內質網蛋白TMEM166對MPP+誘導的SH-SY5Y細胞氧化應激的影響及其機制
2022-12-10 07:28:14蒲明怡王喆廖肇忠鞏遵雙楊偉燕華君男李寧
精準醫學雜志 2022年6期
蒲明怡 王喆 廖肇忠 鞏遵雙 楊偉燕 華君男 李寧
(1 青島大學基礎醫學院,山東 青島 266071; 2 青島市膠州市婦幼保健計劃生育服務中心)
帕金森病(Parkinson’s disease,PD)是全球常見的人類第二大神經退行性疾病,目前仍沒有有效的治愈手段,只能通過藥物延緩其進程[1]。人口老齡化加劇、治療手段缺乏、靶向用藥困難等因素,使得這類老年慢性疾病的治療情況更加嚴峻[2]。因此,深入揭示PD的發病機制,尋找有效的治療靶點將為PD的治療提供新的策略。
內質網跨膜蛋白166(TMEM166),亦稱Eva-1同源蛋白A,最初是2007年在人腎臟cDNA文庫中鑒定出的一個與凋亡和自噬有關的內質網跨膜蛋白[3]。對TMEM166-/-和野生型小鼠腦的定量蛋白質組學分析發現,有11種蛋白質表達上調,17種蛋白質表達下調,這些蛋白質與ATP合成、氧化磷酸化和三羧酸循環有關[4]。并且通過基因富集分析進一步發現,這些失調的蛋白與一些神經退行性疾病,如阿爾茨海默病(AD)、亨廷頓病(HD)以及PD等密切相關[5],但是TMEM166在神經退行性疾病發病中的具體作用尚未見研究報道。
線粒體功能障礙引起的神經元氧化應激在PD的發展中起著至關重要的作用,MPP+處理的SH-SY5Y細胞是高度模擬神經細胞線粒體功能障礙導致氧化應激的PD細胞模型[6-7]。本研究嘗試初步探索TMEM166對MPP+誘導的SH-SY5Y細胞氧化應激的影響和機制,以進一步探討自噬與PD的相關性,為PD的治療提供新的靶點和策略。
1 材料與方法
1.1 細胞和質粒
人神經母瘤細胞系SH-SY5Y由我院生物化學與分子生物學實驗室提供。Myc-TMEM166過表達質粒和Myc空質粒由浙江大學劉偉教授饋贈。
1.2 試劑
CCK-8試劑盒(北京索萊寶科技有限公司),活性氧(ROS)檢測試劑盒(上海碧云天生物技術有限公司),1-甲基-4-苯基-吡啶碘化物(德國默克公司),3-甲基腺嘌呤(3-MA)(美國MCE公司),兔抗人TMEM166多克隆抗體(英國Abcam公司),兔抗人p62多克隆抗體(武漢三鷹生物技術有限公司),鼠抗人LC3單克隆抗體(北京博爾邁生物技術有限公司),熒光定量PCR試劑盒ChamQ SYBR Color qPCR Master Mix、RNA逆轉錄試劑盒HiScript?Ⅱ Q RT SuperMix(南京諾唯贊生物科技股份有限公司)。
1.3 細胞培養
將SH-SY5Y細胞培養于含體積分數0.01青霉素/鏈霉素溶液和體積分數0.10胎牛血清的高糖DMEM培養基中,置于37 ℃、含體積分數0.05的CO2無菌恒溫培養箱內培養至對數生長期且適宜密度時用于后續實驗。
1.4 研究方法
1.4.1細胞相對活力檢測 接種SH-SY5Y細胞至96孔板中,待細胞貼壁后,按照高糖DMEM培養基中加入的MPP+濃度不同分為A組(0 mmol/L)、B組(1.0 mmol/L)、C組(1.5 mmol/L),每組設5個復孔,每組細胞培養24、48、72 h時吸出原培養基,每孔分別再加入110 μL含有10 μL CCK8的高糖DMEM培養基,混勻,置于培養箱中繼續孵育2 h,采用酶標儀測定波長450 nm處的吸光值。
1.4.2細胞中ROS含量檢測 接種SH-SY5Y細胞至6孔板中,待細胞貼壁后,按照A、B組處理方式于高糖DMEM培養基中加入MPP+,培養24 h后吸出原培養基,用無血清基礎培養基洗滌2次,每孔加入1 mL DCFH-DA(10 μmol)探針,在細胞培養箱孵育30 min后,棄探針,無血清培養基清洗細胞3次,加入1 mL無血清培養基或PBS,熒光顯微鏡下觀察并拍照,實驗重復3次,實驗結果取均值。
1.4.3蛋白質印跡(Western blot)實驗檢測細胞中TMEM166及自噬相關蛋白質的表達水平 取生長狀態良好的SH-SY5Y細胞,以每孔2×105個接種于6孔板中。待細胞密度達到80%時,按照A~C組處理方式于高糖DMEM培養基中加入MPP+,培養24 h后,提取總蛋白。配制濃度為12%及5%的SDS-PAGE分離膠以及濃縮膠,等質量上樣,先80 V后110 V恒壓電泳,300 mA恒流濕轉法轉膜。轉膜完成后,取出PVDF膜放入50 g/L BSA的TBST溶液中室溫封閉2 h,取出膜于一抗中孵育,4 ℃冰箱過夜。TBST洗膜3次,每次10 min,接著室溫下于二抗中孵育1 h,TBST洗膜3次,在避光條件下于ECL顯影液中孵育,上機曝光,并使用Image J軟件分析目的條帶TMEM166、p62、LC3Ⅱ/Ⅰ和內參蛋白β-actin條帶的灰度值,以目的條帶與內參條帶灰度值比值表示相關蛋白的表達水平。
1.4.4實時熒光定量PCR(RT-qPCR)反應檢測細胞中TMEM166 mRNA水平 采用TRIZOL法分別提取A~C組SH-SY5Y細胞培養24 h后細胞中總RNA,測定RNA純度與濃度,引物名稱及序列見表1。以GAPDH為內參照,逆轉錄為cDNA并進行RT-qPCR。每個樣品分別設5個復孔,實驗重復3次,使用2-△△CT公式計算目的基因的相對表達水平。

表1 引物名稱及其序列
1.4.5細胞轉染及3-MA處理 取對數生長期的SH-SY5Y細胞,以每孔約2×105個細胞的密度接種于6孔板中,待細胞密度達到80%時,使用lipofectamine 2000按照說明書的用量分別轉染Myc-TMEM166質粒(D組)和Myc載體空質粒(E組),6 h后加入1.0 mmol/L MPP+處理,24 h后裂解細胞提取蛋白,按照1.4.3的方法檢測p62、LC3Ⅱ/Ⅰ和內參蛋白β-actin的表達水平。將SH-SY5Y細胞分為D、F 2組,D組按照上述的方法轉染Myc-TMEM166質粒,6 h后加入1.0 mmol/L 的MPP+處理24 h,F組轉染Myc-TMEM166質粒,6 h后加入1.0 mmol/L的MPP+和1.0 mmol/L的3-MA共同處理24 h,然后按照1.4.2 的方法檢測細胞中ROS的含量。
1.5 統計學方法
2 結 果
2.1 各組SH-SY5Y細胞的細胞相對活力及ROS含量比較
析因設計方差分析結果顯示,時間、組別及交互作用均對細胞相對活力有明顯影響(F組別=32.85,F時間=93.36,F時間*組別=7.60,P<0.05)。單獨效應結果顯示,B、C組細胞在不同時間的細胞相對活力比較有顯著差異(F=58.56、35.81,P<0.05),在同一時間點,各組細胞的細胞相對活力比較也有顯著差異(F=21.56~45.16,P<0.05),見表2。MPP+處理24 h后,A、B組細胞的ROS熒光強度分別為2.548±0.169、8.772±0.379,兩者比較有顯著差異(t=25.94,P<0.05)。
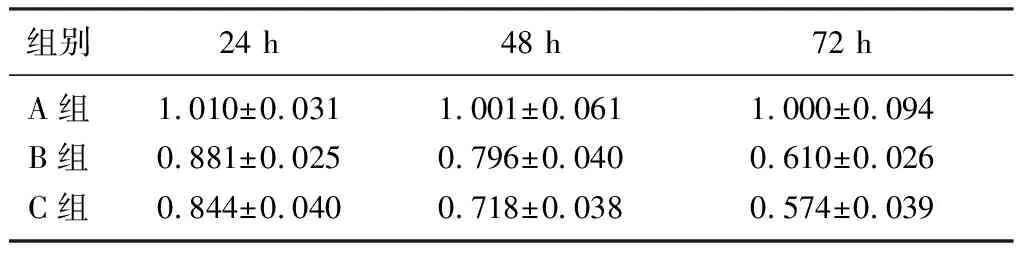
表2 不同時間各組SH-SY5Y細胞相對活力比較
2.2 各組SH-SY5Y細胞中TMEM166蛋白表達水平比較
A~C組SH-SY5Y細胞中TMEM166蛋白的相對表達水平分別為1.004±0.012、0.875±0.035、0.442±0.036,組間比較差異有顯著性(F=290.80,P<0.05),其中各組間比較差異均具有顯著性(t=5.29~23.02,P<0.05)。A~C組SH-SY5Y細胞中TMEM166 mRNA的表達水平分別為1.005±0.049、2.301±0.108、3.330±0.157,組間比較差異有顯著性(F=315.20,P<0.05),其中各組間比較差異均有顯著性(t=11.08~25.05,P<0.05)。
2.3 TMEM166過表達對MPP+誘導的SH-SY5Y細胞的相對活力和ROS含量的影響
D、E組SH-SY5Y細胞TMEM166蛋白表達水平分別為1.944±0.040、0.431±0.017,兩組比較差異有顯著性(t=3.96,P<0.05);D、E組細胞相對活力分別為0.627±0.023、1.003±0.016,兩組比較差異具有顯著性(t=4.77,P<0.001);D、E組細胞的ROS熒光強度分別為22.310±1.528、12.320±2.084,兩組比較差異有顯著性(t=6.69,P<0.05)。
2.4 過表達TMEM166對細胞自噬相關蛋白表達水平的影響
D組細胞中LC3Ⅱ/Ⅰ和p62蛋白的表達水平分別為0.772±0.049、0.634±0.071,E組分別為0.137±0.021、1.093±0.033,兩組上述兩種蛋白蛋白表達水平比較差異均具有顯著性(t=2.91、3.99,P<0.05)。
2.5 藥物抑制TMEM166導致的自噬對SH-SY5Y細胞中ROS含量的影響
D、F組細胞ROS熒光強度分別為19.580±1.824、13.280±2.106,兩組比較差異有顯著性(t=3.91,P<0.05)。
3 討 論
PD是第二大神經退行性疾病,目前沒有有效治愈方法,其治療耗費巨大,給患者家庭帶來沉重負擔。近年來對PD的深入研究發現,細胞自噬與PD的發生及發展密切相關[5,8],例如細胞自噬廣泛參與了PD致病蛋白α-突觸核蛋白(α-syn)的降解[9],SNCA、LRRK2和PINK1/Parkin等家族性PD相關突變基因的蛋白都參與到了自噬過程中[10-11]。一方面,在家族遺傳性的PD患者群體中,中腦黑質多巴胺能神經元中的自噬普遍受到阻滯,不能有效清除細胞內的錯誤折疊蛋白而逐漸變性死亡;另一方面,基于這些PD相關基因(Parkin,PINK1等)構建的動物模型中,細胞內線粒體質量不能得到有效的控制,導致細胞能量平衡破壞,多巴胺能神經元之間不能進行高效的細胞信號交流,最終造成PD或其他中樞神經系統相關的疾病[12]。基于PD患者神經元細胞自噬受阻的這一特點,探索其中的自噬調節機制,或許是治療該疾病的關鍵“鑰匙”。靶向自噬治療PD的理論已被提出并廣泛研究,然而一些新發現的自噬調節蛋白與PD的聯系還未知,探索這些自噬調節蛋白與PD的關系及其對PD靶向治療的價值具有重要意義。
TMEM166是一種新發現的促進自噬的內質網跨膜蛋白,經前期研究發現,TMEM166與自噬相關蛋白16L(ATG16L)存在相互作用,促進ATG12-ATG5/ATG16L復合物在自噬體膜上的招募,進而促進自噬小泡的形成[13]。TMEM166在腫瘤中的作用也備受關注,TMEM166在眾多的腫瘤組織中呈明顯的低表達,且其外源性的過表達會減弱腫瘤細胞的“癌性”,表明TMEM166是一種抑癌基因[14-20]。值得注意的是,TMEM166-/-小鼠的蛋白質組學研究提示,TMEM166極有可能在PD等神經退行性疾病中發揮作用[4]。基于此,本研究借助MPP+誘導的SH-SY5Y PD細胞模型,初步研究了TMEM166在PD發生和發展中的作用。首先,本研究經過篩選采用1 mmol/L濃度的MPP+處理神經母瘤SH-SY5Y細胞系24 h,產生可辨別的ROS模擬氧化應激狀態,并且細胞存活數量較多,適合后續實驗。其次,本研究發現,在MPP+處理過程中,TMEM166的mRNA表達水平升高,而其蛋白水平降低,表明TMEM166蛋白在MPP+處理過程中被降解;外源過表達TMEM166再經MPP+處理,會促進MPP+誘導的ROS產生,表明TMEM166過表達加重了SH-SY5Y細胞的氧化應激,不利于細胞存活,這與TMEM166對腫瘤細胞有抑制作用的觀點是一致的[14],因此細胞會自主地選擇將TMEM166降解。TMEM166在PD細胞模型中被降解以及其降解方式,之前還未見相關報道,鑒于已有研究的自噬相關跨膜蛋白LC3及GABARAP都是通過自噬途徑被降解的,而一般的跨膜蛋白是通過溶酶體降解,并且有研究鑒定出TMEM166蛋白存在溶酶體定位[17],因此推測TMEM166作為內質網跨膜蛋白,很可能通過自噬途徑被降解,但還需要進一步研究證實。本研究對TMEM166促進氧化應激的機制進一步探索發現,過表達TMEM166會造成自噬銜接蛋白p62的表達水平降低,以及代表噬體膜含量的LC3Ⅱ/Ⅰ蛋白水平的升高,均提示TMEM166促進了SH-SY5Y細胞的自噬,這與先前發現TMEM166能夠與ATG16L相互作用促進自噬小泡的形成,并正向調控自噬功能的研究結果相一致[13]。對SH-SY5Y細胞過表達TMEM166后,聯合使用自噬抑制劑3-MA和MPP+處理細胞,結果顯示藥物抑制自噬可以減少MPP+誘導的ROS產生,提示TEMEM166促進的自噬是MPP+誘導的SH-SY5Y細胞的氧化應激反應加重的原因之一。
在自噬參與的各種癌癥中,自噬往往扮演著雙重角色。一方面,自噬的減弱會增加異常功能蛋白質的聚集,蛋白質質量不能得到控制,導致細胞向癌變方向發展;另一方面,自噬的增強可以為腫瘤細胞的增殖提供能量和原料[21-22]。而在神經退行性疾病中,神經元中的自噬往往表現出受阻,導致α-syn、tau蛋白或亨廷頓致病蛋白的堆積,而目前已有通過靶向自噬治療神經退行性疾病的研究報道[23-25]。
綜上所述,本研究通過MPP+誘導SH-SY5Y PD細胞模型探究TMEM166對神經細胞氧化應激的影響和機制,發現TMEM166促進的自噬對神經細胞的生存是不利的,神經細胞會降解TMEM166以減弱MPP+的細胞毒性,TMEM166降解障礙可能是促進PD進展的潛在機制之一,這一新發現可以為PD的自噬靶向治療提供指導。
利益沖突聲明:所有作者聲明不存在利益沖突。
ConflictsofInterest:All authors disclose no relevant conflicts of interest.
作者貢獻:李寧、王喆、蒲明怡參與了研究設計;蒲明怡、王喆、廖肇忠、鞏遵雙、楊偉燕、華君男、李寧參與了論文的寫作和修改。所有作者均閱讀并同意發表該論文。
Contributions:The study was designed byLINing,WANGZhe, andPUMingyi.The manuscript was drafted and revised byPUMingyi,WANGZhe,LIAOZhaozhong,GONGZunshuang,YANGWeiyan,HUAJunnan, andLINing.All the authors have read the last version of the paper and consented submission.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
西南軍醫(2016年6期)2016-01-23 02:21:19
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
西南軍醫(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
