當歸MADS-box生物信息學及SOC1克隆與表達分析
2023-03-10 01:48:26崔秀文黃天苗李美玲栗孟飛魏建和
中草藥 2023年5期
崔秀文,劉 迪,黃天苗,李美玲,栗孟飛*,魏建和
1.甘肅農(nóng)業(yè)大學生命科學技術學院 干旱生境作物學國家重點實驗室,甘肅 蘭州 730070
2.中國醫(yī)學科學院北京協(xié)和醫(yī)學院 藥用植物研究所,北京 100193
當歸Angelica sinensis(Oliv.)Diels 為傘形科多年生草本植物,干燥根是我國傳統(tǒng)中藥材,素有“十方九歸”之稱,具有補血活血、調經(jīng)止痛、潤腸通便等功效[1]。現(xiàn)代藥理學研究表明,當歸根中的有機酸類、揮發(fā)油類和多糖類等化學成分在消炎、抗癌和治療心腦血管疾病等方面具有顯著效果[2]。目前,當歸年需求量超3 萬t,年種植面積達4.35 萬hm2[3-4]。然而,當歸在第2年肉質根成藥過程中,出現(xiàn)高達50%植株早薹開花的現(xiàn)象,導致肉質根木質化,藥用有效成分含量降低且大量減產(chǎn),藥農(nóng)經(jīng)濟收入減少,影響當歸種質資源及產(chǎn)業(yè)發(fā)展[5-8]。
MADS-box 基因家族是一類廣泛存在于真核生物中的轉錄因子,在植物花器官分化、開花時間調節(jié)、以及果實發(fā)育等方面起到重要的調控作用[9];其通常分為I型和II型,其中,II型有4個保守結構域:M-domain(MADS-domain)、I-domain (intervening domain)、 K-domain(keratin-like domain)和 C-domain(C-terminal domain)[10]。目前,在模式植物擬南芥中發(fā)現(xiàn)了deficiens (DEF)、SQUAMOSA(SQUA)和TOMATO gene 3 (TM3-like)等12 個MADS-box亞家族[11]。前期在當歸轉錄組學的研究中發(fā)現(xiàn),MADS-box 亞家族基因,如SOC1(suppressor of overexpression of constans1)、FLC(flowering locus C)和AGL26(agamouslike 26)等,在抽薹開花過程中差異表達[12-16]。
研究證實,SOC1基因是花器官分生組織形成過程中的關鍵基因,其通過整合外界環(huán)境和內(nèi)在因素等各種成花途徑信號,激活下游花器官發(fā)育所需的基因,如LEAFY(LFY)、APETALA 1(AP1)和AGAMOUS(AG),進而促進植株開花[17]。前期當歸研究中發(fā)現(xiàn),SOC1基因在完成春化作用的種苗根莖頂端分生組織以及早薹植株中高表達[12-13],其上游基因[如COL16(constans-like)]和下游基因(AGL62)在抽薹開花過程中也呈現(xiàn)高表達[14]。綜合以上研究表明,盡管前人及本課題組已通過轉錄組測序獲得了當歸抽薹開花相關的MADS-box基因家族序列,并對關鍵基因(如SOC1、AG和VRN1)的表達水平進行了qRT-PCR 檢測與分析[12-16,18],然而,針對當歸MADS-box 基因家族以及關鍵基因SOC1的生物信息學分析等方面的系統(tǒng)研究尚未有報道。因此,本研究基于前期當歸全長轉錄組測序結果,開展了MADS-box基因家族生物信息學系統(tǒng)分析、SOC1-4基因克隆及表達驗證,旨在深入揭示MADS-box基因家族的生物學功能,為有效抑制抽薹開花提供理論基礎。
1 材料與儀器
1.1 材料
當歸MADS-box 編碼序列來源于:(1)不同品種(岷歸1 號和岷歸2 號)兩年生大田植株葉片和葉柄的全長轉錄組,結果已提交NCBI(Access:PRJNA782300),樣品采集等詳細信息見Zhu 等[18];(2)岷歸1 號種苗春化作用過程中[T1(0 ℃、14 d;未通過春化)、T2(0 ℃、60 d;通過春化)和T3(-3 ℃、125 d;低溫規(guī)避春化)]根莖頂端分生組織的全長轉錄組,結果已提交NCBI(Access:PRJNA789039),樣品采集等詳細信息見Luo 等[12],樣品原植物均由甘肅農(nóng)業(yè)大學栗孟飛教授鑒定為當歸A.sinensis(Oliv.)Diels。
1.2 儀器
臺式高速離心機(德國SORVAL 公司);ABIQuantStudio 5 實時熒光定量PCR 儀(美國ABI公司);超微量分光光度計(上海寶予德科學儀器有限公司)。
2 方法
2.1 MADS-box 家族鑒定、理化性質及結構分析
蛋白質序列(包括氨基酸長度、相對分子質量和理論等電點)分析利用 ExPASy(https://web.expasy.org/protparam/);亞細胞定位利用Cell-PLoc(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/);蛋白質二級結構預測利用 PRABI-Gerland(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?pa ge=/NPSA/npsa_sopma.html);蛋白質3D 建模利用Phyre 2 (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi)。
2.2 MADS-box 家族系統(tǒng)發(fā)育樹構建
在 NCBI 數(shù)據(jù)庫(https://www.ncbi.nlNm.nih.gov)選擇模式植物擬南芥Arabidopsis thalianaL.和傘形科植物黃胡蘿卜Daucus carotasubsp.sativus(Hoffm.)中與當歸29 個MADS-box基因家族置信度較高的116 個蛋白質,利用MEGA7.0 軟件最大似然法(maximum likelihood estimate,MLE)構建系統(tǒng)進化樹(重復次數(shù)1000 次,其他參數(shù)為默認值)。蛋白質保守基序分析利用 MEM E(https://memesuite.org/meme/tools/meme),并利用TBtools 進行可視化。利用DNAMAN 軟件進行蛋白質多序列比對(深藍色表示相似度100%、粉色>75%、淺藍色>50%)。
2.3 當歸SOC1-4 基因克隆
以岷歸1 號溫室栽培植株功能葉片為材料(“1.1”項中T2:0 ℃、60 d 通過春化,種苗移栽生長40 d),種苗種植及生長環(huán)境等詳細信息見Liu等[19]。總RNA 提取使用Plant RNA Kit R6827(Omega Bio-Tek,Norcross,GA,United States),其純度和濃度檢測使用超微量分光光度計(Micro Drop,上海寶予德科學儀器有限公司);反轉錄使用First-Strand cDNA Synthesis SuperMix(北京百泰克生物技術有限公司)得到cDNA;利用NCBIPrimer-BLAST(https://www.ncbi.nlm.nih.gov/tools/primer-blast/)設計SOC1-4基因引物序列,forward:5’-TGAGGGGAAAGACTCAGA-3’ 和 reverse :5’-CTGTTTCGACATCGGAAT-3’;擴增產(chǎn)物利用1%TAE 瓊脂糖凝膠電泳進行檢測;膠回收使用瓊脂糖凝膠純化試劑盒TIANgel Midi Purification Kit(天根生化科技有限公司),具體反應體系及條件見說明書。基因克隆使用平末端克隆試劑盒pHANDY?-Blunt Cloning Kit(哈爾濱曄健生物科技有限公司);引物合成及陽性克隆測序由蘭州天啟基因生物科技有限公司完成。
2.4 當歸SOC1-4 基因表達分析
基因表達材料:(1)岷歸1 號兩年生大田種植早薹(early bolting,EB)和非早薹(un-early bolting,Un-EB)植株葉和側根(混合1∶1),植株生長環(huán)境和樣品采集等詳細信息見Li 等[13];(2)岷歸1號三年生不同生長發(fā)育期(S1:營養(yǎng)生長期、S2:營養(yǎng)生長到生殖生長過渡期、S3:抽薹初期、S4:抽薹伸長期)植株葉和側根(混合1∶1),植株生長環(huán)境和樣品采集等詳細信息見Li 等[14]的實驗;(3)岷歸1 號不同春化期(T1、T2 和T3)種苗根莖頂端分生組織;(4)岷歸1 號溫室栽培植株(“1.1”項中T2:0 ℃,60 d 通過春化,種苗移栽生長60 d)不同器官(根、莖、葉)。
利用NCBI Primer-BLAST 設計SOC1-4基因qRT-PCR 表達引物,forward:5’-CGAAACGGCGAAATGGACTG-3’和reverse:5’-CTGAATGTCTTGCCCAGCAG-3’,引物合成由上海生工生物工程技術服務有限公司完成。以Actin作為內(nèi)參基因[20],設計引物序列,forward:5’-TGGTATTGTGCTGGATTCTGGT-3’和 reverse:5’-TGAGATCACCACCAGCAAGG-3’ ,利用 FastKing cDNA Kit(KR116,天根生化科技有限公司)合成cDNA;利用 SuperReal PreMix Plus(SYBR Green)進行qRT-PCR 檢測,具體反應體系及條件見說明書。利用2-△△Ct法計算SOC1-4基因的相對表達水平[21]。
2.5 統(tǒng)計分析
在qRT-PCR 實驗中,所采集材料進行了3 個生物學重復和3 個技術重復。運用Excel 2019 進行數(shù)據(jù)的計算及制圖。
3 結果與分析
3.1 當歸MADS-box 基因家族鑒定及理化性質分析
通過對當歸不同品種葉片和葉柄、種苗春化作用過程中根莖頂端分生組織的全長轉錄組進行篩選,發(fā)現(xiàn)有29 個MADS-box 基因,基于所編碼蛋白質序列和亞細胞定位分析,結果顯示,蛋白質序列長度為49(AP1-1)~422(At5g65490-1)、相對分子質量為5 697.56~49 624.90、等電點為5.06(J)~11.00(AGL14-1),亞細胞定位分析預測 29 個 MADS-box 蛋白分別位于葉綠體(AGL14-1、At3g28050-2和At5g65490-1)、細胞核(AGL14-2、AGL16和AGL19-1等22 個)、細胞質(AP1-1、AP1-2和At3g28050-1等7 個)和線粒體(SVP)(表1)。

表1 當歸MADS-box 基因家族蛋白質特征及亞細胞定位Table 1 Protein characteristic and subcellular location of MADS-box gene family in A.sinensis
3.2 當歸MADS-box 基因家族蛋白質結構分析
通過對29 個MADS-box基因所編碼的蛋白質進行二級結構預測以及3D 建模,結果顯示,蛋白質二級結構由α 螺旋、延伸鏈、β 轉角和無規(guī)則卷曲組成(表2);三級結構預測結果(圖1)與二級結構(表2)相符合,主要由α 螺旋、無規(guī)則卷曲和延伸鏈進一步折疊組裝為三級結構,各亞家族內(nèi)蛋白質三級結構較為相似,如AGL14-2、SOC1-1、SOC1-2、SOC1-3 和SOC1-4均以α 螺旋為主,無規(guī)則卷曲次之,β 轉角最少。但各亞家族之間空間結構差異較大,如AGL65、SOK2 及AP1-2 等以無規(guī)則卷曲為主、而AP1-1無延伸鏈(圖1)。
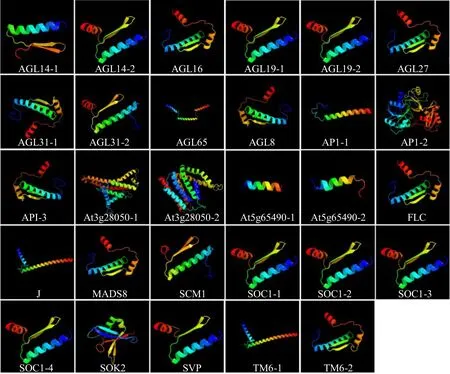
圖1 當歸MADS-box 基因家族蛋白質三級結構Fig.1 Tertiary structure of MADS-box gene family in A.sinensis

表2 當歸MADS-box 基因家族蛋白質二級結構Table 2 Secondary structure of MADS-box gene family in A.sinensis
3.3 當歸MADS-box 基因家族發(fā)育樹構建
通過對29 個當歸MADS-box基因家族蛋白質、61 個擬南芥和55 個黃胡蘿卜MADS-box 蛋白質進行系統(tǒng)進化樹構建,基于蛋白質序列相似性,145個MADS-box 蛋白質分為10 個亞家族,可分為I型和II型,其中,I型包括suppressor-like,II型包括SOC1、TM3、DEF、MADS8、SQUA、FLC、STMADS11、MIKC 和SOK;另外,SOC1 亞家族包括5 個當歸MADS-box 成員(SOC1-1、SOC1-2、SOC1-3、SOC1-4 和AGL14-2)(圖2)。

圖2 當歸、擬南芥和黃胡蘿卜MADS-box 蛋白質系統(tǒng)進化樹Fig.2 Phylogenetic tree of MADS-box family proteins in A.sinensis, A.thaliana and D.carota subsp.sativus
3.4 當歸MADS-box 基因家族基序分析
通過對以上29 個當歸MADS-box 基因家族蛋白質、61 個擬南芥和55 個黃胡蘿卜MADS-box 蛋白質進行結構域和基序分析,結果顯示,MADS-box蛋白質分為10 個亞家族,與圖2 系統(tǒng)進化樹關系一致,序列含有6 個保守基序(表3);當歸MADS-box同一亞家族蛋白質基序具有高度相似性,其中,SOC1 亞家族均含有Motif 1~Motif 5;不同亞家族蛋白質基序存在較大差異,比如,Motif 6 只存在于suppressor-like 亞家族(圖3)。
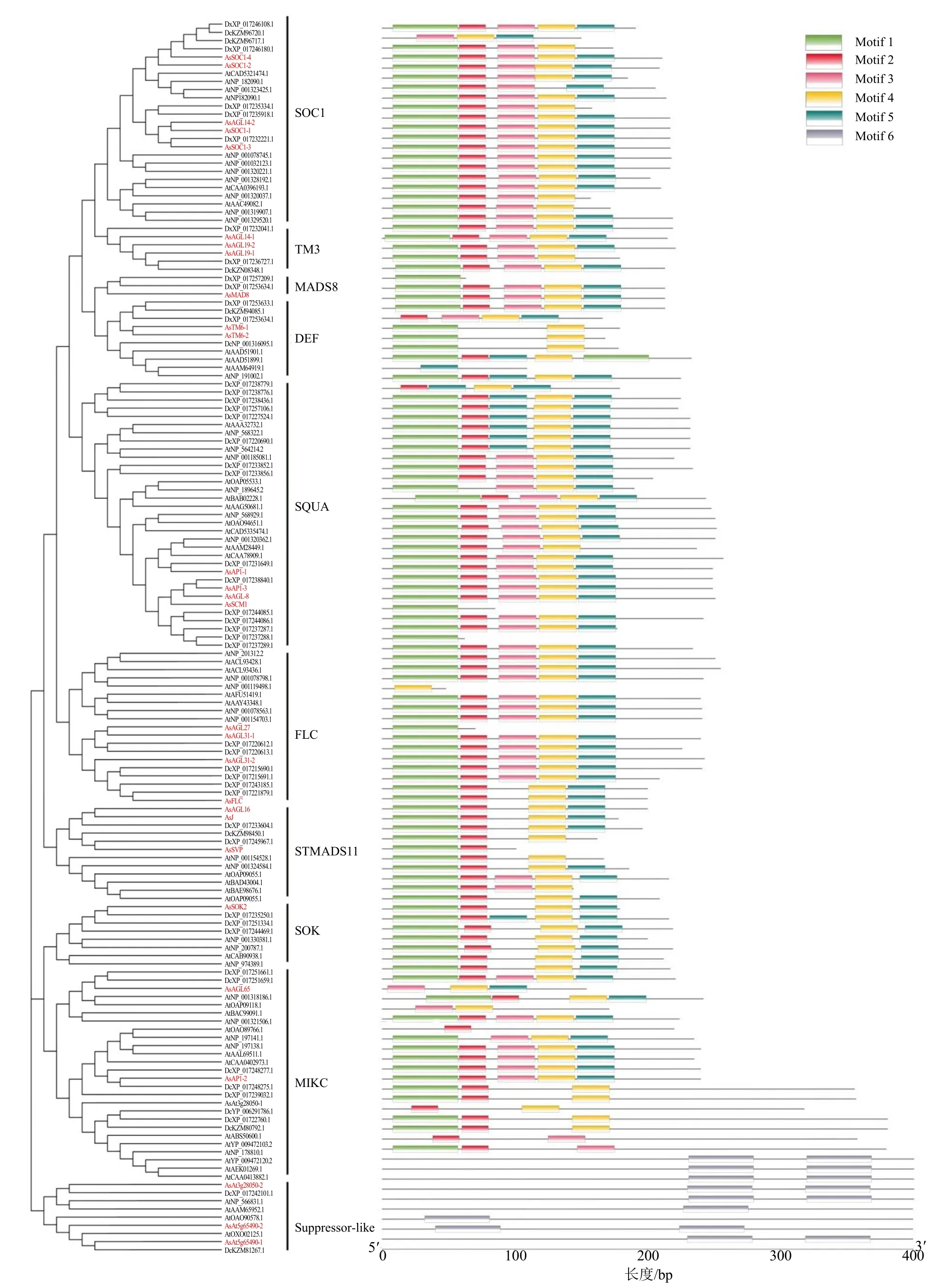
圖3 當歸、擬南芥和黃胡蘿卜MADS-box 蛋白質保守基序Fig.3 Conserved sequences of MADS-box proteins in A.sinensis, A.thaliana and D.carota subsp.sativus

表3 MADS-box 蛋白質6 個保守基序及其序列Table 3 Six motifs and their conserved sequences of MADS-box proteins
3.5 當歸SOC1 亞家族蛋白質多序列比對
通過對當歸SOC1 亞家族5 個成員(SOC1-1、SOC1-2、SOC1-3、SOC1-4 和AGL14-2)及來自擬南 芥 (AtNP_182090.1) 和 黃 胡 蘿 卜(DcXP_017232221.1)2 個物種的SOC1 同源蛋白質等7 個蛋白質進行多序列比對,結果顯示,SOC1亞家族5 個蛋白質均含MADS-domain、I-domain、K-domain 和C-domain 結構域,其中,MADS-domain結構域高度保守,C-domain 結構域保守性較低(圖4);通過對SOC1-1、SOC1-2、SOC1-3、SOC1-4進行進一步比對,發(fā)現(xiàn)SOC1-1、SOC1-3 和SOC1-4蛋白質的4 個結構域較為完整、且保守性較高,而SOC1-2 在C-domain 不完整;其中,SOC1-1、SOC1-3和SOC1-4 的相似性為73.27%。
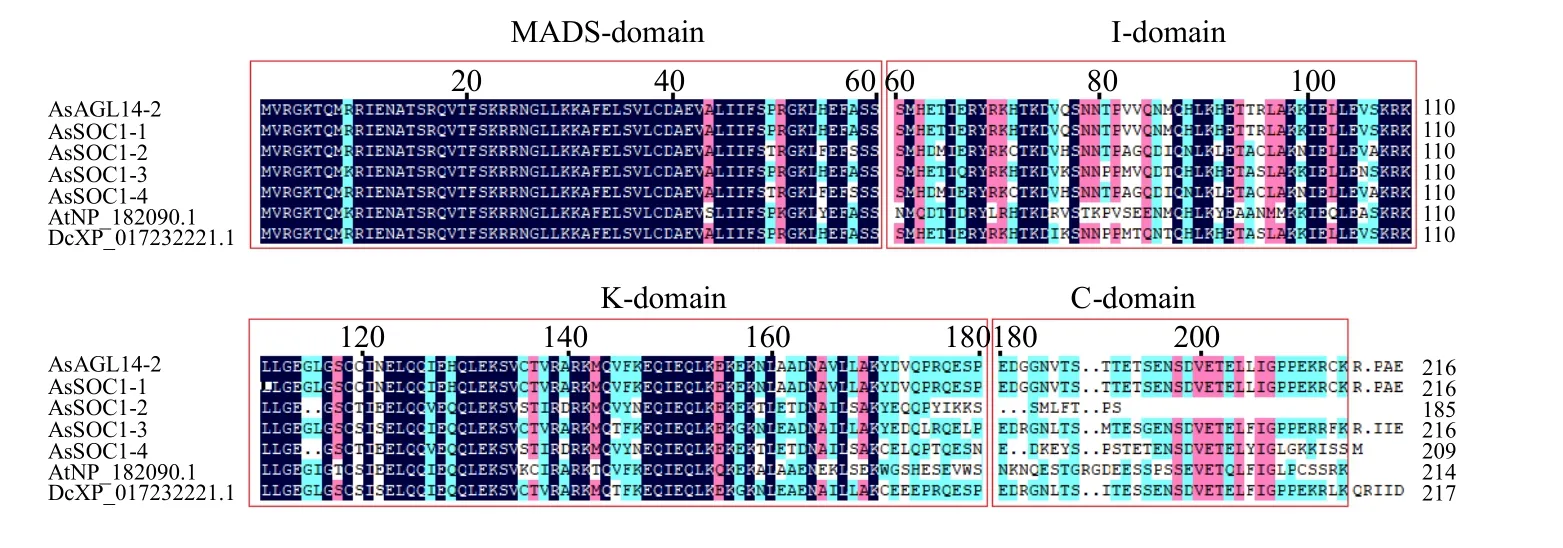
圖4 當歸SOC1 亞家族蛋白質多序列比對Fig.4 Multiple sequence alignment of SOC1 subfamily proteins in A.sinensis
3.6 當歸SOC1-4 基因克隆
為了保證全長轉錄組中當歸SOC1-4基因堿基序列的準確性,基于全長轉錄組中SOC1-4基因堿基序列設計擴增引物,以岷歸1 號功能葉片中RNA反轉錄所得的cDNA 為模板,進行SOC1-4基因克隆。瓊脂糖凝膠電泳顯示,SOC1-4基因擴增片段大小在500~750 bp(圖5);產(chǎn)物回收與堿基測序顯示,SOC1-4基因克隆長度為585 bp;通過與全長轉錄組測序獲得的SOC1-4進行序列比對,根據(jù)引物設計所得克隆結果與測序序列5-590 bp 相似度為100%(圖6)。
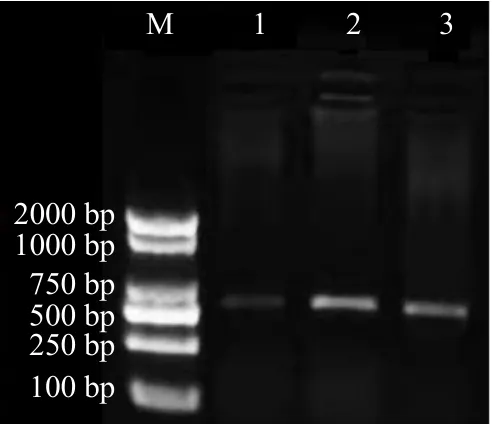
圖5 當歸SOC1-4 基因擴增產(chǎn)物瓊脂糖凝膠電泳Fig.5 Agarose gel electrophoresis of amplification products of SOC1-4 gene in Angelica sinensis

圖6 當歸SOC1-4 基因克隆與全長轉錄組測序的序列比對Fig.6 Sequence alignment of SOC1-4 gene obtained from gene clone and full-length sequencing in A.sinensis
3.7 當歸SOC1-4 基因表達分析
為了進一步驗證SOC1-4基因的生物學功能,本研究對當歸不同材料中SOC1-4基因的表達水平進行了qRT-PCR 檢測與分析。結果顯示,早薹植株相對非早薹植株,SOC1-4基因表達量上調6.42 倍;不同生長期植株中,SOC1-4基因表達水平隨著時間延長呈現(xiàn)逐漸增加,S2、S3 和S4 時期相對于S1時期分別增加2.16、2.31 和5.79 倍;不同春化期種苗根莖頂端分生組織中,通過春化作用(T2)處理相對于未春化作用(T1)SOC1-4基因表達水平呈現(xiàn)3.39 倍高表達,而規(guī)避春化作用(T3)處理相對于T1 處理SOC1-4基因表達水平呈現(xiàn)0.48 倍降低;不同器官中,莖和葉相對于根SOC1-4基因表達水平分別上調9.23 和4.90 倍(圖7)。
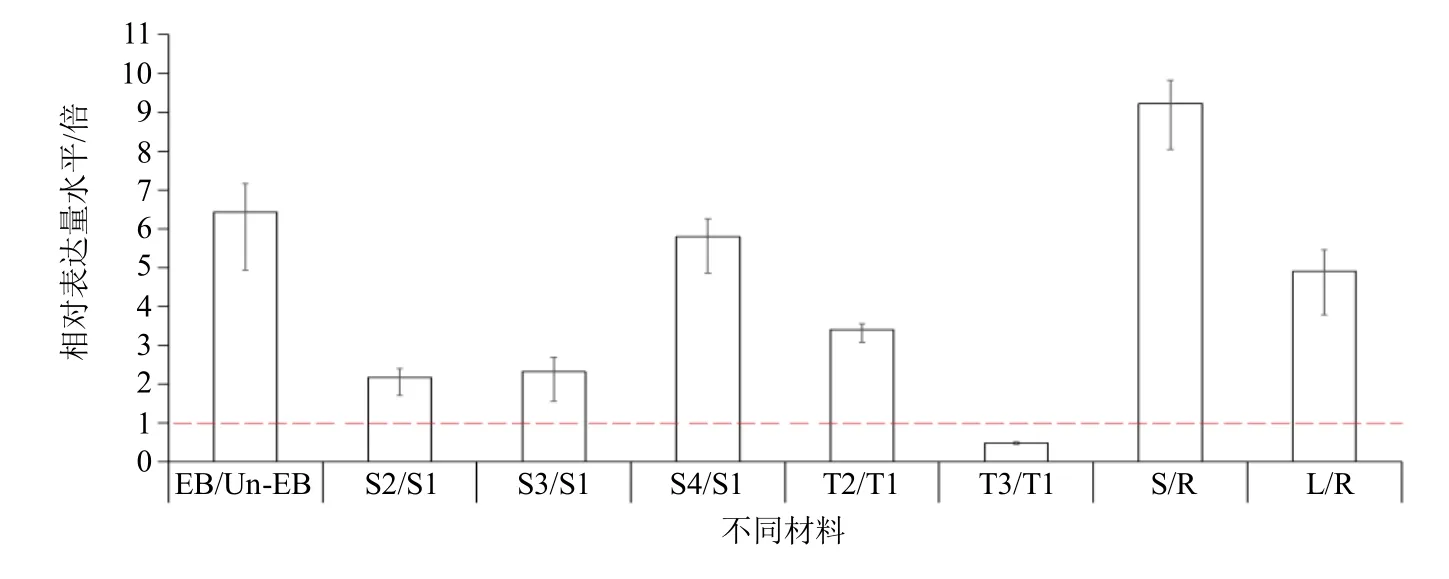
圖7 當歸SOC1-4 基因在不同材料中的相對表達水平Fig.7 Relative expression level of SOC1-4 gene in different materials of A.sinensis
4 討論
目前,早薹開花導致根木質化不能入藥仍是困擾當歸生產(chǎn)、質量和效益提升的重大難題[4]。研究發(fā)現(xiàn),當歸抽薹開花受到內(nèi)在(如種質、苗齡和種苗大小等)和外在(如溫度、光照和干旱等)多種因素的影響[4,22]。此外,當歸為“低溫長日照型”植物,即植株由營養(yǎng)生長轉入生殖生長必須同時滿足低溫春化作用和長日照[22-24]。大量研究表明,MADS-box基因家族,尤其是開花抑制因子FLC和整合因子SOC1,在植物花器官分化和開花時間調節(jié)等方面起到核心調節(jié)作用[17]。盡管MADS-box基因家族在模式植物擬南芥及其他植物中調節(jié)開花的作用已進行了深入研究,然而在藥用植物當歸中的生物學功能還鮮見報道。本研究基于前期當歸全長轉錄組測序,發(fā)現(xiàn)有29 個MADS-box基因分布于10 個亞家族,所編碼蛋白質序列含有6 個保守基序,其中,亞家族有5 個成員;另外,還對SOC1-4基因進行了克隆和表達分析,為后續(xù)探究SOC1 在當歸抽薹開花的分子調控研究奠定了基礎。
模式植物擬南芥中有DEF、STMADS11 和TM3-like 等12 個MADS-box 亞家族[11];山茶中有83 個MADS-box基因,可分為I型(Mα、Mβ 和Mγ亞家族)和II型(MIKCC和MIKC*亞家族)[25];荔枝中有101 個MADS-box基因,可分為I型和II型,其中,50 個I型分為Mα、Mβ 和Mγ 3 個亞家族,51 個II型進一步分為MIKC*、SOC1 和FLC等13 個亞家族[26]。本研究基于全長轉錄組鑒定出29 個MADS-box基因家族成員,基于蛋白質序列分為10 個亞家族,可分為I型(Suppressor-like)和II型(SOC1、TM3、DEF、MADS8、SQUA、FLC、STMADS11、MIKC 和SOK);其中,SOC1 亞家族包括 SOC1-1、SOC1-2、SOC1-3、SOC1-4 和AGL14-2。另外,前人預測22 種植物(擬南芥、水稻、煙草等)的SOC1 二級結構與亞細胞定位[27],與本研究MADS-box 蛋白質理化性質和結構分析,尤其是SOC1 亞家族等的結果基本一致,表明表明SOC1基因在進化上較為保守,在植物的生殖發(fā)育過程起著重要作用。
植物中MADS-box 的I型和II型蛋白質結構域各具有高度相似性,其中I型只含有一個保守結構域,而II型均具有M-domain、I-domain、K-domain和C-domain 結構域,推測當歸MADS-box 基因功能存在多樣性[7,11,28-30]。蛋白質保守基序比對發(fā)現(xiàn)不同亞家族保守基序存在數(shù)目及位置差異,預測不同亞家族功能有所差異。本研究中,SOC1 亞家族均含有II型的4個結構域,通過對5個成員SOC1-1、SOC1-2、SOC1-3、SOC1-4 和AGL14-2 進行多序列比對,發(fā)現(xiàn)MADS-domain 結構域高度保守,而C-domain 結構域保守性較低。這表明,5 個SOC1亞家族成員在當歸抽薹開花過程中可能發(fā)揮相似或協(xié)同的功能。另外,SOC1-1、SOC1-3 和SOC1-4蛋白質的4 個結構域較為完整,而SOC1-2 在C-domain 不完整,而樊世婷等[31]克隆的當歸SOC1基因(XM_017379845.1)與本研究中4 個SOC1基因比對存在差異,可能由于當歸無參考基因組序列,轉錄組序列拼接參考不同物種而產(chǎn)生的差異;或全長轉錄組樣本來自不同品種及材料所致。
盡管前期研究已獲得了當歸全長轉錄組,為了保證SOC1基因堿基序列的準確性、以及更深入探究SOC1基因的生物學特性,本研究通過克隆當歸SOC1-4基因,獲得了585 bp 片段,與全長轉錄組測序中SOC1-4基因序列 5-590 bp 結果完全一致,大量研究證實,作為開花整合因子SOC1,在植物花器官分化和開花時間調控過程中高表達[17,32]。本研究發(fā)現(xiàn),SOC1基因表達量隨著種苗春化作用和植株生長發(fā)育時期延長隨之增加、在抽薹植株中高于非抽薹植株、而在冷凍規(guī)避春化作用種苗中顯著降低。
綜合以上研究表明,本課題組首次對當歸MADS-box基因家族進行了挖掘和生物信息學分析,并對開花整合因子SOC1基因進行了基因克隆與表達驗證。但對于SOC1基因片段的完整性、以及其它調控當歸抽薹開花的關鍵基因(如FLC、AG和AP1等)還需要進一步研究和驗證。
利益沖突所有作者均聲明不存在利益沖突
