敏感[20]。
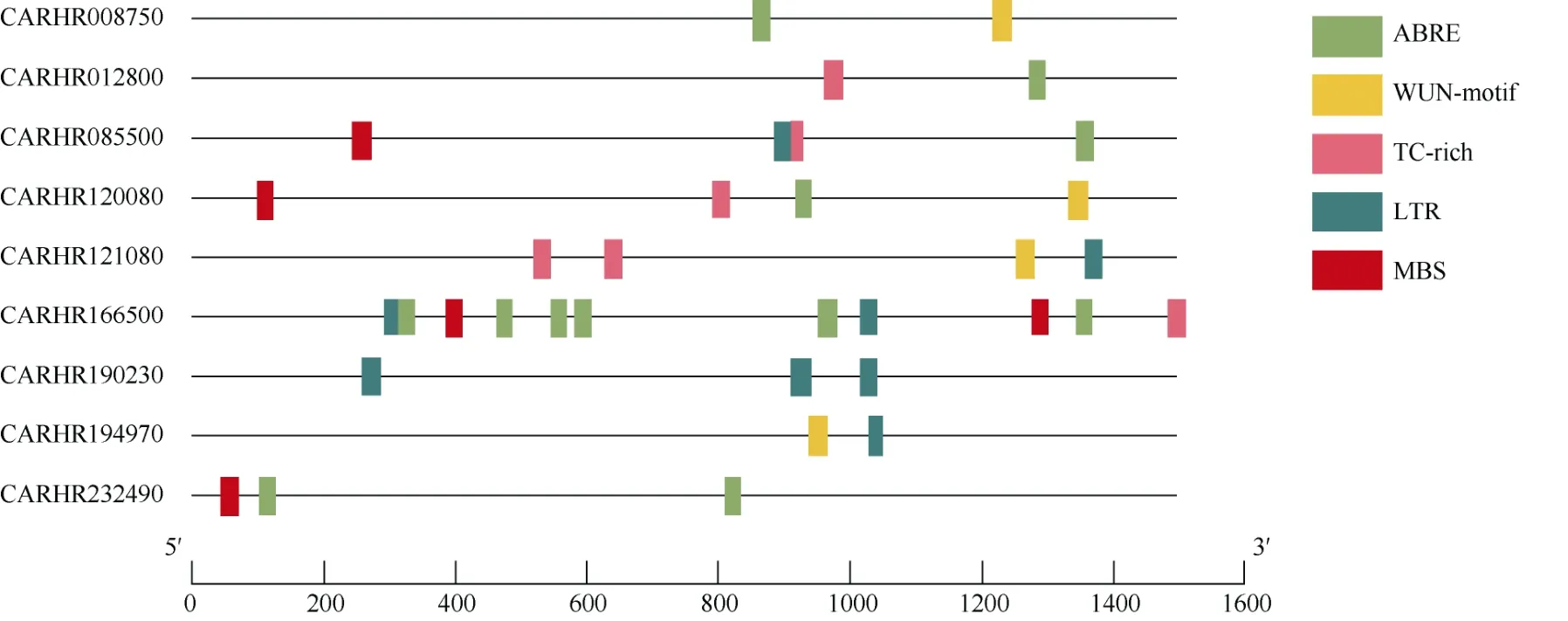
圖5 預測ChSOD 啟動子中的順式元件。ABRE: ABA; LTR: 低溫; MBS: 干旱; TC-rich: 應激反應; WUN-motif: 創傷。Fig. 5 Predicted cis-elements in ChSOD promoters. ABRE: ABA; LTR: Low temperature; MBS: Drought; TC-rich: Stress; Wun-motif: Trauma.
2.5 ChSODs 表達分析
基因的組織表達與功能密切相關,從圖6 可見,ChSODs 在碎米薺根、莖和葉中均有表達,以葉中的表達量最高,其次為莖和根。另外,CARHR08 5500 在葉和莖中的表達量最高(分別為根的8 和5倍), 其次為CARHR256690 (分別為根的5 和3 倍)。

圖6 ChSODs 在不同組織中的表達。1~10 見表2。n=3; 相同基因柱上不同字母表示差異顯著(P<0.05)。Fig. 6 Expression of ChSODs in different tissues. 1-10 see Table 2. n=3; Different letters upon column of the same gene indicate significant difference at 0.05 level.
2.6 ChSOD 蛋白的二級、三級結構預測
利用SOMPA 預測可知,在ChSOD 家族成員的蛋白質二級結構中,α-螺旋、無規則卷曲和延伸鏈較高,而β-轉角較低。其中多數蛋白質的α-螺旋占25.08%~54.55%,無規則卷曲為29.82%~56.36%,延伸鏈為11.69%~32.89%,而β-轉角絕大多數低于10%。因此,預測在ChSOD 蛋白二級結構中,α-螺旋、無規則卷曲起主要作用(表3)。

表3 ChSOD 蛋白二級結構Table 3 Secondary structure of ChSOD
SWISS-MODEL 對ChSOD 家族成員的蛋白三級結構進行預測,ChSOD 蛋白均含有α-螺旋、β-折疊、無規則卷曲等空間構象,但結構具有差異性;另外,各ChSOD 蛋白含有的螺旋-轉角-螺旋(HTH)數目存在差異(圖7)。

圖7 ChSOD 蛋白的三級結構Fig. 7 Tertiary structure of ChSOD
3 結論和討論
SOD 是植物體內清除氧自由基的關鍵酶,其活性與植物生長發育和抗逆性等密切相關[3-5]。近年來,隨著全基因組分析技術的發展,已對許多植物的SOD 家族成員進行了分析,并證明其參與調控植物的多重生理過程,如玉米(Zea mays)[21]、擬南芥[22]、水稻[23]、油菜(Brassica napus)[24]等。隨著基因組學的發展和碎米薺全基因組測序的完成,為解析碎米薺SOD基因家族成員的功能提供了理論和技術支持。
本研究中,通過生物信息學方法共鑒定出了10個碎米薺ChSOD基因,其結構簡單,且大多編碼親水性蛋白,表明碎米薺ChSOD基因結構較穩定,基因在進行復制時較難發生可變剪切,功能相對穩定。根據SOD 蛋白活性中心金屬輔因子的類型, 可將其分為Mn-SOD、Cu/Zn-SOD 和Fe-SOD,其中,組成型表達的Cu/Zn-SOD 和Fe-SOD 含量較穩定,既參與清除代謝過程中產生的ROS,也參與植物抗逆性形成;Mn-SOD 的表達極易受外界影響,在清除逆境脅迫產生的大量ROS 中發揮著重要作用[25]。研究表明,Cu/Zn-SOD 對植物的多種抗逆性尤為重要,如抗寒、抗旱、耐鹽堿等[24,26-27]。碎米薺10 個ChSOD基因中,有6 個薺ChSOD基因鑒定為Cu/Zn-SOD基因,說明碎米薺抗逆性與SOD 密切相關,且Cu/Zn-SOD 可能發揮主導作用。
前人根據SOD 蛋白的結構同源性推測,不同物種的Mn-SOD 和Fe-SOD 蛋白可能起源于同一祖先——光合細菌[28-29]。劉家林等[23]證實了水稻Mn-SOD 和Fe-SOD 可能起源于同一祖先。對碎米薺SOD蛋白進行預測,顯示Cu/Zn-SOD 歸于一支,而Mn-SOD 和Fe-SOD 歸于另一分支,表明在碎米薺中,Mn-SOD 和Fe-SOD 蛋白也可能起源于同一祖先。而不同SOD 家族成員間的理化性質的差異性可能與產生旁系同源基因,形成新的生物學功能,以更好地適應環境[30]。與水稻相比,碎米薺的ChSOD與擬南芥的AtSOD 親緣關系更近,可能與同屬于雙子葉植物有關,據此,可通過擬南芥AtSOD功能來推測碎米薺ChSOD基因的功能。基因在組織中的表達量與功能密切相關,ChSOD在葉中的高表達可能與葉片活躍的光合作用有關。CARHR085500和CARHR256690 在碎米薺葉和莖中表達量較高, 這與前人[31]報道Cu/ZnSOD轉錄產物主要集中于根和莖的結果一致。此外,高等植物的基因表達主要在轉錄水平上進行調控,且受多種順式作用元件和反式作用因子的相互調節。對ChSOD啟動子順式作用元件預測表明,碎米薺SOD啟動子中含有ABA、干旱、低溫、防御、應激和創傷等非生物脅迫響應元件,且含有ABA 和低溫響應元件的基因較多,推測ChSOD在響應非生物脅迫時,可能對ABA 和低溫脅迫響應更敏感。這有助于了解基因調控模式,對深入了解碎米薺的生長機制及其外界環境的調控模式具有重要意義。
在碎米薺全基因組中,共鑒定出10 個SOD基因,分布在7 條染色體上;ChSOD 蛋白的分子量與氨基酸數目成正比,且多數為酸性蛋白;ChSOD基因含有2~11 個外顯子。碎米薺SOD基因包含6 個Cu/Zn-SOD、3 個Fe-SOD和1 個Mn-SOD,可分為2 個亞家族,位于同一亞族的SOD基因具有相似的保守基序和基因結構。碎米薺ChSOD 與擬南芥AtSOD 的同源性較高。qRT-PCR 分析表明,ChSODs在碎米薺根、莖和葉中均有表達,以葉中的表達量最高。CARHR085500 在碎米薺葉和莖中的表達量最高,其次為CARHR256690。在響應非生物脅迫時,ChSODs 對ABA 和低溫脅迫響應較敏感。ChSOD蛋白質的二級、三級結構中,以α-螺旋和無規則卷曲比例較高,可能起主導作用,但不同ChSOD 蛋白質的空間構象和螺旋-轉角-螺旋(HTH)存在差異。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
