雌雄異株植物長梗柳傳粉系統和生殖策略研究
2022-06-10 10:30:28曾思文梁恬李林鑫邢曉閣陳泓樸何理
熱帶亞熱帶植物學報 2022年3期
曾思文, 梁恬, 李林鑫, 邢曉閣, 陳泓樸, 何理,2*
(1, 福建農林大學林學院, 福州, 350000; 2, 上海辰山植物園, 華東野生瀕危資源植物保育中心, 上海, 201602)
雌雄異株植物約占被子植物的5%~6%[1], 其繁殖資源分配主要包括繁殖分配和性別分配[2]。雌雄異株不同性別植株在生殖功能上存在顯著差異, 雄性生殖活動僅需要產生雄花, 而雌性在生殖過程中需要產生雌花和果實, 其能量消耗和資源分配遠遠大于雄性[3]。因此多年生植物中, 雄株常在開花階段分配更多資源, 雌株則在結實階段分配更多資源[4]。繁殖策略不僅與環境資源(如光、溫、水等)有關, 還與植株大小、花結構及傳粉系統等密切相關[2,5-6]。
風媒傳粉系統及蟲媒傳粉系統被認為是雌雄異株植物主要的傳粉系統, 兩種及以上傳粉系統同時出現在同種植物中即稱為混合傳粉系統[7]。風媒花一般通過增加雄花數量來維持花粉可用性以確保繁殖成功率[8-10]。蟲媒花中, 昆蟲為了獲得花報酬(如花粉、花蜜等)而進行訪花[11]。“傳粉者限制假說(Pollinator limitation hypothesis)”認為傳粉者通常因雄花能同時提供花粉和花蜜2種花報酬, 而歧視只提供花蜜的雌花, 出現偏好訪問雄花的現象[12]。為平衡傳粉者偏好訪問的選擇壓力, 雌花會通過氣味模仿[13]、獎勵機制[14-15], 以及欺騙引導[16]等策略來吸引傳粉昆蟲, 以提升繁殖效率。對于具有混合傳粉系統的雌雄異株植物, 如何在雌雄個體中進行生殖資源分配來提升繁殖效率值得深入研究。
大多數植物是依賴花部釋放的揮發物向傳粉者傳遞信息[17-18]。Tollsten 等檢測了柳屬(Salix)植物黃花柳(S.caprea)、灰柳(S.cinerea)和S.repens的雌雄花序揮發物, 認為柳屬植物雌雄株的揮發物保持了高度的相似性, 推測雌雄花序相似的揮發物能誘使傳粉者同時訪問雌雄花, 保證傳粉的有效性[13,15]。D?tterl 等[19]對黃花柳花香揮發物進行了研究, 認為雄株的1,4-二甲氧基苯含量比雌株更高, 因而更具吸引力。在自然居群中黃花柳雌株明顯多于雄株, 雌雄花序揮發物的相似性或雄株具有更高比例的吸引傳粉者的揮發物, 都能減少花粉限制的風險[20]。這一現象是否在其他柳屬植物中存在, 仍需進一步研究。
楊柳科(Salicaceae)柳屬在全球約有450 種[21],均為雌雄異株植物。該屬植物傳粉系統常為風媒和蟲媒混合的傳粉系統[7,22]。不同地區和不同物種柳屬植物的蟲媒和風媒的傳粉效率占比不同, 從幾乎完全風媒到幾乎完全蟲媒不等, 如S.lasiolepis蟲媒傳粉效率達99%, 即幾乎完全蟲媒[23]; 而S.lanata和S.myrsinites的蟲媒傳粉效率僅8%和2%, 即幾乎完全風媒; 另外,S.lapponum、S.herbacea、S.reticulata的蟲媒傳粉效率分別為47%、57%和52%,是典型的混合傳粉系統[24]。而柳屬植物在雌雄株中如何平衡生殖投資策略, 同時滿足風媒和蟲媒條件以提升繁殖效率, 這尚需深入探討。
本文以長梗柳(Salix dunnii)為研究對象, 通過調查福建省長梗柳野外居群的傳粉系統、主要訪花昆蟲, 性別比例、花和花序特征、揮發物的種類及相對含量, 探討長梗柳的傳粉機制和生殖分配策略。
1 材料和方法
1.1 研究地點
本研究主要在3 個野外居群開展,福建省三明市沙縣沙溪支流的郭墩村(117°38′55″ E, 26°25′39″N, 海拔190~229 m)和高地村(117°39′48″ E, 26°24′59″ N, 海拔217.4~224.2 m),及福建省武夷山市崇陽溪的東邊村(118°2′46″E, 27°46′45″N, 海拔205~212 m),均處于亞熱帶海洋性氣候區, 三明市年均溫20.9 ℃, 年均降水量1 268 mm, 武夷山市年均溫21.1 ℃, 年均降水量1 158 mm, 氣候溫和濕潤。
1.2 花和花序的特征統計和性別比例
對郭墩村和高地村2 個長梗柳野外自然居群的雌雄株花序進行抽樣調查和測量[20,25]。在調查地沿岸隨機抽取長勢良好的植株, 兩地各選擇4~5 棵雌株和雄株進行調查(兩地共19 株, 雌、雄分別為9 和10 株), 每株選擇樹冠下部長勢良好、長度大于20 cm的末端分枝枝條5~13 根, 統計和測量分枝長度和花序數, 計算單位長度的花序數, 使用精度為0.01 mm的游標卡尺測量花序長度, 并利用體視顯微鏡(LEICA DFC550)拍攝花部。此外, 每株統計8~46個單花序的花數量, 枝條單位長度花序數=單枝條花序數/枝條長度,單位長度以1 cm 計。雌花序直接用計數法進行統計; 雄花序近5 朵小花排列成一輪(圖1: A), 因此雄花序花數量以輪數×5 進行統計。
兩地隨機抽取雌雄各4 株進行統計分析。以植株性別為因變量, 即1 (雄性)或0 (雌性), 以株數為數據分組(表1), 隨機不放回抽取2 個居群雌雄株各20 根枝條和分別300、40 個花序, 統計枝條單位長度花序數、花序長度和花序單花數, 運用廣義線性混合模型(generalized linear mixed model, GLMM)[26-27]對長梗柳雌雄株枝條單位長度花序數、花序長度和單花序花總數差異進行顯著性程度檢驗(P<0.05)。使用Microsoft Excel 2016 進行數據統計計算。
根據線路調查法, 對郭墩村、高地村和東邊村河流兩岸分布的長梗柳雌雄株個體數量進行統計。使用Pearson’s Chi-squared test 對性別比例(雌/雄)偏離1∶1 零假設的顯著性程度(P<0.05)進行檢驗。
1.3 長梗柳傳粉系統
選擇郭墩村和高地村長梗柳各5 棵雌株, 分別編號為XG1~XG5 和LT1~LT5, 于3 月2 和3 日(長梗柳初花期)進行套袋套網試驗[23-24,28]。長梗柳花小且多, 因此本試驗對枝條整枝進行套袋套網。設置3 個處理, 每株設置3 個重復:①套硫酸紙袋; ②套100 目圓柱尼龍網袋, 用鐵絲網支撐; ③僅標記不套袋。分別檢測長梗柳無融合生殖、風媒傳粉和自然結實情況。
花期過后(約4 月下旬)將網袋和硫酸紙袋摘除,于果期采收枝條, 利用體視鏡統計正常發育的果和空果數量,以及果內發育和敗育的種子數量, 計算座果率和結實率,座果率=(每個花序正常發育的果實數/該花序所有果實數)×100%;結實率=發育正常的種子數/總種子數×100%,其中, 正常發育果實內含至少1粒發育正常的種子, 而敗育果實則含敗育種子或空果; 發育正常的種子為綠色、圓潤,而敗育種子為枯黃色、皺縮[29]。使用單因素方差分析(ANOVA)來檢驗平均座果率、平均結實率是否在不同處理(套硫酸紙袋、套網袋及無處理)之間存在差異。
1.4 主要訪花昆蟲和訪花頻率
選擇郭墩村長梗柳盛花期長勢良好的雌雄株進行連續3 d 觀察, 每日觀察和捕捉時間為8:00—18:00 (計劃觀察時間為2019 年3 月12—14 日, 由于3 月14 日陰雨天氣, 為了數據準確穩定, 順延至3 月15 日), 觀察訪花昆蟲種類和訪花頻率[30-31]。我們對訪問并停留在花序上超過0.5 min 的昆蟲進行捕捉, 捕捉范圍網桿(5 m)所能及高度內的訪花昆蟲, 雌雄株每30 min 交替捕捉, 統計每30 min 雌雄株上出現的昆蟲數量, 將捕獲的昆蟲放入裝有乙酸乙酯的毒瓶中殺死, 制成標本, 標本交由中國科學院動物研究所鑒定。
采用Microsoft Excel 2016 和R v3.6.3 進行數據統計計算并出圖, 使用Pearson’s Chi-squared test 對訪花頻率進行顯著性程度檢驗(P<0.05), 按訪花昆蟲對長梗柳每個性別平均日訪花次數確定該昆蟲訪花頻率[30]。
1.5 花序揮發物分析
取東邊村長梗柳盛花期長勢良好的雌株和雄株各4 株, 每株采集3 個重復, 共計24 個花序樣品;同時收集當地空氣作為空白對照。采集的花序送往福州大學國家科技園測試中心測定揮發物。
利用頂空固相微萃取(SPME)法對長梗柳花序揮發物進行采集, 選用非極性的PDMS 萃取頭(65μm PDMS/DVB), 利用全二維氣相色譜-高分辨飛行時間質譜聯用儀(LECO GC×MS PEGASUS HRT 4D plus)進行分析。采氣前孵化(35 ℃) 1 h, 使花香揮發,同時萃取頭在280 ℃下老化0.5 h,以后萃取前均280 ℃老化5 min。將老化的萃取頭插入采樣瓶中,置于待測樣品上方約1 cm 處萃取0.5 h,拔出針頭后入GC-MS 進樣口解吸4 min。一維GC 條件為:Rxi-5ms 石英毛細管柱(30 m×0.25 mm×0.25μm),進樣采用不分流模式, 柱溫40 ℃保持7 min, 以3 ℃/min 升溫至260 ℃, 保持10 min。傳輸線溫度280 ℃, 進樣口溫度280 ℃。載氣為高純氦氣, 流速為1.4 mL/min。二維GC 條件為: Rxi-17Sil 石英毛細管柱(2 m×0.25 mm×0.25μm), 二維爐溫在一維爐溫基礎上+5 ℃, 調制器溫度在二維爐溫基礎上+15 ℃, 調制周期4 s, 每次熱吹0.8 s, 每次冷吹1.2 s。MS 條件為: 以電子轟擊方式(EI)電離, 掃描范圍35~500 amu, 電子能量為70 eV; 離子源溫度250 ℃, 溶劑延遲240 s, 掃描速度200譜圖/s, 一維和二維MS 條件一致。
揮發物鑒定首先利用儀器自帶的標準譜庫(NIST 2017 庫)和增加wiley10 數據庫檢索, 并參閱文獻資料加以確定化合物。可能由于樣品濃度過低或儀器質譜檢測限較高導致2 個雌花序樣本和1 個雄花序樣本數據無法使用, 最終我們使用了10 個雌花序樣本及11 個雄花序樣本進行后續統計分析。根據CAS 號, 利用化源網(https://www.chemsrc.com/)確定化合物名稱。對雌雄花序揮發物含量的分析時,要求各個樣本揮發物的篩選條件為相對含量大于1%且與標準譜庫的相似度大于85%, 篩選過后有1個雌花序樣本不滿足上述條件, 因此僅使用了9 個雌花序樣本及11 個雄花序樣本進行NMDS 分析。
根據非度量多維標度分析法(NMDS), 使用Vegan (https://github.com/vegandevs/vegan) (R v3.6.3)
對雌雄株花序揮發物組成成分進行劃分, 以Stress值(值越小, 擬合效果越好)作為判斷NMDS 排序結果擬合優度的標準, 并對不同樣本的相似性在二維平面進行可視化展示[32]。采用Mann-WhitneyUtest對長梗柳雌雄花序揮發物相對含量進行顯著性檢驗。
2 結果和分析
2.1 花和花序統計及性別比例情況
長梗柳花期在3 月上旬至4 月下旬, 花藥顏色為黃色, 雄花和雌花均含有腺體(圖1: A), 可以為蜜蜂提供花蜜來源。

圖1 長梗柳雌雄花和花序形態。A: 雄花和雌花; B: 花序橫切,示著生方式, 單花約5 朵一輪; C: 雌花序; D: 中華蜜蜂訪問雄花序; E: 雌花序花蜜珠。標尺=1 mmFig. 1 Morphology of male and female flowers and catkins of Salix dunnii. A: Male and female flower; B: Cross section of male catkin showing about five flowers per round; C: Female catkin; D: Apis cerana was visiting male catkin; E: Nectar of female catkin. Bars=1 mm
運用GLMM 對長梗柳雌雄株1 cm 枝條花序數、花序長度和單花序花數進行統計(表1), 共測量197根枝條(雌株88 根, 雄株109 根)、3 044 個花序(雌花序1 199 個, 雄花序1 845 個), 并統計375 個花序(雌花序134 個, 雄花序241 個)上的花。結果表明,雄株單位長度枝條的花序數顯著多于雌株(M: 0.53±0.03,n=40; F: 0.41±0.03,n=40;F=8.31,P=0.005);
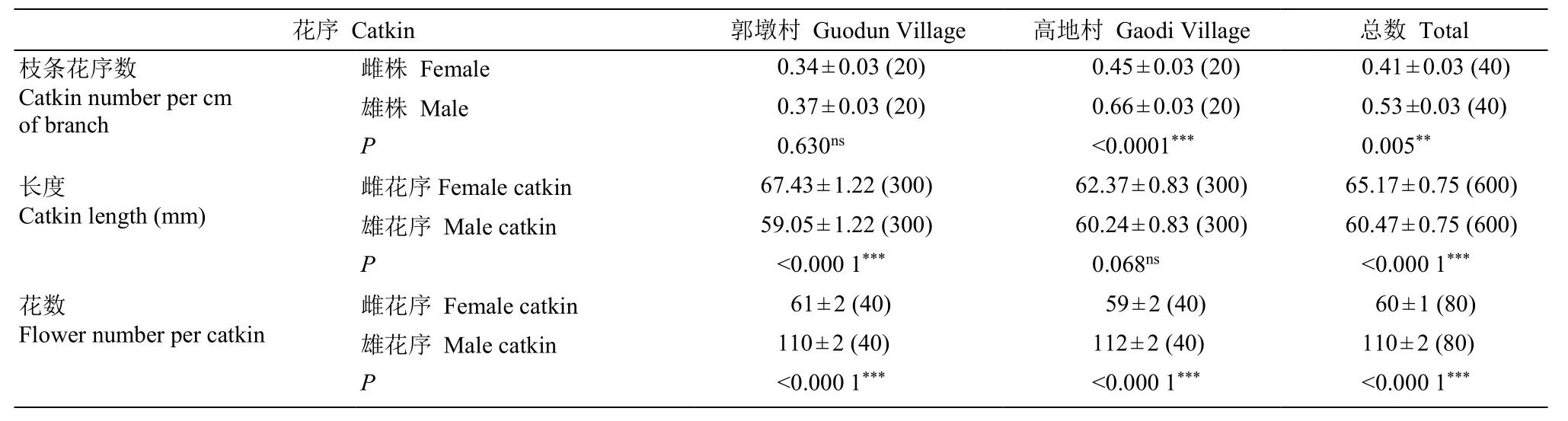
表1 長梗柳雌雄株的花序特征Table1 Catkin characters of female and male of Salix dunnii
此外雄株單花序的花朵數也明顯多于雌株(M: 110±2,n=80; F: 60±1,n=80;F=611.63,P<0.000 1),而雌株的平均花序長度顯著長于雄株[F: (65.17±0.75) mm,n=600; M: (60.47±0.75) mm,n=600;F=19.81,P<0.000 1]。可見, 長梗柳雌雄株的枝條花序數、花序長度和單花序花數的差異均達到顯著水平。
居群的性別比調查結果表明(圖2),東邊村顯著偏雄(♂∶♀=1.28∶1,n=336∶263,χ2=4.22,P=0.04<0.05), 郭墩村(♂∶♀=1.21∶1,n=318∶263,χ2=2.423,P=0.12)和高地村(♂∶♀=1.06∶1,n=132∶125,χ2=0.049,P=0.83)均不顯著偏離1∶1。
2.2 座果率及結實率
由于雨水和人為等因素的干擾, 部分套袋被破壞, 排除被破壞的套袋數據后, 最終統計了4 棵長梗柳雌株(郭墩村XG1、XG2 和高地村LT1、LT3)共4 053 個果實16 930 粒種子。其中, 套硫酸紙袋的枝條共產生297 個空果,沒有成熟果, 座果率0.00%; 套網袋的枝條共產生562個空果和427個成熟果, 座果率達43.90%, 成熟果里有718 粒發育種子和61 粒敗育種子; 自然狀態下的枝條共產生116個空果和2 551 個成熟果, 座果率高達90.90%, 成熟果中含14 844 粒發育種子和1 307 粒敗育種子。經單因素方差分析(ANOVA)表明, 自然傳粉的座果率顯著高于套網袋處理(F=9.63,P=0.02), 套網袋處理顯著高于套硫酸紙袋處理(F=8.94,P=0.02); 自然傳粉的結實率與套網袋處理的差異不顯著(F=0.87,P=0.39), 套網袋處理的平均座果率顯著高于套硫酸紙袋處理(F=14.53,P<0.05) (表2)。
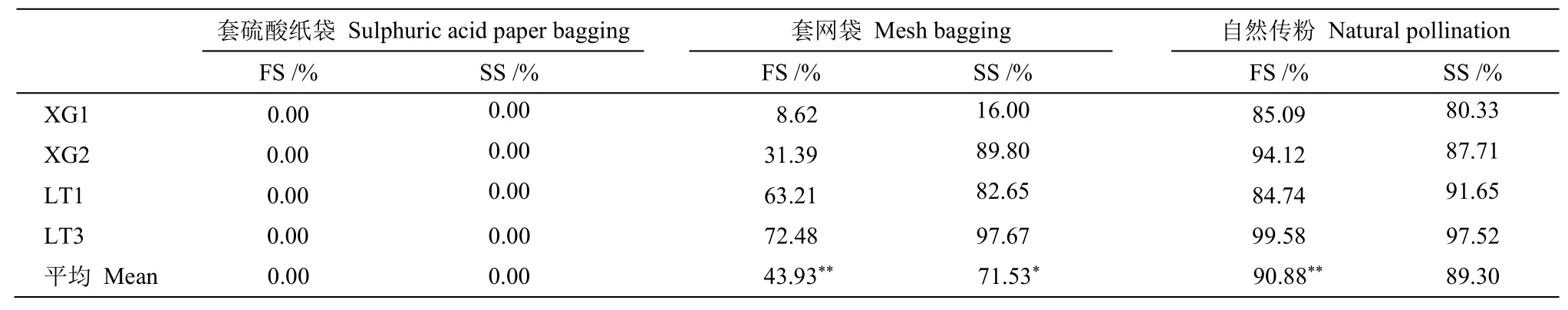
表2 長梗柳的座果率和結實率Table 2 Fruiting rate and setting rate of Salix dunnii
2.3 主要訪花昆蟲及訪花頻率
本研究共捕獲和鑒定長梗柳的訪花昆蟲4目22科40 屬44 種(表3), 主要是膜翅目(Hymenoptera)的蜜蜂科(Apidae)、隧蜂科(Halictidae)、胡蜂科(Vespidae)、地蜂科(Andrenidae)和葉蜂科(Tenthredinidae);雙翅目(Diptera)的實蠅科(Tephritidae)、食蚜蠅科(Syrphidae)、花蠅科(Anthomyiidae)、麗蠅科(Calliphoridae)、麻蠅科(Sarcophagidae)、舞虻科(Empididae)、毛蚊科(Bibionidae)和蠅科(Muscidae);鞘翅目(Coleoptera)的瓢蟲科(Coccinellidae)、偽葉甲科(Lagriidae)、花螢科(Cantharidae)、步甲科(Carabidae)、大蕈甲科(Erotylidae)、象甲科(Curculionidae)和天牛科(Cerambycidae);半翅目(Homoptera)的蝽科(Pentatomidae)和葉蟬科(Cicadellidae)。雌雄株共有訪花昆蟲包括蜜蜂科、隧蜂科、胡蜂科、葉蜂科、麗蠅科、毛蚊科、舞虻科、食蚜蠅科、花螢科昆蟲,雌株獨有的訪花昆蟲主要包括隧蜂科隧蜂屬、胡蜂科約馬蜂(Polistes jokahamae)、舞虻科平須舞虻屬、麻蠅科昆蟲,雄株獨有的訪花昆蟲主要包括瓢蟲科紅頸盤瓢蟲(Lemnia melanaria)以及偽葉甲科昆蟲。
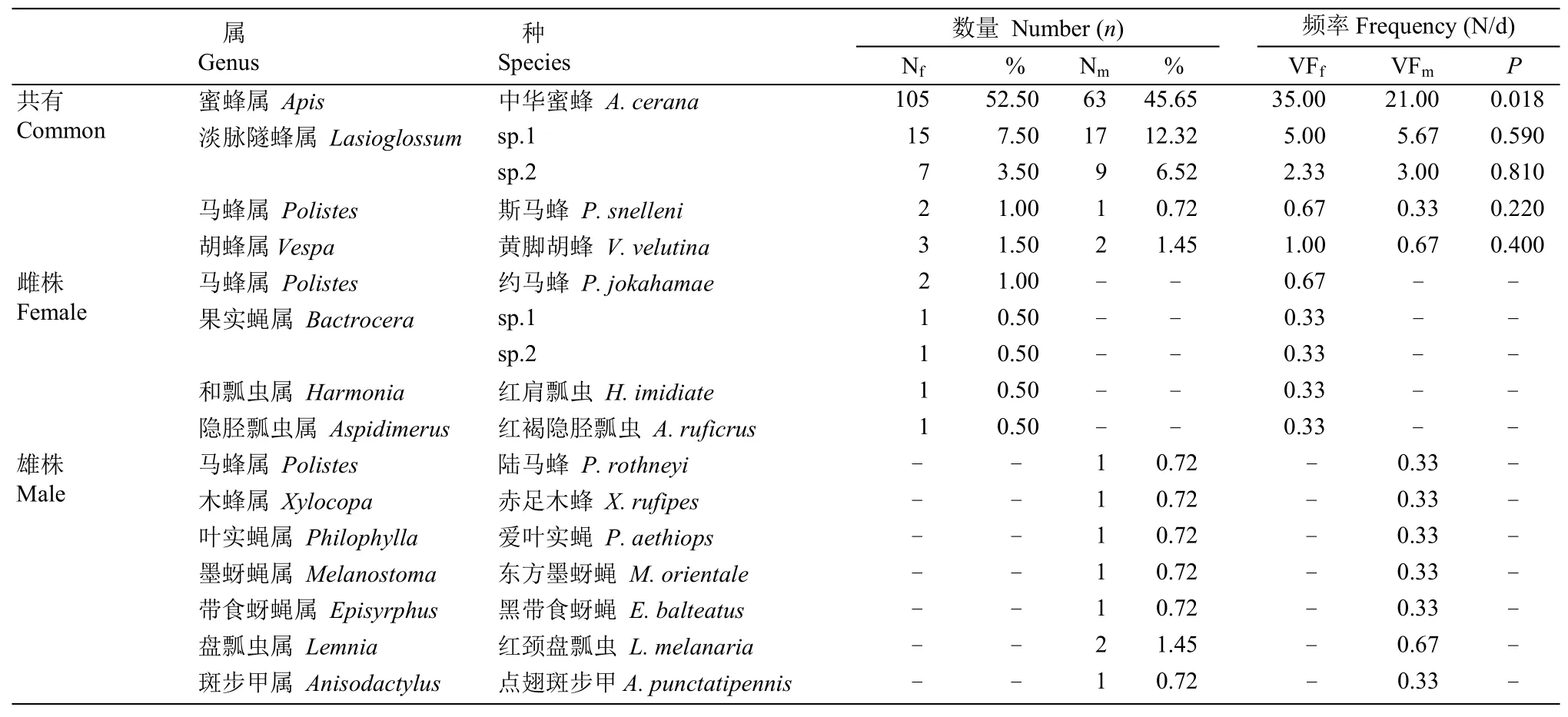
表3 長梗柳主要訪花昆蟲名錄Table 3 List of main flower-visiting insects of Salix dunnii
主要訪花昆蟲為蜜蜂科膜翅目的中華蜜蜂(Apis cerana, 1 793)和隧蜂科淡脈隧蜂屬昆蟲(Lasioglossumsp.1 和sp.2)。中華蜜蜂占雄株總訪花昆蟲的45.65%, 占雌株總訪花昆蟲的52.50%, 淡脈隧蜂屬昆蟲次之, 分別占雄雌株訪花昆蟲的18.84%♂和11.00%♀, 且停留時間為0.5~5 min。其他訪花昆蟲分別占雄雌株總訪花昆蟲的35.51%♂和36.50%♀。
對訪花的中華蜜蜂進行3 d 網捕,結果表明(圖2), 12 日訪問雌花的中華蜜蜂(37 只)多于訪問雄花的(12 只), 13 日訪問雌花的中華蜜蜂(24 只)與訪問雄花的(26 只)相近, 15 日訪問雌花的中華蜜蜂(44 只)多于訪問雄花的(25 只), 中華蜜蜂顯著傾向訪問雌花(χ2=8.07,P=0.018<0.05)。
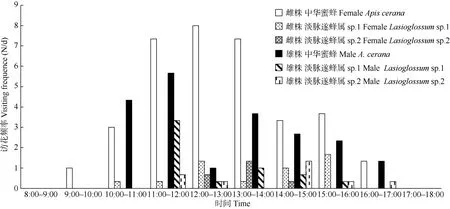
圖2 長梗柳雌雄株主要訪花昆蟲日訪花頻率Fig. 2 Daily visiting frequency of main visiting insects of male and female Salix dunnii
2.4 花序揮發物
雌株和雄株共有揮發物14 種, 雌株特有的9 種,雄株特有的15 種(表4)。雌雄株花序共有揮發性成分相對含量有所差異, 雌株比雄株多的有:β-石竹烯(P<0.001)、2-甲基丁腈(P<0.05)、環五聚二甲基硅氧烷(P<0.001)、2,2,4,6,6-五甲基庚烷(P<0.001)、八甲基環四硅氧烷(P<0.001)、Germacrene D 和2,3-dimethyl butanedinitrile; 雄株比雌株多的有:α-蒎烯、檜烯、trans-α-ocimene、月桂烯、(-)-β-蒎烯、(S)-(-)-檸檬烯和(3E)-4,8-二甲基-1,3,7-壬三烯。

表4 長梗柳雌、雄株花序的揮發性成分Table 4 Volatile compounds in female and male catkins of Salix dunnii
對雌雄花序中14 種共有揮發性成分進行NMDS分析(Stress=0.16), 結果表明雌雄花序揮發性成分存在部分重疊, 但分離顯著(圖3)。對雌雄花序不同揮發性成分相對含量進行Shapiro 正態性檢驗, Mann-WhitneyUtest 比較結果表明,β-石竹烯在雌花序中的含量與雄花序呈極顯著差異(P=0.000 093 62<0.001)。
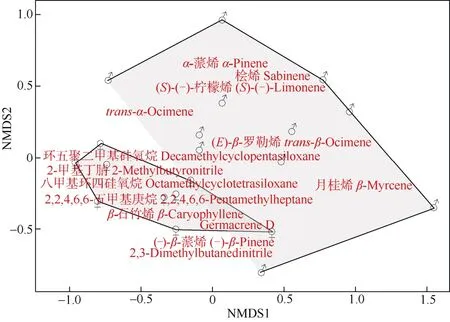
圖3 長梗柳雌雄花序共有揮發物NMDS 分析Fig. 3 Non-metric multidimensional scaling (NMDS) analysis of volatile components shared in male and female catkins of Salix dunnii
3 結論和討論
3.1 長梗柳的傳粉系統
套硫酸紙袋試驗結果表明, 長梗柳不存在無融合生殖的現象; 套網袋的平均座果率為43.9%, 而套硫酸紙袋的為0.00%, 二者差異顯著,說明風媒傳粉具有重要作用; 自然狀態的平均座果率為90.9%, 較套網袋處理的顯著增加, 說明昆蟲傳粉也具有重要作用[23], 因此長梗柳傳粉系統為風媒和蟲媒混合系統, 平均風媒、蟲媒傳粉效率分別為71.5%和28.5%,說明風媒傳粉效率更高, 這與Tamura 等[7]對S.miyabeana和S.sachalinensis傳粉系統的研究結果一致。雌雄株上同時出現且數量最豐富的昆蟲是膜翅目蜜蜂科蜜蜂屬的中華蜜蜂(45.65%♂, 52.50%♀), 其次是膜翅目膜隧蜂科淡脈隧蜂屬(Lasioglossum)的昆蟲(18.84%♂, 11.00%♀), 是長梗柳潛在的主要傳粉者, 中華蜜蜂訪問雌花頻率顯著高于雄花(P=0.018),淡脈隧蜂屬的2 種昆蟲訪雌雄花頻率差異不顯著。這進一步支持蜂類是柳屬植物的主要傳粉者[19,33]。
3.2 長梗柳的生殖投資策略
雌花序長度明顯大于雄花序, 較長的花序可能更有利于擴大吸附花粉的面積和訪花昆蟲的落腳面積, 從而提升結實率, 達到資源配比的最優值[24]。雄株的平均單枝著生花序數和平均花序花數均明顯多于雌株, 這與前人[8,19,22]的研究結果一致,可能利于風媒傳粉。
繁殖資源分配在個體水平可以體現在兩性的性別比例上[6,34], 長梗柳的3 個野外居群中雄株數量均較雌株多, 但其中2 個居群差異不顯著, 仍需長期更大尺度的調查, 以進一步確定[34-36]。在雌雄異株多年生植物中, 雄性早于雌性成熟的現象普遍存在, 性別比例會有所偏倚, 這可能是性分配差異所致[36-37]。此外, Pucholt 等[38]報道S.viminalis的雌雄株性比偏倚與ZW 性別決定系統有關,而長梗柳偏雄性比是否與其具有的XY 性別決定系統[39]有關尚需進一步研究。
長梗柳雌花β-石竹烯的相對含量[(14.58±8.34)%]顯著高于雄花(P<0.001), 而β-欖香烯[(0.18±0.54)%]、芳樟醇[(0.22±0.66)%]僅在雌花序中檢出。這些花香揮發性成分在吸引中華蜜蜂及其他訪花昆蟲中起到關鍵信號作用[40-43]。根據NMDS 結果,雌雄花序揮發性成分僅有部分重疊, 分離達到了顯著水平, 而雄性占據了更多的面積代表雄花序揮發性成分的相對含量更多, 但吸引主要訪花昆蟲中華蜜蜂的物質β-石竹烯在雌花序中的含量極顯著(P<0.001)高于雄花序。有研究表明,溫帶和寒帶柳屬植物的揮發性成分中均有對訪花昆蟲具有吸引作用的1,4-二甲氧基苯[44-45], 但我們在亞熱帶的長梗柳中并沒有鑒定出1,4-二甲氧基苯, 這可能是由于植物的花香氣味受到地理環境因素的影響而產生了相應的變化[46]。然而長梗柳花序揮發性成分多達38 種, 含量較高的揮發性成分,如α-蒎烯、2-甲基丁腈、環五聚二甲基硅氧烷和2,2,4,6,6-五甲基庚烷等是否具有吸引訪花昆蟲的作用尚不明確。
綜上所述, 長梗柳雄株在早期繁殖資源分配中投入更多的能量, 體現在花朵和花序數量上, 這是確保風媒傳粉效率的前提; 雌株則通過產生吸引訪花昆蟲的揮發性成分來抵抗“傳粉者限制”效應, 或許可以加強蟲媒傳粉的效率。本研究支持長梗柳為蟲媒風媒的混合傳粉系統, 通過風媒和蟲媒有效組合提升繁殖效率。
致謝特別感謝福州大學測試中心劉薇老師對揮發性成分進行測試, 中國科學院動物研究所吳超老師鑒定訪花昆蟲, 福建農林大學馬曉開老師對本文提出的建設性建議。
