藏藥多刺綠絨蒿MYB轉錄因子家族的鑒定與分析
2023-03-10 01:48:32郭佳妮向妮艷張文才陳忠海丁亞麗
中草藥 2023年5期
郭佳妮,向妮艷,張文才,陳忠海,劉 星,3*,丁亞麗
1.西藏大學理學院 青藏高原生態與環境研究中心極端環境生物適應與進化實驗室,西藏 拉薩 850000
2.西藏大學醫學院,西藏 拉薩 850000
3.武漢大學生命科學學院,湖北 武漢 430072
轉錄因子是一類重要的基因表達調控因子,通常具有DNA 結合結構域、反轉錄激活結構域、核定位信號和寡聚位點,能夠調控特定基因的轉錄與表達。在轉錄因子家族中,MYB 轉錄因子數量眾多、功能多樣,是植物中最重要的轉錄因子家族之一[1],MYB 轉錄因子家族具有高度保守的DNA 結合結構域,該保守結構域由1~4 個串聯且不完全重復的R 基序組成,每一個R 基序編碼50~53 個氨基酸殘基,經過折疊形成HTH 的螺旋-轉角-螺旋結構[2]。MYB 轉錄因子依據其N 端含有的特有保守基序的數量劃分為 1R-MYB(MYB-related)、
2R-MYB(R2R3-MYB)、3R-MYB(R1R2R3-MYB)和4R-MYB 4 個亞家族[3],其中2R-MYB 是植物中最常見和目前功能研究最多的亞家族[4-5]。MYB轉錄因子作為調控生長發育、次級代謝物和脅迫響應的關鍵調控因子[6],在植物應對高鹽、干旱、極端溫度和營養缺乏等非生物脅迫方面具有重要意義[7-8]。目前,MYB 轉錄因子在植物中得到了廣泛研究,Cao 等[9]過表達蘋果MdoMYB121 基因顯著增強了轉基因番茄和蘋果植株對高鹽、干旱和低溫脅迫的耐受性;Cao 等[10]通過外源過表達楊梅MrMYB12 基因增加了煙草中黃酮醇的積累,從而減少了花青素的合成,產生淡粉色和純白色的花;Zhang 等[11]發現在脅迫環境下,擬南芥AtMYB49基因可以調節葉片角質層的沉積,從而調控植株的耐鹽性;Vannini 等[12]將水稻OsMYB4 基因在擬南芥中過表達顯著的提高了轉基因植株的耐寒性和抗凍性;Feng 等[13]發現擬南芥AtMYB68 基因在根中柱鞘細胞中的特異表達能參與調節木質素的生成,影響植株根系發育。
多刺綠絨蒿Meconopsis horridulaHook.f.et Thoms.為罌粟科綠絨蒿屬一年生草本植物,葉全部基生,葉片披針形,花芽近球形,主根肥厚而延長,生長于海拔3600~5100 m 以上的草甸、高山和流石灘地帶,在中國主要分布于西藏、甘肅西部、青海東部至南部和四川西部等高原地區。多刺綠絨蒿作為傳統藏藥,具有清熱止痛、活血化瘀等功效,被藏醫廣泛用于治療頭傷、骨折和跌打損傷等疾病[14]。現代藥理學研究表明,多刺綠絨蒿在抗病毒、抗腫瘤及心肌保護方面也有重要療效[15-16]。李長山等[17]對多刺綠絨蒿乙醇提取物進行抗病毒活性測試,結果顯示其提取物可以抑制甲型流感病毒。郭志琴[18]發現多刺綠絨蒿乙醇提取物對心肌缺血導致的血脂紊亂和心功能下降具有一定改善作用。多刺綠絨蒿具有豐富的次級代謝產物,目前從多刺綠絨蒿中分離得到黃酮類、萜類、生物堿類等化合物,其中黃酮類物質作為重要的藥效物質,其含量與生長環境密切相關[19-22],在高原環境下黃酮類物質的積累,有助于植物抵御強輻射所造成的氧化損傷[23-24]。近年來多刺綠絨蒿的研究大多集中在化學成分鑒定及臨床醫藥方面,對其如何應對高原極端環境的適應性進化及次級代謝物的調控機制卻鮮有報道[25]。
本研究基于轉錄組數據鑒定了多刺綠絨蒿MYB轉錄因子家族成員,對其理化性質、二級結構、保守基序、系統進化關系和表達模式進行分析,并預測了部分多刺綠絨蒿MYB轉錄因子的功能,篩選出可能參與黃酮類代謝及非生物脅迫等方面的MYB 轉錄因子,為進一步揭示和深入研究MYB 轉錄因子在多刺綠絨蒿的生長發育、次生代謝及脅迫響應機制等方面提供理論依據和基礎數據,為藏藥多刺綠絨蒿的可持續開發利用、生物多樣性保護及環境適應性機制的研究提供科學依據和數據支撐。
1 材料
多刺綠絨蒿實驗材料采集于西藏自治區拉薩市達瓊村(89°19?57″E,30°42?97″N,4925 m)、東德措(92°94?52″E,30°99?18″N,4872 m)和奪底溝(91°11'16″E,29°43'33″N,3980 m)3 個不同海拔地點,每個地點3 個生物學重復,共采集9 份葉片組織樣品,液氮速凍后,存放于超低溫冰箱(-80 ℃);進行轉錄組測序,共獲得60.15 Gb 轉錄組數據。樣品由武漢大學生命科學學院劉星教授鑒定多刺綠絨蒿M.horridulaHook.f.et Thoms.植株,標本現保存于西藏大學青藏高原生態與環境研究中心極端環境生物適應與進化實驗室。
2 方法
2.1 多刺綠絨蒿MYB 家族鑒定及理化性質分析
在Pfam(http://pfam.xfam.org)網站中下載MYB轉錄因子的隱馬爾可夫模型(PF00249),利用HMMER3.0 軟件中hmmsearch 程序進行鑒定;下載擬南芥已發表MYB 轉錄因子家族成員作為種子文件,利用Blast 軟件進行blastp 比對;將hmmsearch與blastp 2 種鑒定結果進行合并取交集作為候選基因提交至SMART(https://smart.embl.de/)、NCBI-CDD(https://www.ncbi.nlm.nih.gov/)、Pfam.search(http://pfam.xfam.org/) 和 PROSITE (https://prosite.expasy.org/)數據庫進行保守結構域的驗證。
通過ExPASy(https://web.expasy.org/)網站對MYB 轉錄因子的基本理化性質進行預測;利用CELLO[CELLO:Subcellular Localization Predictive System]網站(nctu.edu.tw)進行亞細胞定位分析;利用 SOPMA[NPS@:SOPMA secondary structure prediction]網站(ibcp.fr)對蛋白質進行二級結構預測。
2.2 多刺綠絨蒿MYB轉錄因子基因本體(gene ontology,GO)GO 功能注釋
利用 Blast2GO (Basic 6.0 https://www.blast2go.com/)軟件對多刺綠絨蒿MYB轉錄因子進行功能預測,利用軟件的Blast 工具將多刺綠絨蒿MYB轉錄因子蛋白序列與非冗余(nr)蛋白數據庫進行比對,并對每個MYB 轉錄因子進行GO 注釋,使用R 軟件(version 4.2.1)繪制GO 注釋分類圖。
2.3 多刺綠絨蒿MYB轉錄因子保守基序分析
利用MEME(MEME,V.5.4.1,http://memesuite.org/tools/meme)在線分析網站,對多刺綠絨蒿MYB轉錄因子家族成員進行保守基序的鑒定,并使用TBtools 軟件(version 1.09876)對保守基序分析結果進行可視化。
2.4 多刺綠絨蒿MYB轉錄因子系統發育分析
利用MEGA 軟件(version 11)內置的Clustal W程序,將多刺綠絨蒿106 個MYB 轉錄因子和擬南芥195 個MYB 轉錄因子蛋白序列以及52 個R2R3-MYB 和擬南芥126 個R2R3-MYB 轉錄因子蛋白質序列分別進行序列比對,選用鄰接(neighborjoining,NJ)法構建系統發育樹,Bootstrap method參數取值1000,其余參數選擇默認值,并使用iTOL網站(https://itol.embl.de/)進行進化樹美化。
2.5 多刺綠絨蒿R2R3-MYB 蛋白特征結構域分析
利用DNAMAN(version 9.0)軟件對多刺綠絨蒿R2R3-MYB 轉錄因子蛋白序列進行多序列比對,提取其R2 和R3 保守結構域中相應的氨基酸殘基分布,利用WebLogo 3 網站(http://weblogo.Threeplusone.com/)繪制特征結構域的序列標識。
3 結果與分析
3.1 多刺綠絨蒿MYB轉錄因子鑒定與理化性質分析
通過Hmmsearch 和blastp 比對在多刺綠絨蒿中共得到187 個MYB 轉錄因子,利用SMART、PROSITE、NCBI-CDD 和Pfam 數據庫篩選,去掉保守結構域不完整的和重復的序列,最終鑒定到106 個MYB 轉錄因子,其中1RMYB 類49 個、R2R3-MYB 類52 個、3R-MYB 類5 個,命名為MhMYB1~106(表1)。
對106 個多刺綠絨蒿MYB轉錄因子進行蛋白長度、相對分子質量、等電點和酸堿性等基本性質預測(表 1),結果顯示其蛋白序列長度為 77(MhMYB101)~976(MhMYB50)個氨基酸殘基;蛋白相對分子質量為8750(MhMYB101)~110 680(MhMYB50);等電點為4.720(MhMYB78)~10.452(MhMYB101),其中53 個轉錄因子蛋白等電點小于7,偏酸性,其中53 個轉錄因子等電點大于7,偏堿性;蛋白疏水性為-1.12(MhMYB37)~-0.31(MhMYB17),表明多刺綠絨蒿MYB轉錄因子均為親水性蛋白質;不穩定系數為28.348(MhMYB29)~71.011(MhMYB9),其中MhMYB77、MhMYB47、MhMYB44、MhMYB25、MhMYB30、MhMYB94 和MhMYB29 的不穩定系數系小于40 為穩定蛋白,其余蛋白序列不穩定系數均大于40,表明多刺綠絨蒿MYB轉錄因子以不穩定蛋白為主。亞細胞定位預測結果顯示106個MYB 轉錄因子有97個預測到細胞核(nuclear)中,8 個預測到線粒體(mitochondrial)中,1 個預測到細胞質(cytoplasmic)中,表明MYB 轉錄因子蛋白主要在細胞核中發揮作用。
為明確多刺綠絨蒿MYB轉錄因子的蛋白質結構組成,對MhMYBs 蛋白序列進行了二級結構預測(表1),結果顯示多刺綠絨蒿MYB轉錄因子家族二級結構主要由4 種類型組成,其中無規則卷曲(random coil)占比為29.07%~73.45%;α-螺旋結構(alpha helix)占比為11.43%~60.47%;β-轉角(beta turn)占比為1.02%~14.61%;延伸鏈(extend strand)占比為0~20.45%。
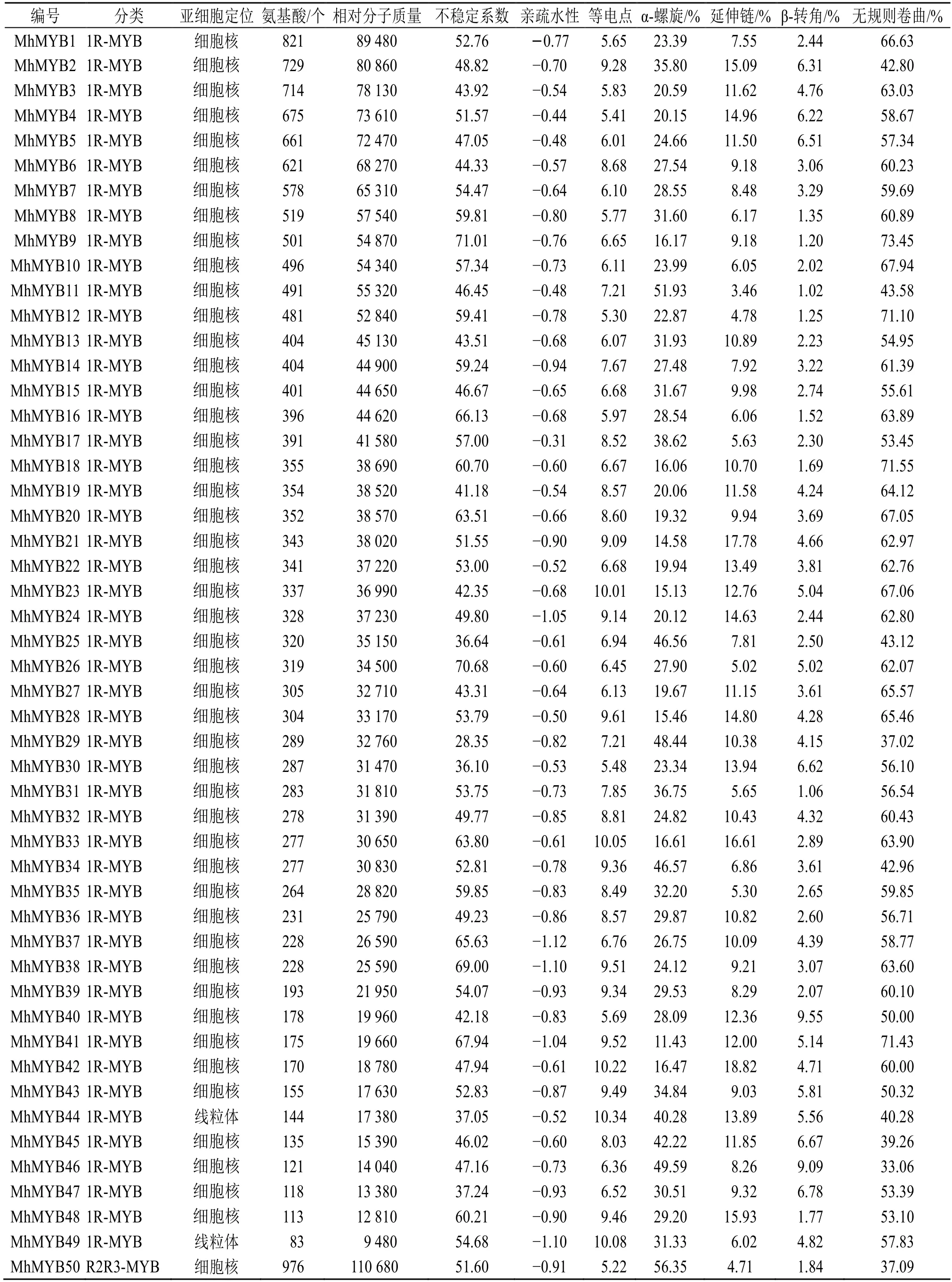
表1 多刺綠絨蒿MYB 蛋白基本信息及理化性質預測Table 1 Basic information and physicochemical properties prediction of MYB protein of M.horridula
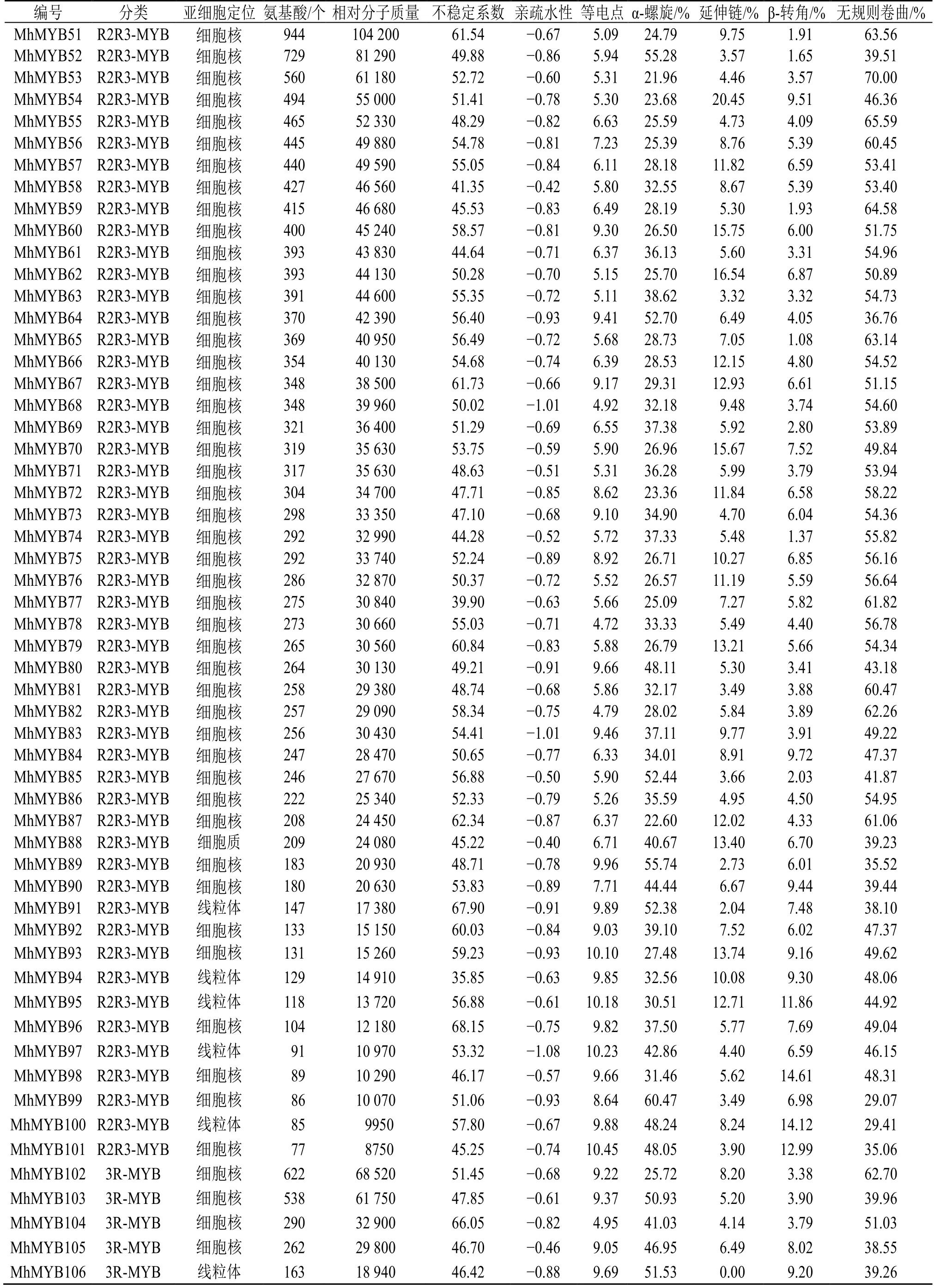
續表1
3.2 多刺綠絨蒿MYB轉錄因子GO 功能注釋
多刺綠絨蒿MYB轉錄因子GO 功能預測結果顯示(圖1),多刺綠絨蒿MYB轉錄因子蛋白主要注釋在細胞成分(cellular component,CC)、生物過程(biological process,BP)、和分子功能(molecular function,MF)3 大類。GO 富集分析結顯示,MYB轉錄因子蛋白功能在細胞成分(cellular component,CC)方面主要富集在細胞核(GO:0005634)、細胞上細胞器(GO:0043229)和核糖體(GO:0000786)等細胞成分;生物過程(biological process,BP)中主要富集在細胞發育過程(GO:0048869)、細胞分化(GO:0030154)和生物合成調節(GO:0048583)等方面;分子功能(molecular function,MF)方面主要富集在轉錄調節活性(GO:0140110)、DNA結合(GO:0003677)和有機環化合物結合(GO:0097159)等方面。

圖1 多刺綠絨蒿MYB轉錄因子家族GO 注釋Fig.1 GO annotation of MYB transcription factor family in M.horridula
3.3 多刺綠絨蒿MYB轉錄因子保守基序分析
為進一步說明多刺綠絨蒿MYB轉錄因子的多樣性和進化關系,利用MEME 網站進行保守基序分析,并結合系統發育樹進行可視化(圖2)。結果顯示,多刺綠絨蒿MYB轉錄因子家族共鑒定到10 個保守基序,其中motif3(89.62%)、motif1(51.89%)和motif7(46.23%)三者出現的頻率較高。其中2R-MhMYBs 亞家族基序保守性強,大多都包含了motif2、motif3、motif5、motif1 和motif7,且都位于蛋白序列N 端。1R-MhMYBs 保守基序的數量較少且位置變化較大,大多都包含了motif3 和motif4。結合系統進化分析發現,同一亞家族具有相似的motif 的種類與分布,不同亞家族在種類與分布上具有較大差異,且處于鄰近分支的MYB 轉錄因子的保守基序的位置結構與類型更相近。

圖2 多刺綠絨蒿MYB轉錄因子蛋白保守motifFig.2 Motif analysis of MYB transcription factors in M.horridula
3.4 多刺綠絨蒿MYB轉錄因子家族系統進化分析
對106 個多刺綠絨蒿MYB轉錄因子與擬南芥195 個MYB 轉錄因子的蛋白序列進行多序列比對及系統進化分析(圖3)。結果顯示,多刺綠絨蒿MYB轉錄因子不同亞家族成員均能與擬南芥相應類型的轉錄因子聚類到一起,該結果與結構域鑒定分類結果基本一致,說明MhMYBs 的鑒定及分類結果可靠,表明多刺綠絨蒿和擬南芥的MYB 轉錄因子家族具有相似的進化過程,具有比較高的保守性。

圖3 多刺綠絨蒿和擬南芥MYB 轉錄因子家族系統進化樹Fig.3 Phylogenetic tree of MYB transcription factors in M.horridula and A.thaliana
為研究多刺綠絨蒿R2R3-MhMYB 的系統進化關系,對多刺綠絨蒿與擬南芥R2R3-MYB 轉錄因子構建系統發育樹(圖 4),并結合擬南芥R2R3-MYB 蛋白亞組的分類依據[3],將多刺綠絨蒿R2R3-MYB 轉錄因子分為了24 個亞組(C1~C24),其中有18 個亞組能與擬南芥已分類亞組聚為一支,包括C1(S4)、C6(S1)和C10(S6)等,C3、C12 和C14 等6 個多刺綠絨蒿亞組包含擬南芥未分類的轉錄因子,有S3、S10、S12、S15和S16 這5 個擬南芥亞組未能與多刺綠絨蒿MYB轉錄因子聚類。多刺綠絨蒿與擬南芥R2R3-MYB轉錄因子的聚類結果表明,聚類在一起的轉錄因子具有較高的同源性與序列相似性,并可能具有類似的功能,多刺綠絨蒿與擬南芥物種后期朝不同的進化方向發展,也導致了部分R2R3-MYB 轉錄因子的特異性分化。

圖4 多刺綠絨蒿與擬南芥R2R3-MYB 系統進化樹Fig.4 Phylogenetic tree of R2R3-MYB of M.horridula and A.thaliana
3.5 多刺綠絨蒿R2R3-MYB 轉錄因子特征結構域分析
為研究多刺綠絨蒿R2R3-MYB 轉錄因子的結構域同源性特征,進一步展示在特定位置的保守性,分別對多刺綠絨蒿和擬南芥R2R3-MYB 蛋白序列進行多序列比對分析,并對R2 和R3 保守結構域特征進行可視化(圖5)。結果顯示多刺綠絨蒿R2R3-MYB 蛋白與擬南芥R2R3-MYB 蛋白相似,在R2 和R3 重復序列中存在多個保守的氨基酸,尤其是特征色氨酸(W),且都具有[-W-(X19)-W-(X19)-W-]和[-F-(X18)-W-(X18)-W-]結構域;其中R2 結構包括3 個極度保守的色氨酸(W),每2 個色氨酸殘基間隔19 個氨基酸殘基;R3 結構包括2 個極度保守的色氨酸(W),第1 個色氨酸被苯丙氨酸(F)、異亮氨酸(I)及亮氨酸(L)取代,第2 個色氨酸和第3 個色氨酸殘基間隔18 個氨基酸殘基;除高度保守的色氨酸(W)外,R2 和R3 結構中也存如賴氨酸(K)、蘇氨酸(T)、精氨酸(R)、天冬酰胺(N)、甘氨酸(G)和谷氨酸(E)等保守氨基酸殘基,這類保守氨基酸殘基通常出現在各重復區的后半部分,即第2 和第3 個的保守色氨酸之間的區域,且在每個重復區中與前2 個螺旋區域相比第3 個螺旋區保守性更高,表明這類保守氨基酸殘基具有維持MYB 結構域的HTH 結構的作用。
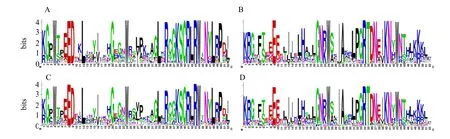
圖5 多刺綠絨蒿和擬南芥R2R3-MYB 轉錄因子特征結構域序列標識Fig.5 Conserved domain logos of R2R3-MYB transcription factors in M.horridula and A.thaliana
3.6 多刺綠絨蒿MYB轉錄因子表達分析
MYB 轉錄因子在非生物脅迫及植物黃酮類代謝方面具有重要調節作用,多刺綠絨蒿生長于青藏高原高海拔地區,生長環境惡劣多變,為探究多刺綠絨蒿MYB轉錄因子在不同海拔間的潛在關系,研究多刺綠絨蒿R2R3-MYB 轉錄因子與黃酮類合成途徑和非生物脅迫相關基因在不同海拔的表達特異性,對多刺綠絨蒿MYB轉錄因子進行表達模式分析(圖6)。結果顯示,106 個MYB 轉錄因子中,有28 個轉錄因子的表達量隨海拔升高而升高,11 個轉錄因子表達量隨海拔升高而降低。52 個R2R3-MYB 轉錄因子中,14個轉錄因子的表達量變化與海拔變化一致,5 個轉錄因子表達量與海拔變化相反。其中MhMYB90、MhMYB84和MhMYB62分別聚類到擬南芥黃酮類代謝物相關的S4、S6、S7 亞組,表達量均與海拔呈正相關;MhMYB 81、MhMYB 82,MhMYB63、MhMYB56、MhMYB55,MhMYB83,MhMYB101,MhMYB 58分別聚類到擬南芥非生物脅迫相關的S2、S13、S21、S22、S23 亞組,其中MhMYB81,MhMYB63、MhMYB56、MhMYB55,MhMYB83,MhMYB101,MhMYB58 表達量變化與海拔變化一致,MhMYB82 表達量變化與海拔變化相反。

圖6 不同海拔梯度下多刺綠絨蒿MYB轉錄因子的表達量分析Fig.6 Expression profiles of MYB transcription factors of M.horridula in different altitude gradients
4 討論
MYB 轉錄因子具有特有的MYB DNA 結合域,能夠特異結合基因啟動子區域的作用元件,根據植物生長發育及環境變化調控相關的基因的轉錄表達,是植物生長過程中的重要調節因子[26],對植物生長發育具有重要意義。目前已在擬南芥、小麥、大豆、刺梨、柑橘和楊梅等多個物種中被鑒定[27-30]。本研究基于多刺綠絨蒿轉錄組數據鑒定獲得了106個MYB 轉錄因子,其中包含49 個1R-MYB、52個R2R3-MYB 和5 個3R-MYB,通過對多刺綠絨蒿MYB轉錄因子理化性質預測顯示,多刺綠絨蒿MYB轉錄因子均為親水性蛋白質,53 個蛋白偏酸性,53 個蛋白偏堿性。不穩定系數顯示多刺綠絨蒿MYB轉錄因子中除了7 個為穩定蛋白,其余99 個都為不穩定蛋白,這與王靜文等[31]對細葉百合的研究結果相類似。在亞細胞定位中預測到細胞核、線粒體和細胞質3 種類型,且主要在細胞核中發揮作用,這與Cao 等[32]分析MYB 轉錄因子在中國梨的細胞核中發揮重要作用相似。二級結構預測表明多刺綠絨蒿MYB轉錄因子家族成員存在4 種二級結構,其中α-螺旋結構和無規則卷曲占比較高,蛋白質的二級結構與蛋白質的功能息息相關,表明這2種結構更有助于MYB 蛋白質的結構形成,這與張耐等[33]對甘草MYB 的研究及李明明等[34]對谷子MYB-CC 研究結果相一致。
多刺綠絨蒿保守基序分析表明,各亞家族motif組成相對保守,聚類到同一分支的MYB 因子具有更為相似的motif 組成和分布類型,而不同分支間則有所差異,這與谷佳等[35]對三色堇MYB 轉錄因子的研究相一致。保守基序分析能夠展現該家族進化過程中的結構特點,為其功能研究提供理論基礎,相似的保守基序組成和分布表明其可能在多刺綠絨蒿的生長發育過程中具有相似的功能作用,而距離較遠的分支其保守基序的位置與結構的出現差異可能是造成轉錄因子功能分化的重要因素[36]。對多刺綠絨蒿R2R3-MYB 特征結構域分析表明,該亞家族轉錄因子均含有[-W-(X19)-W-(X19)-W-]和[-F-(X18)-W-(X18)-W-]的2 個相鄰的典型結構域,這與Liu 等[37]對秈稻R2R3-MYB 的研究結果一致,表明多刺綠絨蒿R2R3-MYB轉錄因子在DNA結合結構域上具有較高的保守性,且在序列比對中發現在重復序列的第一個螺旋區及其臨近區域的保守性相對較低,常會發生氨基酸插入和缺失情況,推測該區域可能是該基因家族進化和功能分化的活躍區域,而色氨酸與其余保守度較高的氨基酸殘基對共同維持HTH 結構具有重要意義。
在植物中MYB 家族有選擇性地擴張,其中R2R3-MYB 是植物中存在最廣泛的一類MYB 轉錄因子,該亞家族成員因在植物生命活動中發揮的重要作用而被廣泛研究[38]。在多刺綠絨蒿中R2R3-MYB 轉錄因子可劃分為24 個亞組,其中有18 個亞組中含有不同數量已分類亞組的AtMYB 和MhMYB 蛋白,說明MYB 轉錄因子進化過程相對保守[8]。擬南芥作為模式物種,MYB 轉錄因子的功能研究較多,以擬南芥AtMYBs 的功能作為參考,結合系統發育關系和表達分析,為探討多刺綠絨蒿MYB轉錄因子的功能探究提供重要參考。聚類結果表明,MhMYB61 與擬南芥S9 的AtMYB16 聚類在一支,擬南芥AtMYB16 有助于花瓣表皮細胞的形成的功能[39];MhMYB77 與AtMYB20 聚類到一支,擬南芥AtMYB20 參與調控擬南芥次生細胞壁形成過程中木脂素和苯丙氨酸的生物合成[40]。在多刺綠絨蒿中有4 個(MhMYB72、MhMYB100、MhMYB75、MhMYB90)、2 個(MhMYB84、MhMYB95)和2 個(MhMYB62、MhMYBS54)轉錄因子分別與擬南芥中黃酮類化合物的調控機制以及花青素積累相關的S4、S6 和S7 亞組聚類為一支[41-43],推測此類多刺綠絨蒿轉錄因子也具有相似功能,表達量分析顯示其中MhMYB72、MhMYB75、MhMYB90、MhMYB84 和MhMYB62 這5 個轉錄因子在兩個高海拔居群中的表達量均高于低海拔居群,這可能與高海拔居群植株會生成黃酮類物質應對強光輻射以及干旱環境相關。擬南芥S4 亞組中的AtMYB4、AtMYB32 和AtMYB7 調控和表達能夠增強植物應對強紫外線脅迫的能力[44],聚類到同支的多刺綠絨蒿轉錄因子在高海拔居群中的上調表達可能也在多刺綠絨蒿應對紫外線脅迫中發揮重要作用。擬南芥S1、S11、S13、S21、S22、S23 亞組主要與抗旱抗寒等非生物脅迫相關[45-48],在多刺綠絨蒿的聚類和表達分析顯示共有15 個轉錄因子分別聚類在S1、S11、S13、S21、S22 和S23 這類亞組當中,其中有10 個轉錄因子在高海拔地區表達量上調,說明多刺綠絨蒿在應對更高海拔惡劣環境時會通過抗逆相關的轉錄因子的表達來調控次級代謝通路以應對脅迫環境,維持自身正常的生命活動。
目前,多刺綠絨蒿MYB轉錄因子的相關研究還非常欠缺,MYB 轉錄因子在多刺綠絨蒿生長發育調控及非生物脅迫的響應機制還不清楚,本研究通過生物信息學方法,鑒定了多刺綠絨蒿的MYB 轉錄因子,并系統地對該家族成員進行了基本理化性質、系統進化關系和表達模式等分析。為今后深入研究MYB 轉錄因子在多刺綠絨蒿生長發育中的作用和應對高原極端環境的調控機制提供了參考基礎。
利益沖突所有作者均聲明不存在利益沖突
