丹參SmVS 基因的克隆和表達分析
2022-06-10 10:30:26吳麗萍蘇興隆曹夢陽王兆健邢世海
熱帶亞熱帶植物學報 2022年3期
吳麗萍, 蘇興隆, 曹夢陽, 王兆健, 邢世海,2*
(1. 安徽中醫藥大學藥學院,合肥 230012;2. 安徽省中醫藥科學院中藥資源保護與開發研究所,合肥 230012)
丹參(Salvia miltiorrhiza)為唇形科(Labiatae)鼠尾草屬多年生直立草本植物,藥用部分為干燥根和根莖,為我國常用的傳統中藥材[1]。丹參主產于安徽、四川、山西、江蘇、河北等地,其性微寒、味苦、歸心、肝經,是治療心腦血管疾病的傳統中藥材[2]。現代藥理研究表明,丹參具有保護心血管系統[3]、防止動脈粥樣硬化[4]、保護神經系統[5]、改善微循環[6]和抗炎抗菌[7]等多種藥理作用,臨床常用于心腦血管疾病、癌癥、糖尿病和肝病等的治療[8-10]。現代化學分析表明丹參的化學成分以脂溶性二萜丹參酮類和水溶性丹酚酸類為主,還有多糖類、黃酮類、甾體類等成分[11-12];近年來還報道丹參中含有生物堿成分[13]。目前對丹參的研究大多集中在丹參酮和丹酚酸上,其生物合成途徑大部分已被解析[14-15];而關于丹參生物堿的研究甚少,尚無其合成途徑的報道。在生物堿中,萜類吲哚生物堿是一類非常重要的具有萜環和吲哚骨架的生物堿化合物,具有顯著的藥理學活性,臨床主要用于抗癌、抗心律失常和抗瘧疾等[16]。
單萜吲哚類生物堿阿嗎靈,其合成路徑已比較透徹[17],其中Vinorine 合成酶(vinorine synthase, VS)是阿嗎靈合成途徑中的1 個關鍵酶,催化蛇根精型生物堿16-epi-vellosimine 可逆性地合成阿嗎靈型生物堿Vinorine,Vinorine 合成酶連接2 種不同類型的生物堿,即蛇根精型(Sarpagan)和阿嗎靈型生物堿(Ajmalan)[17]。Vinorine 合成酶是第一個三維結構被解析的酰基輔酶A 依賴型酰基轉移酶(BAHD)家族成員,BAHD 得名于從植物中分離出的前4 種家族酶,該家族成員在多種次生代謝產物的生物合成中起著重要作用[18-19],但目前在丹參中尚未見報道。本研究從丹參中克隆SmVS基因,對其編碼蛋白進行生物信息學分析,利用實時熒光定量PCR 技術(RT-qPCR)檢測SmVS基因的表達模式,為進一步研究其功能提供理論基礎。
1 材料和方法
1.1 材料
材料采自安徽中醫藥大學藥園,生長光/暗周期為16 h/8 h,經安徽中醫藥大學俞年軍教授鑒定為唇形科丹參(Salvia miltiorrhiza)。采集丹參后用自來水清洗表面雜質,置于冰盒中轉移至實驗室,用蒸餾水沖洗3 次,吸干水分立即用液氮冷凍提取總RNA。
1.2 總RNA 提取和cDNA 的合成
采用改良的TRIzol 法[20]提取丹參根和葉的總RNA,使用FastQuant RT Kit (Tiangen Biotech,北京)試劑盒合成cDNA。
1.3 引物設計和PCR 擴增
根據丹參轉錄組數據庫[21],查找與SmVS同源的cDNA 序列用Primer 5.0 軟件設計引物(表1)進行特異PCR 擴增。引物序列由上海生工生物工程股份有限公司合成。
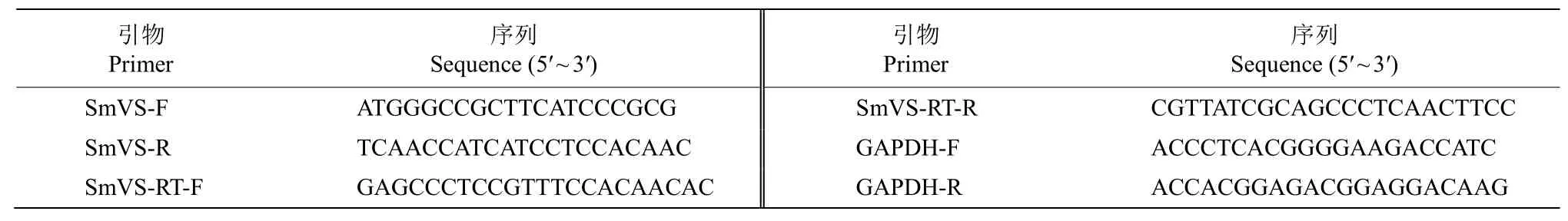
表1 引物序列Table 1 Primer sequences
1.4 生物信息學分析
根據PCR 擴增得到的基因全長序列,利用ORF Finder 網站(http://www.ncbi.nlm.nih.gov/projects/gorf/)尋找開放閱讀框(ORF),并翻譯出氨基酸序列;在線進行序列同源性比對(http://blast.Ncbi.Nlm.Nih.gov),并進行蛋白質理化性質分析(http://web.expasy.org/protparam); 用TMHMM 在線程序(http://www.cbs.dtu.dk/services/TMHMM-2.0)分析蛋白質的可能跨膜區;利用PSORT 軟件(https://www.genscript.com/psort.html)預測蛋白的亞細胞定位;利用GOR4 軟件(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)預測蛋白的二維結構;用SWISS-MODEL 軟件(https://swiss model.expasy.org/)進行編碼蛋白的三維結構預測;并利用MEGA X 軟件(Neighbor-joining,鄰位相連法)構建系統進化樹。
1.5 SmVS 基因表達的RT-qPCR 分析
采用SYBR Premix Ex TaqTM試劑(TaKaRa)和Roche Z480 實時熒光定量PCR 儀,以GAPDH作為內參基因,設計定量引物SmVS-RT (表1),參照韓榮春等[22]的方法進行RT-qPCR 反應,比較SmVS基因在葉和根中的表達差異,并分析其在葉中表達的日變化規律(n=3)。
2 結果和分析
2.1 SmVS 基因的克隆
采用特異性引物進行PCR 擴增,取5μL 產物進行凝膠電泳檢測,結果表明克隆得到的條帶與轉錄組預期基因片段大小一致(圖1)。將其核苷酸序列(圖2)與其他植物VS基因進行比對,表明該基因與其他植物的同源基因具有很高相似性,因此命名為SmVS。基因全長為726 bp,編碼241 個氨基酸(圖2)。該基因在GenBank 的登錄號為MW802630。
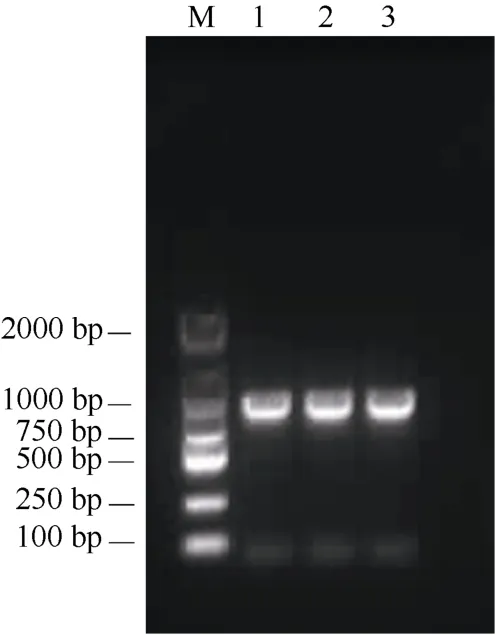
圖1 SmVS 基因PCR 擴增產物電泳圖。M: DL2000; 1~3: SmVS 基因。Fig. 1 Electrophoretic diagram of PCR amplification products of SmVS. M:DL2000 marker; 1-3: SmVS.

圖2 SmVS 基因的核苷酸序列和編碼的氨基酸序列Fig. 2 Nucletide sequence of SmVS and encoded amino acid sequence
2.2 生物信息分析
利用ProtParam 軟件對SmVS編碼的蛋白進行理化性質分析, 結果表明,SmVS 蛋白分子式為C1200H1857N329O347S13,由20 種共241 個氨基酸組成,相對分子量為26 861.72,原子總數為3 746,等電點(pI)為5.66,不穩定系數為43.95,預估在哺乳動物體外的網織紅細胞的半衰期是30 h、酵母體內大于20 h、大腸桿菌體內大于10 h,脂肪族指數74.02,總平均親水性為-0.208。跨膜區分析表明SmVS 未形成跨膜區結構域,屬于非跨膜蛋白。預測SmVS蛋白的亞細胞定位于線粒體,可能性為73.9%;定位于細胞質的可能性為17.4%;定位于細胞質膜和細胞核的可能性較低,均為4.3%。
蛋白質二級結構域預測表明(圖3: A),約有35.27%的氨基酸(85 個)以α螺旋形式存在,17.01%的氨基酸(41 個)為延伸主鏈,4.98%的氨基酸(12 個)為β轉角,42.74%的氨基酸(103 個)為無規則卷曲,表明該蛋白二級結構以α螺旋和無規則卷曲為主。用SWISS-MODEL對SmVS蛋白的氨基酸序列進行三維結構預測(圖3: B),該蛋白的主要結構元件還是螺旋、折疊和卷曲,與二級結構的預測結果相符。
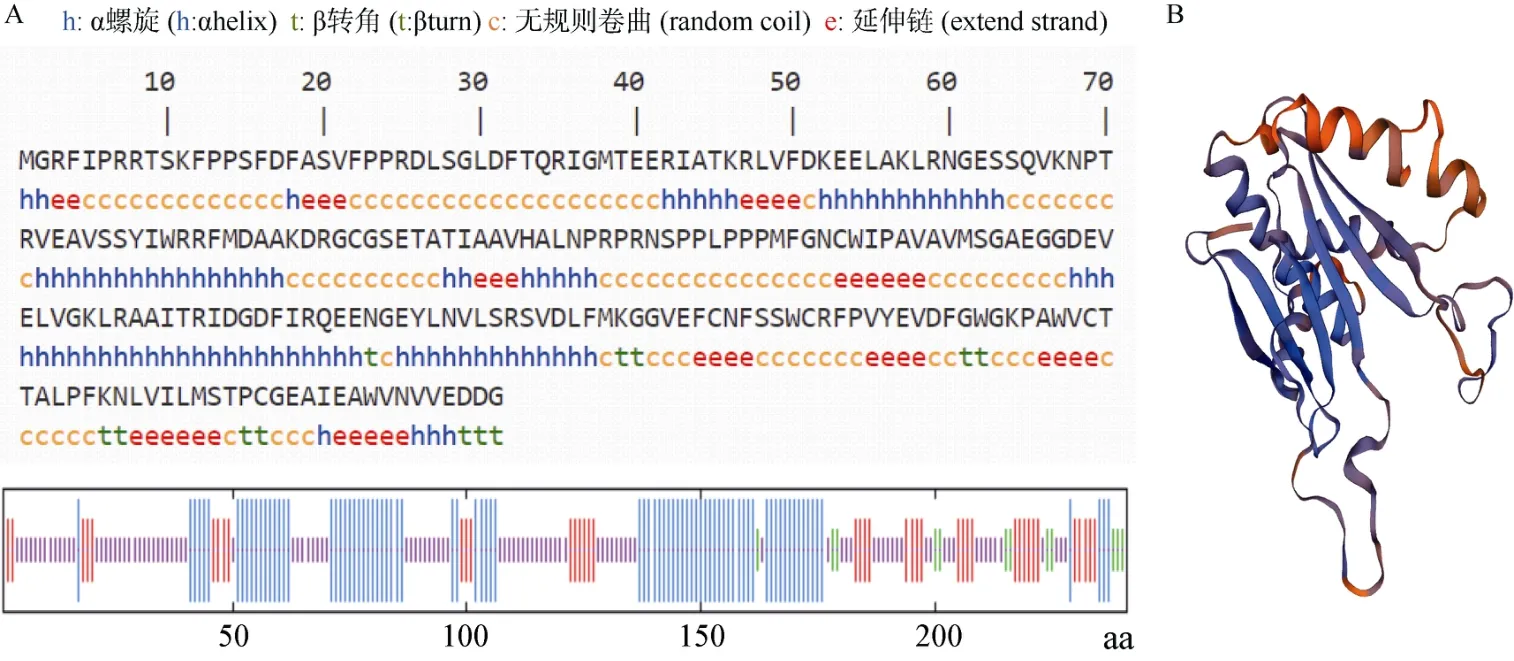
圖3 SmVS 蛋白二級結構(A)和三維結構(B)預測Fig. 3 Prediction of secondary (A) and three dimensional (B) structures for SmVS
2.3 SmVS 蛋白的系統進化分析
選取6 種植物的VS 氨基酸序列,利用DNA MAN 軟件進行序列多重比對分析(圖4: A),結果表明,這些VS 蛋白間具有較高的相似性。同時,SmVS蛋白功能域的氨基酸組成與其他植物幾乎一致,具有相同的保守域(Motif 1)(圖4: B),其中DFGWG結構域是幾乎所有BAHD 家族成員都有的,然而該結構域在不同酶中會有些許差異,如DFGWG 結構域在楊樹(Populussp.)的BAHD 家族基因中成為DYGWG、DFGFG、DFGWA、NFGWG、DFGWK、DLGFG 和NLGWG[18]。

圖4 VS 的氨基酸序列比對(A)和保守域(B)分析Fig. 4 Alignment of amino acid sequence and conserved domain of VS
根據NCBI Blast 對比結果,SmVS 蛋白的氨基酸序列與其他植物的同源性較高,與野生油橄欖(Olea europaeavar.sylvestris)的VS 氨基酸序列相似度為64%。在MEGA X 軟件平臺上采用NJ 法(bootstrap 設置為1 000)對11 種植物的VS 氨基酸序列進行系統進化分析(圖5),包括丹參(MW802630)、野生油橄欖(LOC111382475)、歐基尼奧伊德斯種咖啡(Coffea euge-nioides, LOC113763265)、煙草(Nicotiana tabacum, LOC 107765976)、茶樹(Camellia sinensis, LOC114266018)、牽牛(Ipomoea nil, LOC109155698)、芝麻(Sesamum indicum, LOC105168028)、苦瓜(Momordica charantia,LOC111007275)、南瓜(Cucurbita moschata, LOC11 1439839)、哥倫比亞錦葵(Herrania umbratical, LOC11 0418295)和胡桃(Juglans regia, LOC108985508)。結果表明,SmVS 與野生油橄欖的VS 聚為一支,親緣關系最近, 其次是歐基尼奧伊德斯種咖啡。系統進化和序列比對結果一致, 相似性越高親緣關系越近,可見Vinorine 合成酶在進化過程中是相對保守的。
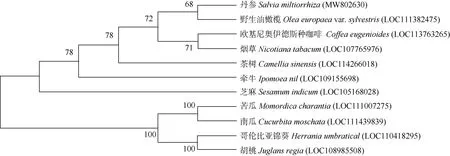
圖5 基于氨基酸序列的VS 系統進化樹Fig. 5 Phylogenetic tree of VS based on amino acid sequences
2.4 實時熒光定量表達分析
為了解SmVS基因在丹參中的表達模式,以GAPDH為內參基因,采用RT-qPCR 技術檢測SmVS基因的相對表達量。可見,SmVS基因在根和葉中均有表達,根中的表達是葉中的近2 倍(圖6: A), 且葉中SmVS基因在21:00 達最大值(圖6: B)。

圖6 SmVS 基因的表達模式。柱上不同字母表示差異顯著(P<0.05)。Fig. 6 Expression pattern of SmVS. Different letters upon column indicate significant differences at 0.05 level.
3 結論和討論
本文從丹參中克隆得到了SmVS基因,全長為726 bp。生物信息學分析表明,SmVS基因編碼241個氨基酸,SmVS 蛋白的分子量為26 861.72,等電點PI 為5.66,為非跨膜蛋白,推測其定位于線粒體。氨基酸同源性分析表明,SmVS 蛋白功能域的氨基酸組成與其他植物的VS 有較高的一致性,具有相同的保守域(Motif 1),SmVS 蛋白具有BAHD 家族的保守結構域DFGWG,因此,推測SmVS基因可能屬于BAHD 家族成員。系統進化樹分析表明丹參的SmVS 蛋白與野生油橄欖親緣關系較近。RT-qPCR結果表明,SmVS基因在丹參根中的表達量比葉中高,說明其在丹參中具有組織特異性;葉中SmVS在21:00 時表達最高。這些為丹參生物堿的代謝調控研究提供了基礎數據。
單萜類吲哚生物堿阿嗎靈,1931 年首次從蛇根木(Rauvolfia serpentina)樹皮中分離得到[23],1959年上市,屬于Ia 類抗心律失常藥物,臨床上主要用于治療預激綜合征伴發的心律失常[24]。乙酰輔酶A依賴酶的Vinorine 合成酶是從蛇根木(Rauvolfifia serpentina)和直瑞茲亞(Rhazya stricta)的雜交細胞懸浮培養物中分離出來的[25]。乙酰輔酶A 依賴的酰基轉移酶在植物細胞的次生代謝中起重要作用[26]。用于癌癥治療的二聚生物堿長春堿和長春新堿的生物合成前體單萜吲哚生物堿長春堿在長春花(Catharanthus roseus)的生物合成過程中,發生了乙酰轉移[27-28];二萜生物堿紫杉醇在東北紅豆杉(Taxus cuspidata)中的生物合成中,涉及到幾種酰基轉移酶,其中有2 種乙酰輔酶A 依賴性酶[29]。在阿嗎靈生物合成途徑中,乙酰輔酶A 依賴型Vinorine合成酶通過連接2 種不同類型的生物堿, 即蛇根精和阿嗎靈生物堿,在合成代謝途徑中起重要作用。
SmVS編碼的氨基酸序列和其他6 種植物具有相同的保守域,即SmVS 蛋白具有BAHD 家族成員的保守結構域DFGWG,因此,推測SmVS 屬于BAHD 家族的一員。最近幾年報道的BAHD 家族成員數目不斷增長,目前在水稻(Oryza sativa)、楊樹、擬南芥(Arabidopsis thaliana)中分別有119、94 和64個相關基因[30-31],而丹參中還未見報道。BAHD 家族中大多數功能已知的成員參與很多次生代謝過程,如花香、花青素的生物合成, 尤其是許多具有臨床治療價值的生物堿的合成,如嗎啡、紫杉醇、長春花堿和阿嗎靈等[30]。該家族成員有很多保守區域,第1 個是位于酶中心HXXXD 結構域,對于乙酰基轉移酶的反應非常重要,第2 個是位于碳端的DFGWG 結構域,是所有BAHD 成員都擁有的結構域[17]。
表達分析表明,SmVS基因的轉錄水平受不同組織和時間的影響,在根中表達量比葉中高,說明丹參生物堿多在根中合成。在基因工程中利用發根農桿菌誘導丹參發根,比利用根癌農桿菌誘導能產生更多的阿嗎靈型生物堿vinorine;葉片中SmVS基因在白天的轉錄水平極低,夜間逐漸升高,21:00達到最大值,可見該生物堿主要在夜間合成。植物的生長發育受外界很多因素影響,如光照、低溫、干旱等,其中光照是一種重要的環境因子,植物體內存在很多光響應的轉錄因子,能夠感受光的刺激而誘導下游基因的表達,最終影響植物的生長發育和代謝等[32-33]。Liu 等[34]報道長春花葉中生物堿vindoline 的合成受光調控,其光應答轉錄因子CrGATA1 可以調控vindoline 生物合成,CrGATA1的瞬時過表達導致長春花文多靈途徑基因的上調,最終增加了vindoline 的積累。因此,我們推測SmVS基因也是一個光應答基因,其轉錄水平受光的調控。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
