基于非靶向代謝組學(xué)研究砷酸鹽脅迫下庫德畢赤酵母代謝物的變化
2022-12-26 06:31:12戚曉雪徐瑩汪東風(fēng)
食品與發(fā)酵工業(yè) 2022年23期
關(guān)鍵詞:差異
戚曉雪,徐瑩,汪東風(fēng)
(中國海洋大學(xué) 食品科學(xué)與工程學(xué)院,山東 青島,266003)
隨著工業(yè)發(fā)展,砷被越來越多地釋放到環(huán)境中,進入食物鏈和生物圈,對人類、動物和環(huán)境造成嚴(yán)重威脅[1-3]。砷酸鹽[arsenate,As(Ⅴ)]是海水中主要的砷化合物,能被浮游植物、藻類、甲殼動物、軟體動物等海洋生物吸收[4],并通過多種方式進入食物鏈,影響食品安全。研究人員對昆山市農(nóng)貿(mào)市場及超市所售食品隨機抽樣,211份檢測食品中,砷的檢出率是21.33%;其中發(fā)現(xiàn)3份海魚樣本砷超標(biāo),最高超過國家標(biāo)準(zhǔn)近3倍[5]。因而急需建立一種安全、高效、適用范圍廣的食品砷脫除方法,安全經(jīng)濟的微生物脫除法已成為砷脫除的優(yōu)選方法之一。
一些關(guān)于酵母的研究顯示了它們在砷生物修復(fù)方面的潛力[6-8]。庫德畢赤酵母(Pichiakudriavzevii)作為一種多抗性酵母,可以抵抗高鹽、高溫和低pH值[9-10],已從果汁、漿果、茶啤酒和一些非洲發(fā)酵乳制品中被分離出來[11-14]。常被用來生產(chǎn)乳酸[10]、琥珀酸[15]和乙醇[16]等物質(zhì),在食品工業(yè)中有著廣泛的應(yīng)用前景。前期對P.kudriavzeviiA16的研究表明,其對鎘、鋅等多種重金屬離子具有良好的抗性和脫除能力[17-18],具有從食品中脫除有害物質(zhì)的應(yīng)用潛能。
非靶向代謝組學(xué)技術(shù)可以盡可能多地定性以及相對定量生物體系中的代謝物,最大程度來反映總代謝物信息,常用于代謝表型的區(qū)分和差異代謝物的發(fā)現(xiàn)[19]。代謝組學(xué)是更接近于表型的組學(xué),能夠更直接、更準(zhǔn)確地反映生物體的生理狀態(tài)。因此,非靶向代謝組學(xué)技術(shù)有利于更直觀地研究As(Ⅴ)脅迫下P.kudriavzeviiA16代謝物的變化。
本研究著眼于微生物脫砷技術(shù)在行業(yè)中廣闊的應(yīng)用前景,以P.kudriavzeviiA16為研究對象,基于非靶向代謝組學(xué)方法探討As(Ⅴ)暴露24 h后P.kudriavzeviiA16細胞中代謝物的變化及可能的通路分析,該研究為如何提高生物體的砷脫除能力提供有益思路,并更好地應(yīng)用于食品中砷的脫除。
1 材料與方法
1.1 試驗菌株
P.kudriavzeviiA16為本實驗室保藏菌株,分離自高溫白酒酒醅[20]。
1.2 試劑與儀器
1.2.1 主要試劑
Na2HAsO4·7H2O(分析純),湖北楚盛威化工有限公司;硝酸(優(yōu)級純);高氯酸(優(yōu)級純);砷標(biāo)準(zhǔn)液(GSB 04-1714—2004),國家有色金屬及電子材料分析測試中心。
1.2.2 主要儀器
QYC 211型恒溫振蕩培養(yǎng)箱,上海福瑪實驗設(shè)備有限公司;LD5-10型低速離心機,北京雷勃爾離心機有限公司;LDZX-50KBS型高壓蒸汽滅菌鍋,上海申安醫(yī)療器械廠;Powerwave XS型酶標(biāo)儀,美國BioTek儀器公司;EHD-24電熱消解儀,北京東航科儀儀器有限公司;8800電感耦合等離子體質(zhì)譜儀,安捷倫科技有限公司。
1.3 主要培養(yǎng)基
YPD培養(yǎng)基(g/L):20葡萄糖、20蛋白胨、10酵母浸粉。固體培養(yǎng)基添加20 g/L瓊脂。含砷培養(yǎng)基中砷的濃度根據(jù)實驗需要而定。
1.4 菌種活化
P.kudriavzeviiA16菌種先后進行固體活化及液體活化。首先將4 ℃保存的菌種接種到Y(jié)PD斜面固體培養(yǎng)基上,28 ℃恒溫培養(yǎng)24 h。從固體斜面上取少量菌體接種到50 mL YPD液體培養(yǎng)基中,在28 ℃恒溫振蕩(180 r/min)條件下培養(yǎng)24 h[20]。
1.5 As(Ⅴ)對P.kudriavzevii A16生長的影響和砷脫除率
取1 mL經(jīng)過液體活化后的P.kudriavzeviiA16菌懸液接種于含不同質(zhì)量濃度As(Ⅴ)(Na2HAsO4·7H2O 0、5、10、15、20 mg/L)的YPD培養(yǎng)基中,28 ℃、180 r/min恒溫培養(yǎng)24 h。取酵母培養(yǎng)液稀釋10倍后600 nm處測定吸光值,即用OD600來描述酵母菌生長情況。酵母培養(yǎng)液5 000 r/min離心5 min,取1 mL上清液加入到消化管中,并加入4 mL濃硝酸與1 mL高氯酸,加熱使其充分消化,超純水適當(dāng)稀釋后,使用電感耦合等離子體質(zhì)譜儀測定總砷含量,砷脫除率R按照公式(1)計算[20]。
(1)
式中:ρ0,生物積累前YPD培養(yǎng)基中總砷的質(zhì)量濃度,mg/L;ρt,生物積累后YPD培養(yǎng)基中總砷的質(zhì)量濃度,mg/L。
1.6 代謝組學(xué)分析樣品的制備
液體活化后的P.kudriavzeviiA16接種到Y(jié)PD培養(yǎng)基(空白對照組,BK組);液體活化后的P.kudriavzeviiA16接種到含12 mg/L As(Ⅴ) YPD培養(yǎng)基(砷脅迫組,As組);P.kudriavzeviiA16的接種量均為4×108個,28 ℃、180 r/min恒溫振蕩培養(yǎng)24 h。
采用離心法收集菌體(保證離心后各樣品菌體體積一致),用預(yù)冷的PBS快速沖洗2~3次,每次清洗后4 ℃、5 000 r/min離心5 min;棄去上清液,菌體收集在2 mL離心管中,液氮速凍15 min,-80 ℃保存。P.kudriavzeviiA16代謝物的提取及后續(xù)分析均委托北京Novogene生物信息科技有限公司完成。主要實驗流程圖如圖1所示。
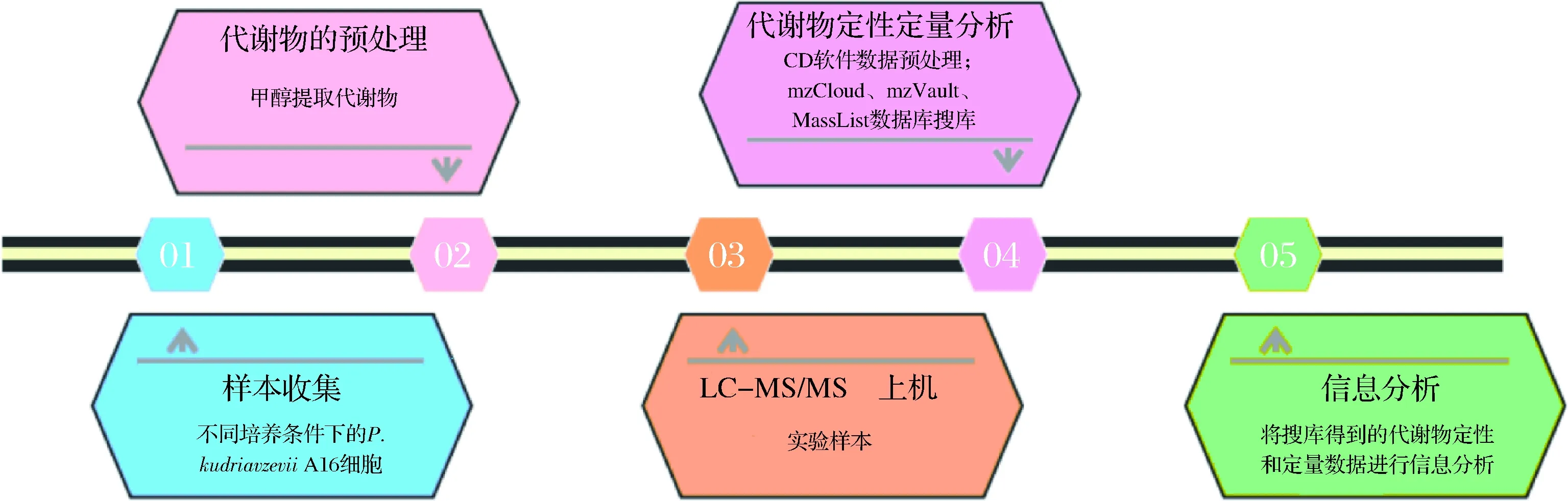
圖1 主要實驗流程圖Fig.1 Main experiment flow chart
1.7 數(shù)據(jù)處理及分析
使用KEGG(Kyoto Encyclopedia of Genes and Genomes)數(shù)據(jù)庫(https://www.genome.jp/kegg/pathway.html)對鑒定到的代謝物進行注釋。多元統(tǒng)計分析部分,使用代謝組學(xué)數(shù)據(jù)處理軟件metaX對數(shù)據(jù)進行轉(zhuǎn)換,再進行主成分分析(principal component analysis,PCA)和偏最小二乘法判別分析(partial least square discriminant analysis,PLS-DA),進而得到每個代謝物在PLS-DA模型中第一主成分的變量投影重要度(variable importance in projection,VIP)。單變量分析部分,基于t檢驗來計算各代謝物在兩組間統(tǒng)計學(xué)顯著性(P值),并計算代謝物在兩組間的差異倍數(shù)(fold change,F(xiàn)C)。差異代謝物的篩選設(shè)定閾值為VIP>1.0,F(xiàn)C>1.5或FC<0.667且P<0.05。
2 結(jié)果與分析
2.1 As(Ⅴ)對P.kudriavzevii A16生長及砷脫除率的影響
不同濃度As(Ⅴ)對P.kudriavzeviiA16毒性有較大差異,P.kudriavzeviiA16培養(yǎng)液OD600及砷脫除率如表1所示。
正常培養(yǎng)條件下,P.kudriavzeviiA16的OD600為0.606,而5 mg/L As(V)脅迫下P.kudriavzeviiA16的OD600降低到0.502,10 mg/L As(V)脅迫下P.kudriavzeviiA16的OD600僅有0.063。隨著砷質(zhì)量濃度升高,生物量降低很多,這說明As(V)抑制了P.kudriavzeviiA16的生長。在As(V)的質(zhì)量濃度為5 mg/L培養(yǎng)基中接種P.kudriavzeviiA16培養(yǎng)24 h后,該組砷的脫除率為61.84%。隨著As(V)質(zhì)量濃度的增加,培養(yǎng)液OD600值逐漸降低,同時各組砷脫除率也逐漸降低。P.kudriavzeviiA16的砷脫除率與P.kudriavzeviiA16的生長密切相關(guān)。

表1 不同質(zhì)量濃度As(Ⅴ)條件下P.kudriavzevii A16培養(yǎng)液OD600及砷脫除率Table 1 OD600 and arsenic removal rate of P.Kudriavzevii A16 culture under different concentrations of As(Ⅴ)
2.2 PCA
通過PCA圖可以觀察到不同處理組間的差異。如圖2所示,正負離子模式下As與BK組組內(nèi)均平行性較好,組間均相隔較遠,分離明顯,這表明每個不同區(qū)域中的樣本具有特定的代謝譜。與BK組相比,在As(Ⅴ)脅迫下的As組,P.kudriavzeviiA16產(chǎn)生了生物響應(yīng),代謝產(chǎn)物發(fā)生了明顯的變化。
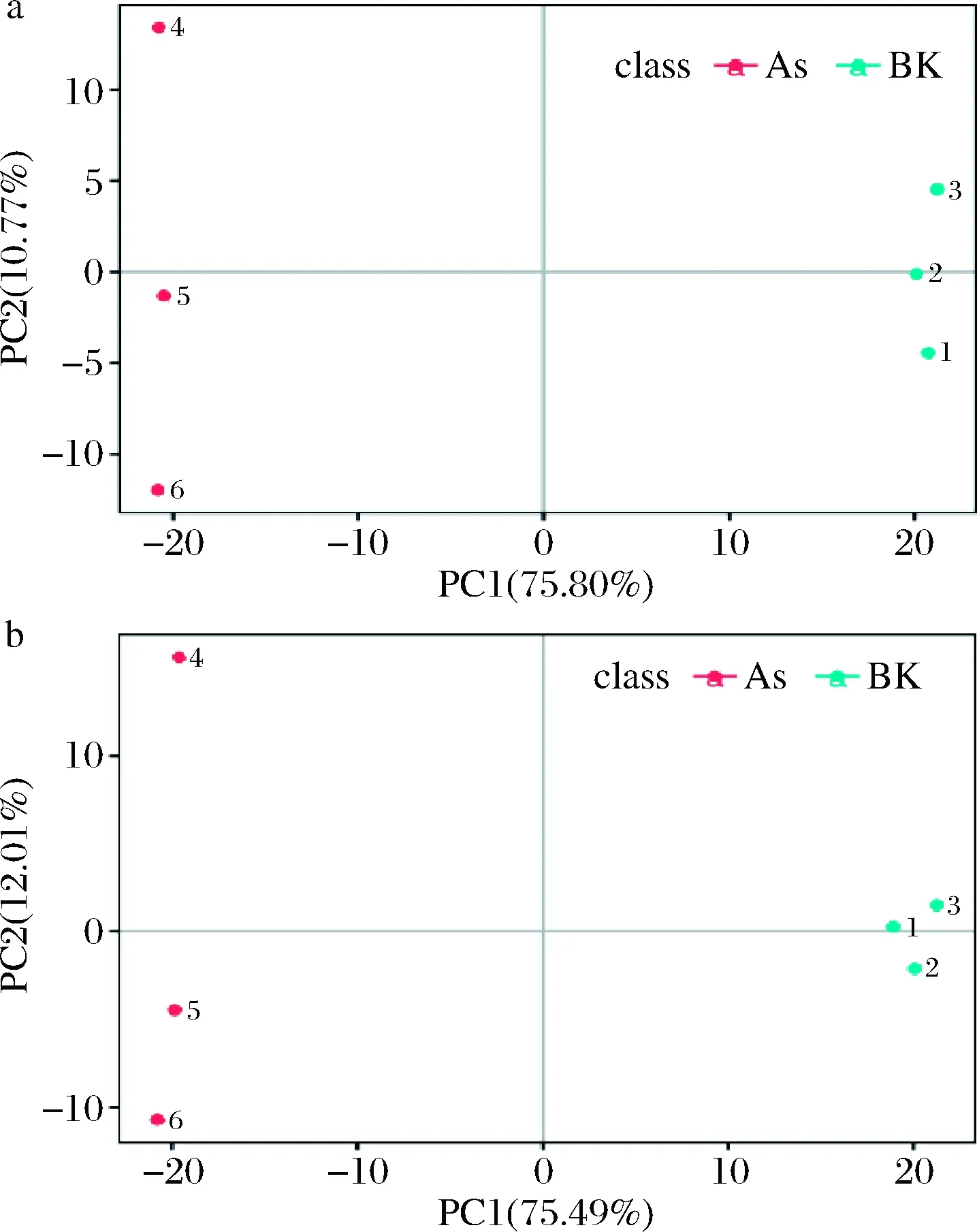
a-正離子模式;b-負離子模式圖2 主成分分析圖Fig.2 Principal component analysis
2.3 PLS-DA
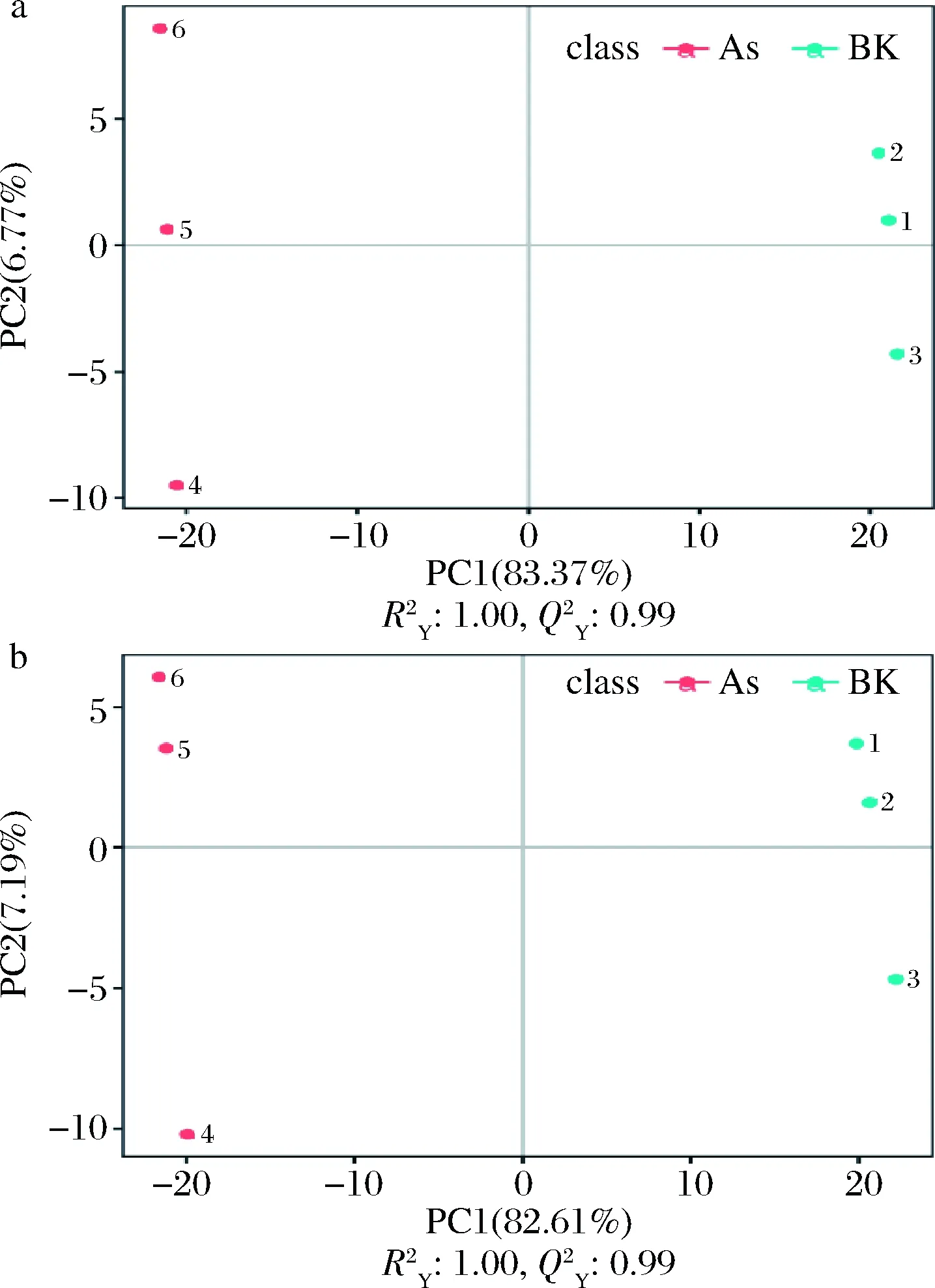
a-正離子模式;b-負離子模式圖3 PLS-DA得分散點圖Fig.3 PLS-DA dispersion point diagram
R2和Q2通常用來衡量模型是否過擬合。如圖4所示,正負離子模式下,As、BK組與POAs、As組,R2>Q2且Q2回歸線與Y軸截距<0,可知該模型未“過擬合”,穩(wěn)定性高。

a-正離子模式;b-負離子模式圖4 PLS-DA排序驗證圖Fig.4 PLS-DA sorting verification diagram
2.4 差異代謝物熱圖分析
不同樣品中代謝物的積累模式差異可以通過聚類熱圖進行分析,圖5顯示了正、負離子模式下As組與BK組的顯著性差異代謝物層次聚類結(jié)果。說明兩組樣本中差異代謝物可明顯區(qū)分,并且As(Ⅴ)脅迫P.kudriavzeviiA16下的代謝物與正常培養(yǎng)組BK組相比發(fā)生了顯著變化。
2.5 差異代謝物的篩選與鑒定
搜庫結(jié)果(mzCloud Results, mzVault Results,MassList Results)設(shè)定為full match,選擇差異代謝物TOP 35進行列表,如表2和表3所示。
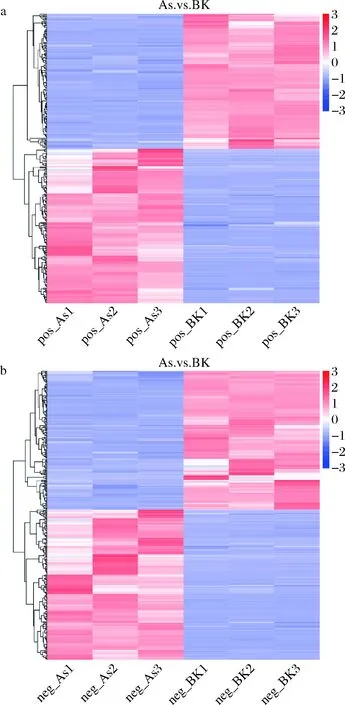
a-正離子模式;b-負離子模式;As1、As2、As3為As組中平行樣品;BK1、BK2、BK3為BK組中平行樣品圖5 差異代謝物聚類分析圖Fig.5 Cluster analysis of differential metabolites注:縱向是樣品的聚類,橫向是代謝物的聚類,聚類枝越短代表相似性越高

表2 正離子模式下As組與BK組差異代謝物Table 2 The significantly differential metabolites in the As group and BK group in positive ion modes
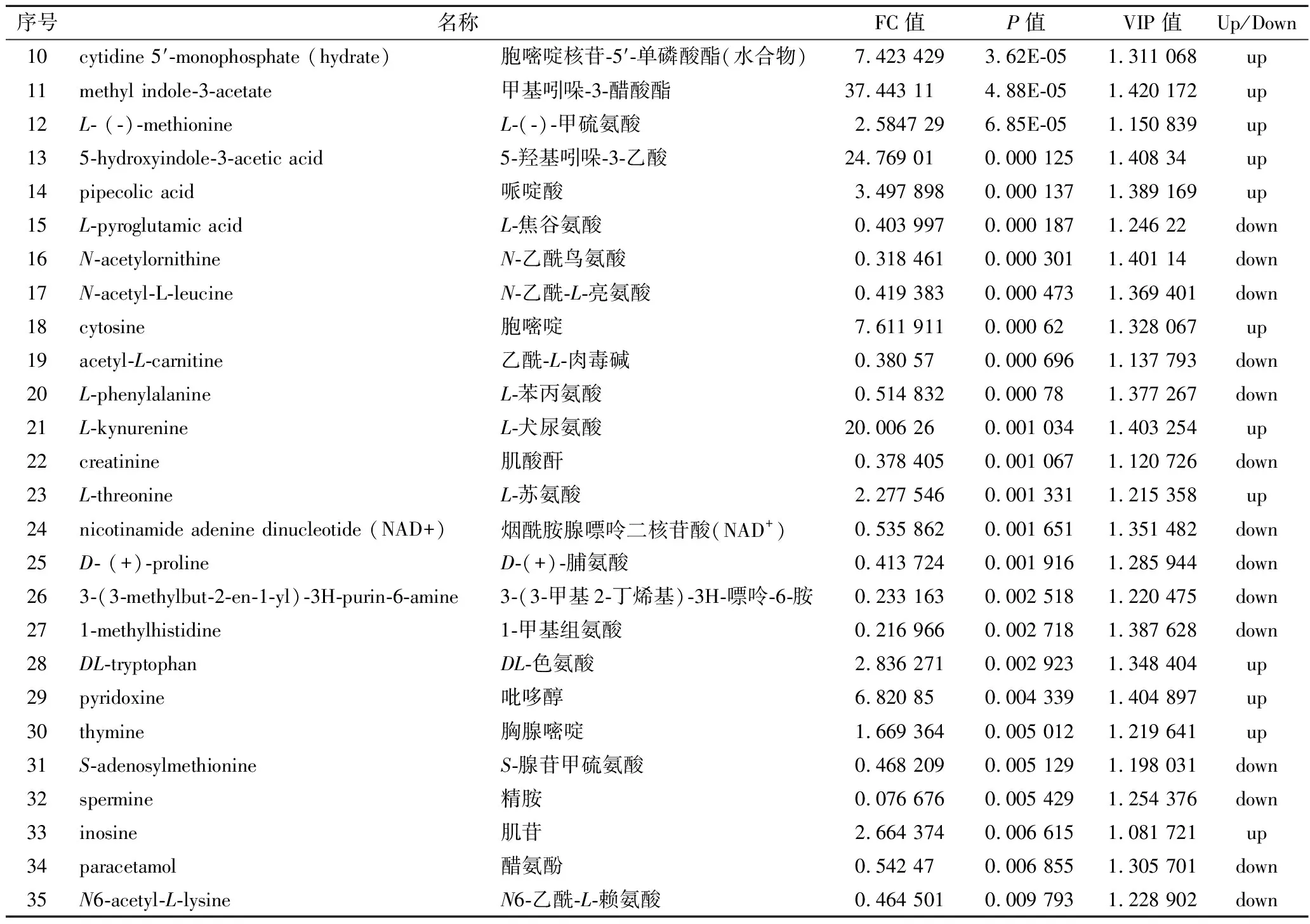
續(xù)表2
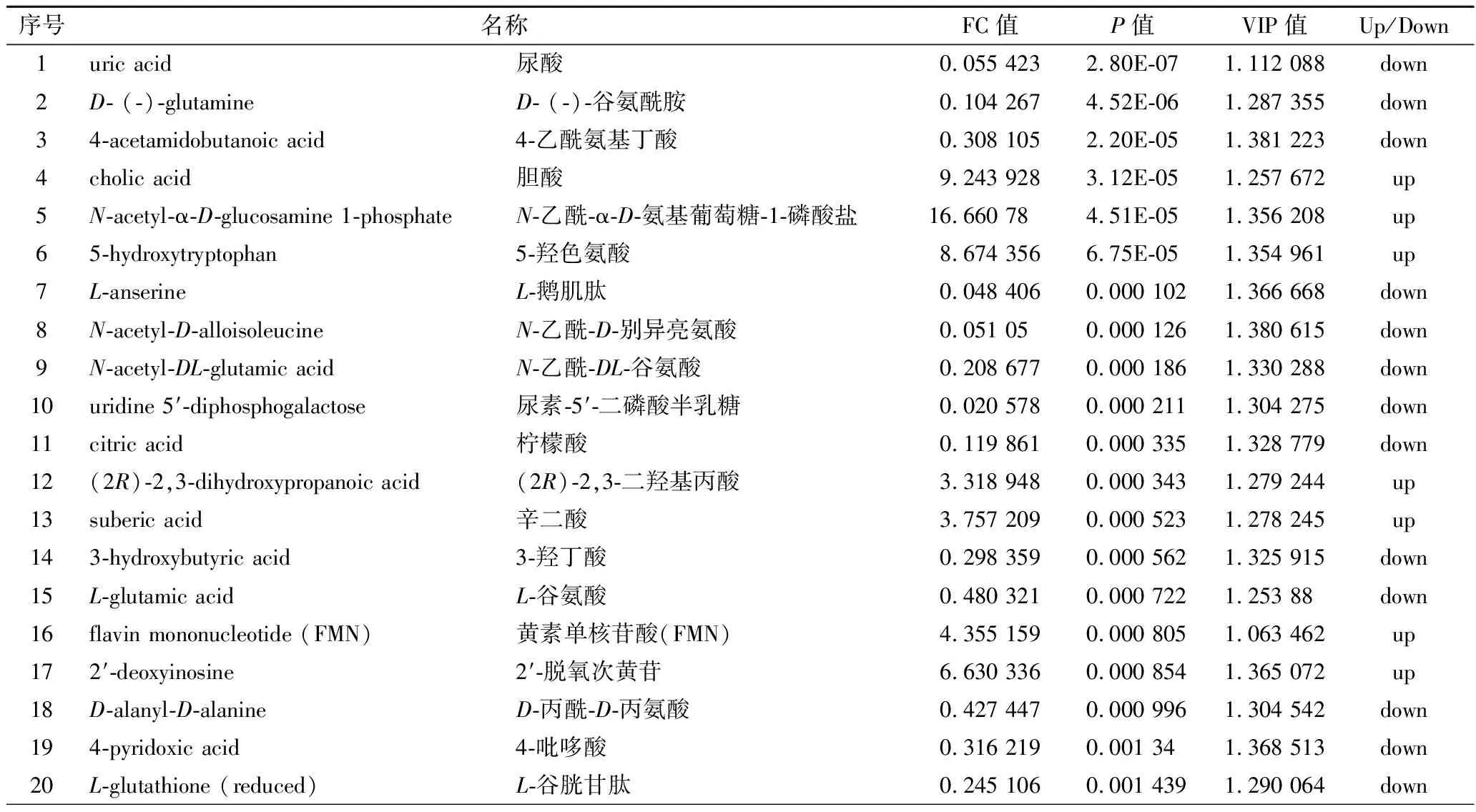
表3 負離子模式下As組與BK組差異代謝物Table 3 The significantly differential metabolites in the As group and BK group in negative ion modes
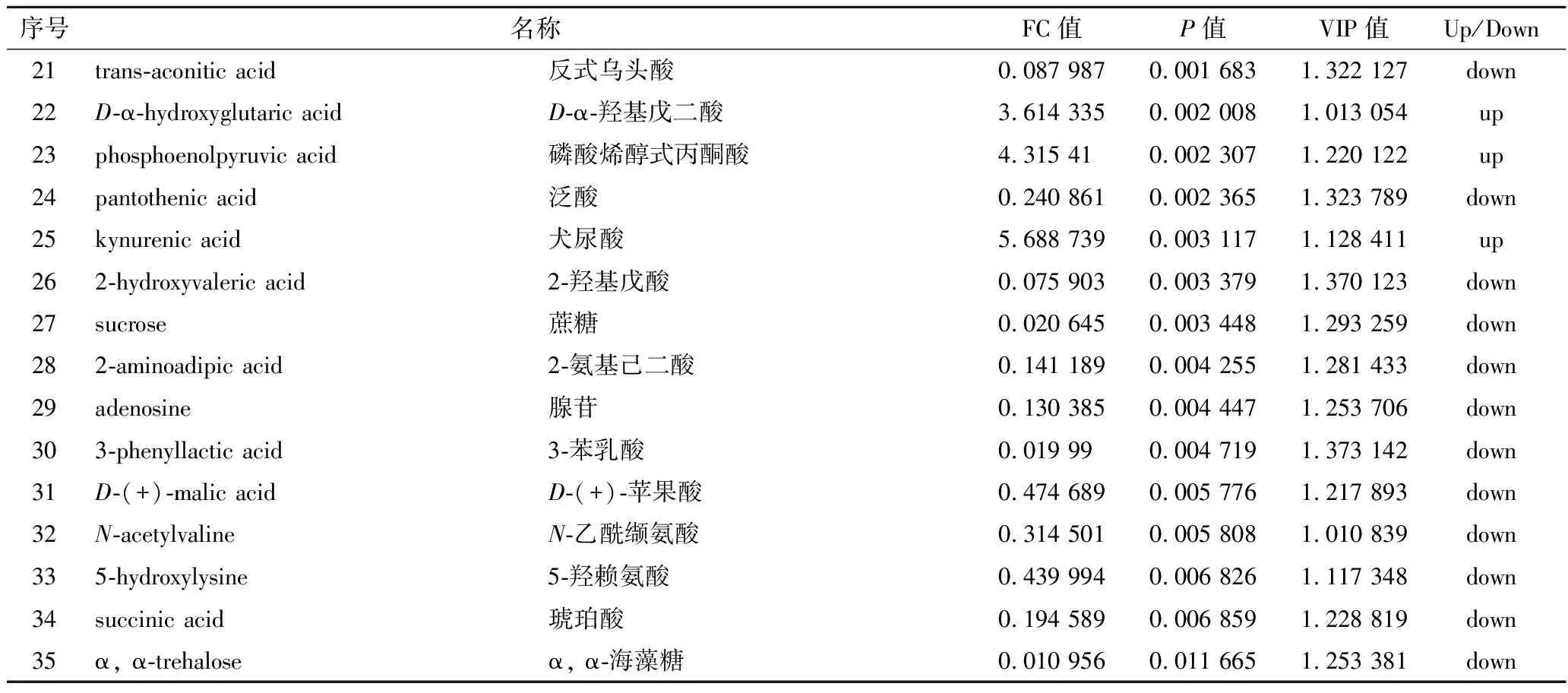
續(xù)表3
通過火山圖可以直觀顯示代謝物在兩組樣品中含量的差異,以及差異的統(tǒng)計學(xué)顯著性。火山圖中每個點代表一個代謝物,顯著上調(diào)的代謝物用紅色點表示,顯著下調(diào)的代謝物用綠色點表示,圓點的大小代表VIP值,如圖6所示。
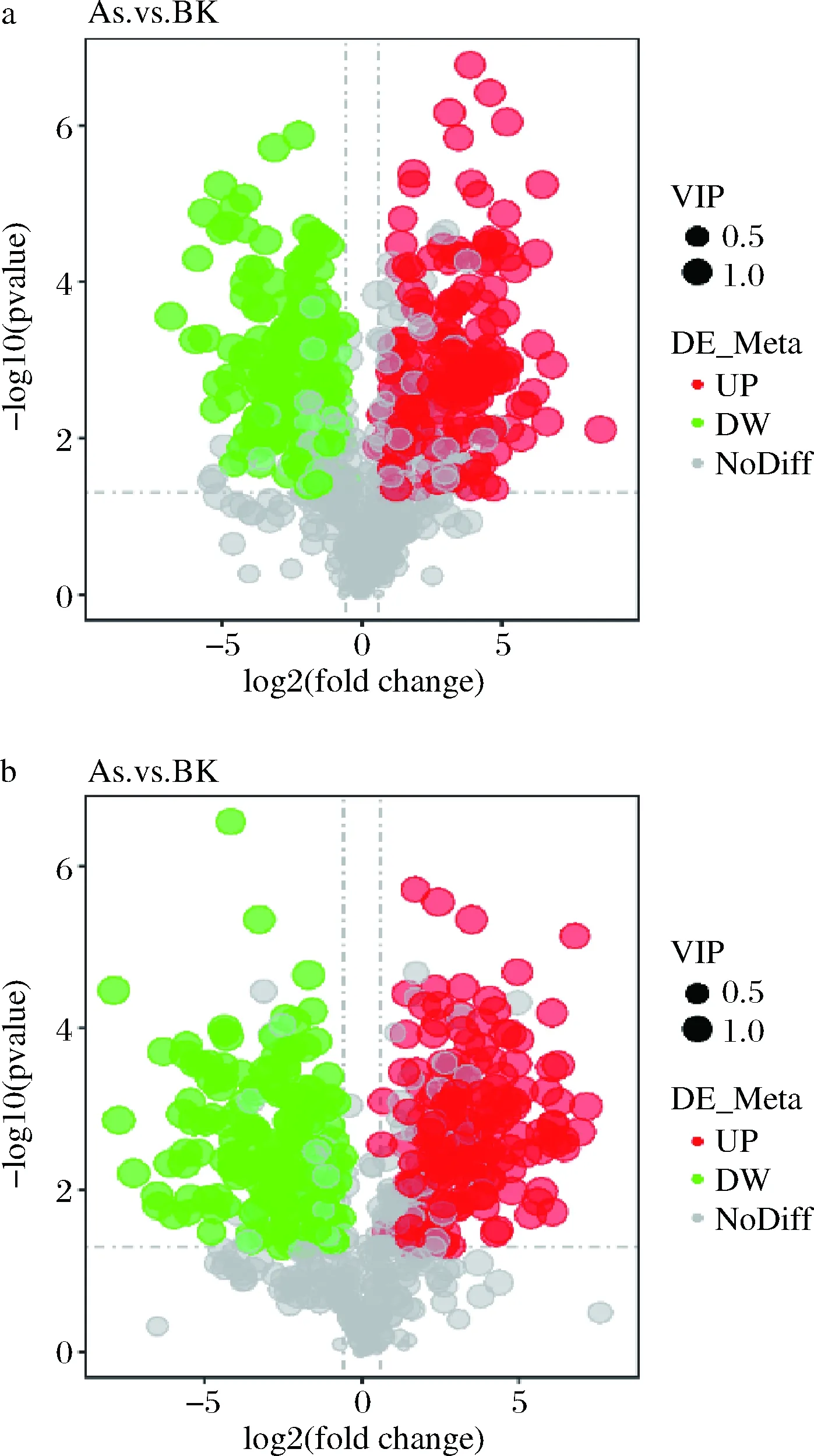
a-正離子模式;b-負離子模式圖6 差異代謝物火山圖Fig.6 Volcanic map of differential metabolites
正離子模式下,在As、BK兩組樣本之間共檢測到差異代謝物317種,其中,有169種顯著上調(diào)表達,其中包括鳥嘌呤、3’-磷酸腺苷、L-半胱氨酸、肌苷等,有148種代謝物顯著下調(diào)表達,包括DL-谷氨酰胺、煙酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD+)、D-(+)-脯氨酸、精胺等;負離子模式下,在As、BK兩組樣本之間檢測到差異代謝物329種,其中,有171種顯著上調(diào)表達,其中包括膽汁酸、黃素單核苷酸(flavin mononucleotide,F(xiàn)MN)、磷酸烯醇式丙酮酸等,有158種代謝物顯著下調(diào)表達,包括D-(-)-谷氨酰胺、檸檬酸、3-羥基丁酸、L-谷胱甘肽、琥珀酸、α, α-海藻糖等。
2.6 差異代謝物KEGG富集通路
將不同比較組中所有的差異代謝物匹配KEGG的數(shù)據(jù)庫從而獲得代謝物參與的通路信息。對注釋完的結(jié)果進行富集分析,獲得差異代謝物富集較多的通路。如圖7所示,正離子模式下,As組與BK組差異代謝物主要注釋和富集在谷胱甘肽代謝通路,氨酰tRNA生物合成代謝通路,甘氨酸、絲氨酸和蘇氨酸代謝通路、氰基氨基酸代謝通路等;負離子模式下As與BK差異代謝物主要注釋和富集在檸檬酸循環(huán),乙醛酸和二羧酸代謝通路,丙酮酸代謝通路,谷胱甘肽代謝通路,丙氨酸、天冬氨酸和谷氨酸代謝通路等。
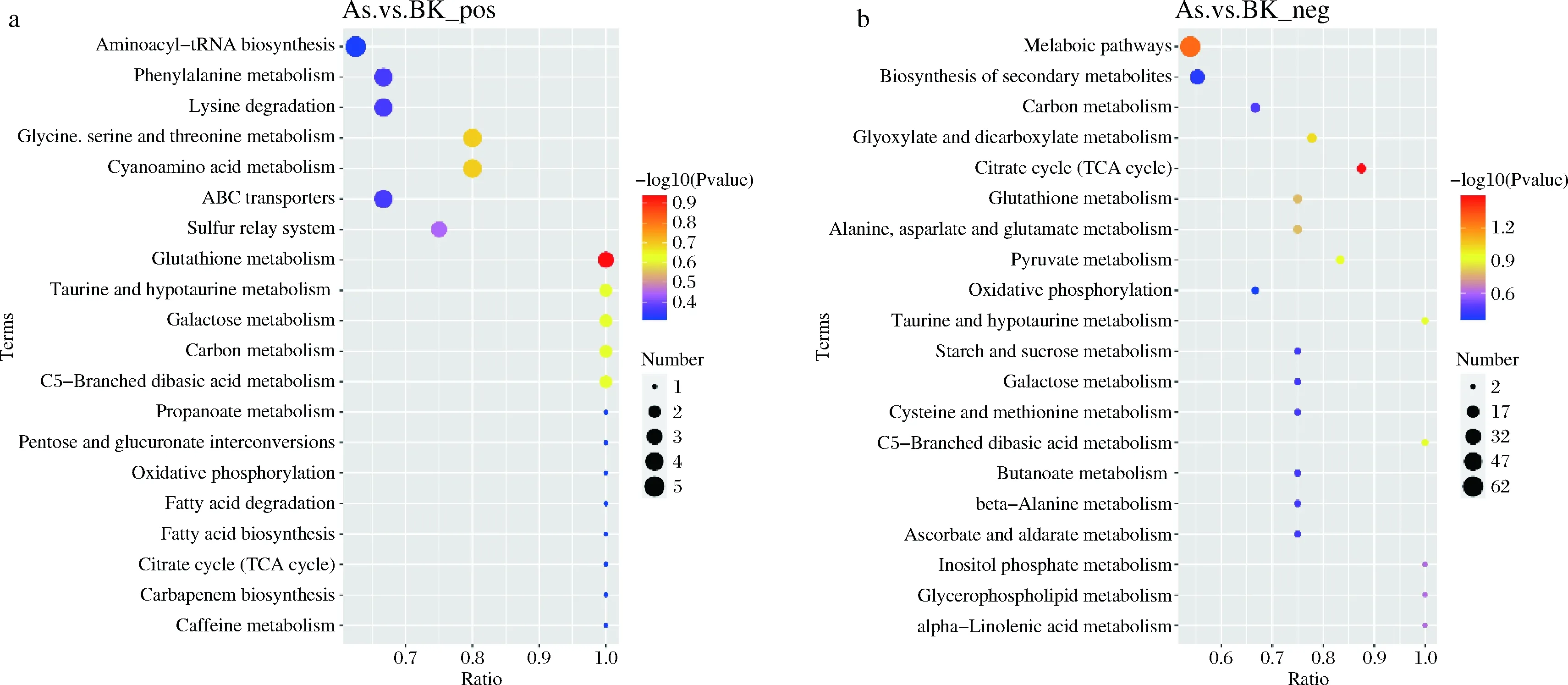
a-正離子模式;b-負離子模式圖7 KEGG富集通路氣泡圖Fig.7 KEGG enrichment pathway bubble map
3 討論
本研究采用先進的非靶向代謝組學(xué)技術(shù)對正常培養(yǎng)與砷酸鹽脅迫條件下P.kudriavzeviiA16的代謝物進行比對,并篩選出關(guān)鍵的差異代謝物,研究As(Ⅴ)脅迫下P.kudriavzeviiA16的代謝物變化。
磷酸烯醇式丙酮酸等代謝物在P.kudriavzeviiA16中的下調(diào)表達,說明As(Ⅴ)可能促進了P.kudriavzeviiA16的糖酵解反應(yīng);脂肪酸羥基化可生成羥基丁酸,而在3-羥基丁酸脫氫酶的作用下羥基丁酸氧化生成乙酰輔酶A,進入三羧酸循環(huán)(tricarboxylic acid cycle,TCA)[21]。As(Ⅴ)脅迫使得3-羥基丁酸等物質(zhì)的下調(diào)表明As(Ⅴ)可能會促進P.kudriavzeviiA16的脂肪酸代謝,上調(diào)TCA循環(huán),同時3′-磷酸腺苷的表達量上調(diào),可為細胞提供更多的能量供應(yīng)。
位于線粒體的內(nèi)膜上的電子傳遞鏈?zhǔn)茄趸姿峄磻?yīng)及ATP合成的主要位點,As(Ⅴ)脅迫下P.kudriavzeviiA16胞內(nèi)NAD+表達量的下調(diào)及FMN表達量的上調(diào),表明As(Ⅴ)可能促進了P.kudriavzeviiA16細胞的氧化磷酸化反應(yīng)。
氨基酸通過TCA循環(huán)可以為生物體供能[22],在本研究中,As(Ⅴ)脅迫下P.kudriavzeviiA16胞內(nèi)甘氨酸、絲氨酸和絲氨酸代謝通路,氰基氨基酸代謝通路,丙氨酸、天冬氨酸和谷氨酸代謝通路顯著上調(diào),說明As(Ⅴ)脅迫導(dǎo)致了酵母細胞的能量代謝發(fā)生紊亂。LI等[23]以甘氨酸等氨基酸的含量變化闡明了斑馬魚受到砷脅迫會導(dǎo)致其能量代謝異常。另外有研究表明,生物體可以通過提高氨基酸的代謝速率來為機體提供應(yīng)對外界脅迫所需的能量[24]。此外,氨酰基tRNA可以通過轉(zhuǎn)運氨基酸影響核糖體蛋白的生物合成,氨酰tRNA生物合成代謝通路顯著上調(diào)表達,表明As(Ⅴ)脅迫對P.kudriavzeviiA16胞內(nèi)的蛋白質(zhì)合成/降解產(chǎn)生干擾。這與當(dāng)砷質(zhì)量濃度大于5 mg/L時生長受到明顯抑制的現(xiàn)象一致。
谷胱甘肽(glutathione,GSH)是防御細胞氧化應(yīng)激關(guān)鍵因素,具有抗氧化的作用,可以緩解細胞的氧化應(yīng)激;除此之外GSH可與蛋白質(zhì)上的活性巰基結(jié)合,從而使其免于不可逆的氧化損傷[25]。GSH被認(rèn)為是細胞抵抗有毒物質(zhì)的一個重要組成部分,是無機砷氧化還原及砷解毒的重要物質(zhì)[22],作為一種金屬螯合劑,它在細胞抗砷毒性中發(fā)揮著重要作用[26]。谷胱甘肽代謝通路的顯著富集表明GSH可能被催化為氧化型谷胱甘肽,參與細胞砷的解毒。因此GSH與P.kudriavzeviiA16的生長和對培養(yǎng)基中砷脫除率關(guān)系密切,提示今后可以通過基因工程手段或者發(fā)酵工藝優(yōu)化來提高GSH的表達。
4 結(jié)論
砷酸鹽對P.kudriavzeviiA16的生長有抑制作用;隨著培養(yǎng)液中As(Ⅴ)濃度升高,P.kudriavzeviiA16對砷的脫除率下降。P.kudriavzeviiA16通過調(diào)節(jié)體內(nèi)糖酵解反應(yīng)、TCA循環(huán)、脂肪酸氧化代謝、氧化磷酸化和氨基酸代謝提高能量供給以應(yīng)對As(Ⅴ)脅迫導(dǎo)致的毒性效應(yīng);此外,谷胱甘肽代謝通路的顯著富集也表明P.kudriavzeviiA16胞內(nèi)GSH參與As(Ⅴ)的解毒作用,以減輕As(Ⅴ)脅迫帶來的細胞氧化損傷,降低As(Ⅴ)毒性。后續(xù)將利用其他組學(xué)聯(lián)合分析,厘清關(guān)鍵性差異代謝物并進行驗證性研究,為建立P.kudriavzeviiA16砷脫除技術(shù)提供有益參考。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
