環境相關濃度雙酚AF暴露對成年雌性弓背青鳉生長和生殖的毒性效應
2023-05-23 06:22:21黃澤胤謝明花陳月碧黎學友郭昱嵩王中鐸董忠典
安徽農業科學 2023年8期
黃澤胤 謝明花 陳月碧 黎學友 郭昱嵩 王中鐸 董忠典



摘要 以成年雌性弓背青鳉(Oryzias curvinotus)為研究對象,研究了不同環境相關濃度(0.93、9.33和102.33 μg/L)雙酚AF(BPAF)短期暴露對其生存、生長和生殖的影響。結果表明,102.33 μg/L BPAF暴露21 d極顯著降低了成年雌性弓背青鳉的存活率和生長狀況因子;9.33和102.33 μg/L BPAF暴露導致成年雌性弓背青鳉肝細胞排列松散、脂肪積累減少,102.33 μg/L BPAF暴露還導致肝細胞形態出現明顯腫脹;102.33 μg/L BPAF暴露導致其卵母細胞大多停留在初級卵母細胞階段,抑制了卵母細胞的成熟。另外,102.33 μg/L BPAF暴露顯著或極顯著上調了vtglike、vtg1、vtg2和chgh在肝臟中的轉錄水平以及cyp11a、cyp19a1a和gsdf在卵巢中的轉錄水平。不同環境相關濃度的BPAF暴露對成年弓背青鳉雌魚有較強的致死效應、生長抑制效應、雌激素效應,并可引起肝損傷和卵母細胞發生障礙。
關鍵詞弓背青鳉;內分泌干擾物;雙酚AF;雌激素效應;生殖毒性
中圖分類號X174文獻標識碼A
文章編號0517-6611(2023)08-0079-07
doi:10.3969/j.issn.0517-6611.2023.08.019開放科學(資源服務)標識碼(OSID):
Toxic Effects of Bisphenol AF Exposure at Environmental Concentrations on the Growth and Reproduction of Adult Female Oryzias curvinotus
HUANG Ze-yin,XIE Ming-hua,CHEN Yue-bi et al(Key Laboratory of Aquaculture in South China Sea for Aquatic Economic Animal of Guangdong Higher Education Institutes/ Fisheries College of Guangdong Ocean University,Zhanjiang,Guangdong 524088)
AbstractAdult female Oryzias curvinotus were used as research objects to study the effects of short-term exposure with BPAF at different environmental concentrations (0.93,9.33 and 102.33 μg/L) on the survival, growth and reproduction of O.curvinotus.The results showed that the survival rate and condition factor of adult female O.curvinotus were extremely significantly reduced by the exposure with 102.33 μg/L BPAF for 21 days.Exposure with 9.33 and 102.33 μg/L BPAF resulted in loose arrangement of hepatocytes and the reduction of fat accumulation in adult female O.curvinotus.Exposure with 102.33 μg/L BPAF caused significant swelling of liver cell morphology.102.33 μg/L BPAF caused oocytes to stay in the primary oocyte stage and inhibited the maturation of oocytes.102.33 μg/L BPAF significantly or extremely significantly up-regulated the transcript levels of vtglike,vtg1,vtg2 and chgh in the liver,and the transcript levels of cyp11a,cyp19a1a and gsdf in the ovary.Exposure with environmental concentrations of BPAF had strong lethal effects,growth inhibitory effects,and estrogen effects on adult female O.curvinotus,and could cause liver damage and oocyte dysfunction.
Key wordsOryzias curvinotus;Endocrine disruptor;Bisphenol AF;Estrogen effects;Reproductive toxicity
內分泌干擾物(endocrine disrupting chemicals,EDCs)污染已被列為繼臭氧層空洞和地球變暖之后迫切需要治理的環境污染問題。雙酚A(bisphenol A,BPA)是最常見的內分泌干擾物,具有類雌激素效應、神經毒性、遺傳毒性和生殖毒性等[1-4],目前已被多個地區和行業禁用[5-6]。雙酚AF(bisphenol AF,BPAF)又名六氟雙酚A,是一種新型的含氟雙酚類物質,也是BPA的主要替代物之一。它在工業上主要用作氟橡膠硫化促進劑,也可用于電子產品、光纖、食品包裝盒加工、醫藥中間體等[7]。隨著BPAF用量的增加,在產品的制造、消費和廢棄過程中BPAF不可避免會釋放到各類環境介質中[6,8-9]。
水體作為容納BPAF的主要介質,近年來水體中BPAF含量呈現增加的趨勢[9-13]。德國地表水中BPAF的濃度為0.1~180.0 μg/L[13];嘉興市BPAF工廠附近河水中BPAF濃度高達15.3 μg/L[14]。2006—2007年,美國74個污水處理廠污水污泥樣品中BPAF檢出率為46%,干重1.79~72.20 μg/kg[15]。目前飲用水中BPAF的相關報道較少,但BPAF在水源和飲用水樣本中的檢出率僅次于BPA[16-17]。Mandrah等[18]在瓶裝碳酸飲料樣品中也檢測到BPAF。
研究表明,BPAF具有內分泌干擾效應、生殖毒性、生長和發育毒性以及神經毒性[19-26]。盡管現階段水環境中BPAF含量較低,但BPAF具有半衰期長、易富集、難降解等特性,同時由于其CF3基團具有更高的電負性和活性,即使低劑量的BPAF也可能具有更高的結合激素受體潛力,嚴重威脅水生生物的健康[27]。污染物的毒性受水體理化因子的影響,同一種污染物在不同鹽度、溫度、pH等環境條件下可能具有不同的毒性,此外不同的物種對相同污染物的敏感度也可能存在差異[28-29]。目前對BPAF的水生生態毒理研究主要集中在斑馬魚上[24-26,30-31]。若要準確評估BPAF的水生生態風險,有必要開展BPAF暴露對不同種類水生生物的生態毒性效應研究。
弓背青鳉(Oryzias curvinotus)體型小、適鹽范圍廣、易于實驗室養殖、世代周期短、產卵量大、胚胎發育透明且遺傳性別明確,是理想的海洋環境監測模型[32-34]。研究環境相關濃度BPAF對弓背青鳉生長和生殖的影響,有助于闡明BPAF對魚類的內分泌干擾效應機制,也為確定水體中BPAF的安全濃度提供支持。筆者以雌性弓背青鳉成魚為研究對象,使用不同環境濃度的BPAF對其進行短期暴露,從個體、組織、細胞、分子水平研究BPAF暴露對弓背青鳉生存、生長、組織結構及繁殖相關基因轉錄表達的影響,旨在為全面評估BPAF的生態風險提供科學依據。
1材料與方法
1.1試驗材料試驗所用弓背青鳉為南海水產經濟動物增養殖重點實驗室繁育的高橋群體F代成年雌魚(10月齡),試驗水體為經過曝氣的自來水。養殖水溫為(26 ± 1)℃,光暗周期為14 h∶10 h,pH 7.2~8.4,每天投喂新鮮孵化的豐年蟲(Artemia salina)2次。BPAF(CAS 1478-61-1)購自Sigma公司。取適量BPAF粉末溶于二甲基亞砜(dimethyl sulfoxide,DMSO,國產分析純),配制濃度分別為10、1和100 μg/mL 的儲存液,用棕色玻璃瓶儲存,并置于-20 ℃冰箱中暗處,備用。
1.2BPAF暴露試驗設計選取120條生長規格一致、健康的成年雌性弓背青鳉用于BPAF暴露試驗;選取12 L的玻璃缸作為暴露容器,暴露溶液體積為8 L,每缸隨機放入10條弓背青鳉。將弓背青鳉暴露于不同環境相關濃度(100、10和1 μg/L)的BPAF(即BPAF暴露處理組),另設置對照組(含0.01‰的DMSO)。所有BPAF暴露處理組均含有0.01‰(V/V)的DMSO。每個處理設置3個平行,試驗暴露周期為21 d,暴露水體每48 h全部更新。試驗過程中水溫、光照周期和投喂方式與常規飼養保持一致。暴露期間,每天用一次性吸管吸取殘餌和糞便,挑出并記錄死亡個體。分別在暴露第8天和第16天從每個玻璃缸中取相同體積暴露48 h的水體和新更換的水體,混合后送至廣東省科學院測試分析研究所(中國廣州分析測試中心)測定暴露水體中BPAF的濃度。暴露結束后,將所有魚置于冰上麻醉后立刻測量魚的體長和濕重,計算生長狀況因子(condition factor,K)。生長狀況因子計算公式為K=濕重/(體長)×100。測量結束后解剖,取肝臟和卵巢組織:肝臟和卵巢組織分成2份,1份置于4%的多聚甲醛(PFA)中,用于組織切片制備;另1份置于RNALater中,用于總RNA提取。
1.3組織切片制備和總RNA提取將獲得的部分肝臟和卵巢組織于4%的PFA中固定18 h,隨后進行乙醇溶液系列脫水、二甲苯透明、石蠟包埋、切片(厚度6~8 μm)、蘇木精-伊紅(H.E)染色,置于光學顯微鏡下觀察并拍照。取用RNALater保存的肝臟和卵巢組織,參照Trizol 試劑盒(Invitrogen,美國)說明書提取總RNA,通過瓊脂糖凝膠電泳檢測總RNA的完整性;然后,使用美國NanoDrop 2000(賽默飛,美國)測定總RNA樣品的A/A。所有RNA樣品18S和28S條帶清晰,A/A為1.8~2.0,符合后續試驗要求。
1.4cDNA制備和熒光定量分析參照HiScript 1st Strand cDNA Synthesis Kit(諾唯贊,南京)使用說明書,配制反轉錄體系,使用PCR儀進行反轉錄,獲得cDNA。將獲得的cDNA用雙蒸水(ddHO)稀釋20倍,-20 ℃下保存備用。參考筆者所在實驗室弓背青鳉轉錄組數據,獲取部分與生殖相關的基因序列[35-37],并設計熒光定量引物(表1)。qPCR反應在羅氏LC96熒光定量PCR儀上運行,反應體系共15.0 μL,包括2×PerfectStart TM Green qPCR SuperMix(全式金,北京)7.5 μL,上下游引物(10 μmol/L)各0.3 μL,cDNA 2.0 μL,ddHO 4.9 μL。反應程序如下:95 ℃ 預變性 30 s;95 ℃ 5 s,60 ℃ 15 s,72 ℃ 10 s(收集熒光),40個循環;隨后進行熔解曲線分析。以eef1b和rps4x為內參基因[37-38],對目的基因進行歸一化處理。定量結果采用2-ΔΔCt方法評估目的基因相對于對照組的相對表達量。
1.5數據統計與分析試驗數據均以平均值±標準差的形式表示,使用GraphPad Prism 8軟件進行數據統計分析并繪圖。采用單因素ANOVA檢驗分析方法中的Tukey多重比較,分析對照組與BPAF暴露處理組各數據的差異顯著性。
2結果與分析
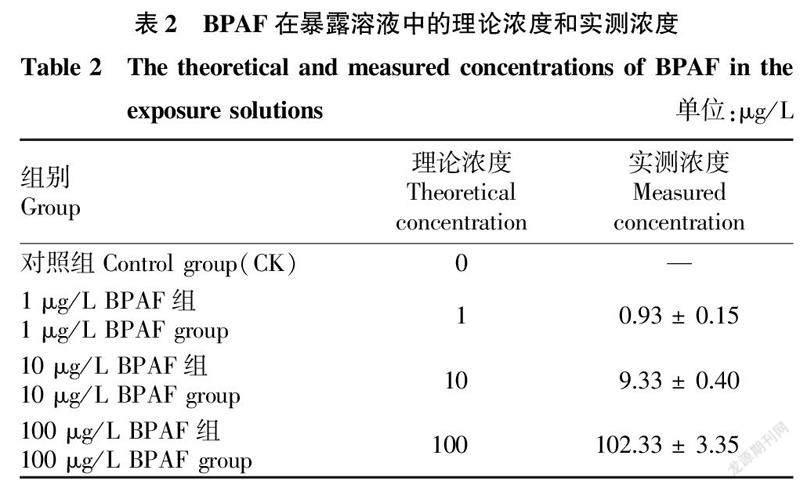
2.1BPAF在暴露水體中的實際濃度如表2所示,BPAF在暴露過程中的濃度保持穩定,3個BPAF處理組在試驗過程中BPAF實際濃度分別為0.93、9.33和102.33 μg/L,實測濃度與理論濃度接近;在對照組中未檢測到BPAF,說明試驗水體未被BPAF污染。所有處理組BPAF暴露濃度均以實測濃度表示。
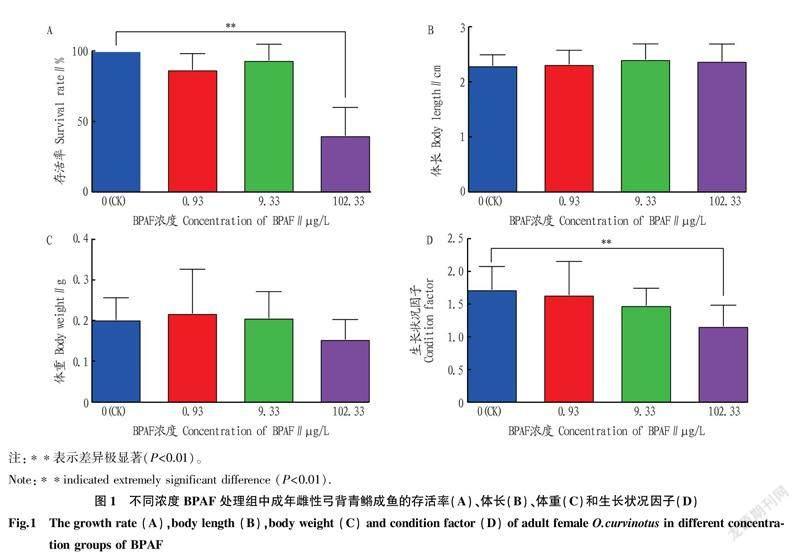
2.2BPAF暴露對雌性弓背青鳉成魚生存、生長的影響0.93和9.33 μg/L的BPAF暴露處理21 d未對雌性弓背青鳉成魚的生存造成影響,而102.33 μg/L的BPAF暴露處理極顯著降低了弓背青鳉的存活率(圖1A)。各濃度BPAF暴露沒有對弓背青鳉體長和體重造成顯著影響(圖1B、C),但102.33 μg/L BPAF暴露極顯著降低了弓背青鳉生長狀況因子(圖1D)。
2.3BPAF暴露對雌性弓背青鳉成魚肝臟和卵巢組織學結構的影響0.93 μg/L BPAF暴露處理21 d后弓背青鳉肝臟組織學結構(圖2B)在形態上與對照組(圖2A)相比沒有明顯差異,肝細胞呈多邊形且排列緊密,僅脂肪含量與對照組相比略有降低;9.33和102.33 μg/L BPAF暴露處理組肝細胞排列松散,肝臟細胞間間隙增大,脂肪含量降低(圖2C、D);102.33 μg/L BPAF暴露還導致肝細胞出現明顯的腫脹和形狀改變(圖2D)。對照組弓背青鳉的卵巢含有各期卵母細胞(圖3A),0.93 μg/L BPAF暴露處理未對卵巢結構造成明顯的影響(圖3B),但9.33和102.33 μg/L的BPAF暴露處理抑制了弓背青鳉卵母細胞的成熟,卵母細胞多停留在初級卵母細胞階段(primary oocyte,PO)(圖3C、D)。
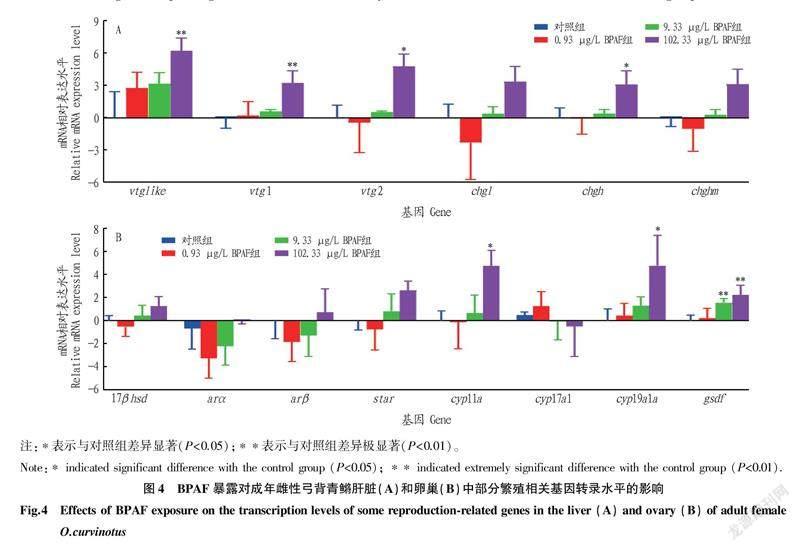
51卷8期黃澤胤等環境相關濃度雙酚AF暴露對成年雌性弓背青鳉生長和生殖的毒性效應2.4BPAF暴露對雌性弓背青鳉成魚基因轉錄水平的影響在肝臟中,102.33 μg/L的BPAF暴露可以顯著上調卵黃蛋白原(vitellogenins,vtgs)和透明帶蛋白前體(choriogenins,chgs)基因的轉錄水平,其中vtg2和chgh的上調顯著(P<0.05),vtglike和vtg1的上調極顯著(P<0.01),chgl和chghm的上調不顯著(P>0.05);0.93和9.33 μg/L的BPAF暴露未對vtgs和chgs的表達造成顯著影響(圖4A)。在卵巢中,102.33 μg/L的BPAF暴露顯著上調了細胞色素P450芳香化酶(cyp19a1a)基因、細胞色素p450介導側鏈裂解酶(cholesterolside chain cleavage enzyme,cyp11a)基因,另外9.33和102.33 μg/L的BPAF暴露極顯著上調了gsdf的轉錄水平(圖4B)。
3討論
該研究在雌性弓背青鳉成魚暴露過程中BPAF的實測濃度與理論濃度基本一致,表明在48 h暴露時間內BPAF的穩定性較高,這與其半衰期較長且難降解的特性[27]相符,提示BPAF對水生生物具有持久性危害。
該研究中102.33 μg/L BPAF暴露導致成年弓背青鳉雌魚存活率顯著下降。目前關于BPAF對水生生物的致死濃度研究較少。Yang等[31]使用1.0 mg/L的BPAF對2月齡斑馬魚進行28 d暴露處理,結果發現試驗組雌、雄斑馬魚的存活率與對照組相比沒有顯著差異;陳亞文[24]研究發現用500 μg/L 的BPAF處理成年斑馬魚30 d并未導致斑馬魚死亡;楊洋等[30]研究發現BPAF暴露24 h對斑馬魚胚胎致畸的半數效應濃度為2.00 mg/L,暴露96 h的半數致死濃度為1.84 mg/L;Mu等[39]研究了BPAF、BPA、BPF和BPS對斑馬魚胚胎的半數致死濃度,分別為1.95、10.43、19.59和>50 mg/L。郭婧穎等[40]通過毒性試驗測得BPAF對大型溞(Daphnia magna)的24 h和48 h半數致死濃度分別為9.70和5.02 mg/L。綜合以上研究可推測出成年雌性弓背青鳉對水體中BPAF的耐受程度較斑馬魚和大型溞等水生動物更低,水體中BPAF會對弓背青鳉的生存造成威脅。
肝臟在魚類物質能量代謝、激素合成等生理過程中具有重要作用,是魚類監測環境污染物毒性效應的主要靶點[41-44],也是動物或人體富集BPAF的重要器官[45]。該研究中短期BPAF暴露導致雌性弓背青鳉成魚肝臟細胞排列松散、肝細胞體積變大。類似的結果也出現在BPAF和其他含氟類有機物的研究中,250和1 000 μg/L BPAF暴露組雄性斑馬魚中出現肝細胞排列松散,1 000 μg/L BPAF暴露組肝細胞明顯腫大、形狀不規則,但相同處理的雌性斑馬魚肝臟并未發生明顯改變[31];全氟十二烷酸腹腔注射斑馬魚,短時間即導致斑馬魚出現肝損傷,包括肝細胞腫脹、空泡化和核固縮等[46];全氟辛烷磺酸暴露處理雌性大鼠后,導致幼鼠肝細胞腫大[47]。因此,雌性弓背青鳉成魚肝臟在組織學上可作為監測水體中BPAF等含氟類化合物的靶點,顯著的肝臟病理學損傷可能是102.33 μg/L BPAF處理組弓背青鳉成魚死亡率高的誘因之一。另外,成年雌魚肝臟是卵黃蛋白原(vitellogenins,vtgs)、透明帶蛋白前體(choriogenins,chgs)等基因的主要表達器官。這些基因對環境中雌激素類物質敏感,在無外界因素刺激的情況下僅在雌魚肝臟中轉錄,在雄魚及幼魚中不發生轉錄,而當水環境中存在雌激素類物質時則誘導這些基因的大量表達[48-50]。該研究中102.33 μg/L BPAF誘導vtglike、vtg1、vtg2和chgh的轉錄水平顯著上升,chgl和chghm的轉錄水平也明顯上調。該結果符合BPAF具有雌激素效應的特性,類似的結果在250 μg/L BPAF暴露處理的雌性斑馬魚肝臟中也有發現[31];Mu等[39]研究表明BPAF顯著提高了斑馬魚胚胎中vtg1的mRNA 水平。綜合BPAF對弓背青鳉肝臟組織病理學特征和基因轉錄水平的影響,推斷BPAF對雌性弓背青鳉成魚既有雌激素效應又有肝臟毒性效應。
9.33 和102.33 μg/L BPAF暴露阻滯了雌性弓背青鳉成魚卵母細胞的發育,這與以前的研究結果相一致。Yang等[31]研究發現BPAF短期暴露(250和1 000 μg/L暴露28 d)導致雌性斑馬魚卵母細胞發育阻滯;陳亞文[24]研究表明經BPAF處理的斑馬魚卵巢中卵母細胞發育受到抑制,其成熟卵母細胞數量較對照組顯著減少;BPAF急性暴露還可能導致性成熟雌性小鼠性腺形態異常和卵巢未分化的導管末端小葉增生[51]。因此,水體中環境相關濃度的BPAF對雌性弓背青鳉成魚具有較強的生殖毒性。該研究中BPAF暴露改變了雌性弓背青鳉成魚卵巢中cyp11a、cyp19a1a和gsdf的轉錄水平。在性激素合成過程中,CYP11A1是已知的唯一催化膽固醇向孕烯醇酮轉化的酶,后者是所有類固醇激素的共同前體[52]。cyp19a1a編碼的性腺芳香化酶可將雄激素轉化為雌激素,從而促進卵巢分化[53-54]。該研究中BPAF暴露上調了雌性弓背青鳉成魚卵巢中cyp11a和cyp19a1a的轉錄水平。性腺體細胞衍生因子(gonadal soma derived factor,GSDF)屬于TGF-β超家族成員,主要參與魚類性別分化,對于維持魚類精原細胞的正常增殖發育具有重要作用[55-56];敲除羅非魚的gsdf導致遺傳雄魚性逆轉為雌魚,而過表達gsdf致使遺傳雌魚的性腺發育為精巢[57]。該研究中9.33和102.33 μg/L BPAF暴露顯著上調了雌性弓背青鳉成魚卵巢gsdf轉錄水平,這可能抑制了卵巢的成熟。據此推測BPAF對雌性弓背青鳉成魚的生殖毒性可能與BPAF改變了繁殖相關基因的轉錄水平有關。
4結論
該研究結果表明,水體中環境相關濃度的BPAF暴露對雌性弓背青鳉成魚的生存和生長造成了不利影響。BPAF暴露還導致雌性弓背青鳉成魚肝損傷,上調了肝臟中vtgs和chgs轉錄水平,導致了卵巢結構異常,阻礙了卵母細胞的成熟,影響了卵巢中cyp11a、cyp19a1a和gsdf的轉錄水平。作為一種新型的內分泌干擾化合物,BPAF致毒機理及其對水生生物的危害需要引起更多的關注。
參考文獻
[1] SAKAUE M,OHSAKO S,ISHIMURA R,et al.Bisphenol-A affects spermatogenesis in the adult rat even at a low dose[J].Journal of occupational health,2001,43:185-190.
[2] MA A T,YANG X Z,WANG Z X,et al.Adult exposure to diethylstilbestrol induces spermatogenic cell apoptosis in vivo through increased oxidative stress in male hamster[J].Reproductive toxicology,2008,25(3):367-373.
[3] NAKAMURA D,YANAGIBA Y,DUAN Z W,et aNl.Bisphenol A may cause testosterone reduction by adversely affecting both testis and pituitary systems similar to estradiol[J].Toxicology letters,2010,194(1/2):16-25.
[4] VANDENBERG L N,CHAHOUD I,HEINDEL J J,et al.Urinary,circulating,and tissue biomonitoring studies indicate widespread exposure to bisphenol A[J].Ciência & saúde coletiva,2012,17(2):407-434.
[5] YE X B,PIERIK F H,ANGERER J,et al.Levels of metabolites of organophosphate pesticides,phthalates,and bisphenol A in pooled urine specimens from pregnant women participating in the Norwegian Mother and Child Cohort Study (MoBa) [J].International journal of hygiene & environmental health,2009,212(5):481-491.
[6] 宋作棟,仇雁翎,張華,等.水體中雙酚類物質的賦存現狀及研究進展[J].環境化學,2020,39(6):1496-1503.
[7] AKAHORI Y,NAKAI M,YAMASAKI K,et al.Relationship between the results of in vitro receptor binding assay to human estrogen receptor alpha and in vivo uterotrophic assay:Comparative study with 65 selected chemicals[J].Toxicology in vitro,2008,22(1):225-231.
[8] YANG Y J,YU J L,YIN J,et al.Molecularly imprinted solid-phase extraction for selective extraction of bisphenol analogues in beverages and canned food[J].Journal of agricultural & food chemistry,2014,62(46):11130-11137.
[9] JIN H B,ZHU L Y.Occurrence and partitioning of bisphenol analogues in water and sediment from Liaohe River Basin and Taihu Lake,China[J].Water research,2016,103:343-351.
[10] YAN Z Y,LIU Y H,YAN K,et al.Bisphenol analogues in surface water and sediment from the shallow Chinese freshwater lakes:Occurrence,distribution,source apportionment,and ecological and human health risk[J].Chemosphere,2017,184:318-328.
[11] LIU Y H,ZHANG S H,SONG N H,et al.Occurrence,distribution and sources of bisphenol analogues in a shallow Chinese freshwater lake (Taihu Lake):Implications for ecological and human health risk[J].Science of the total environment,2017,599/600:1090-1098.
[12] SI W,CAI Y F,LIU J C,et al.Investigating the role of colloids on the distribution of bisphenol analogues in surface water from an ecological demonstration area,China[J].Science of the total environment,2019,673:699-707.
[13] FROMME H,KCHLER T,OTTO T,et al.Occurrence of phthalates and bisphenol A and F in the environment[J].Water research,2002,36(6):1429-1438.
[14] SONG S J,RUAN T,WANG T,et al.Distribution and preliminary exposure assessment of bisphenol AF (BPAF) in various environmental matrices around a manufacturing plant in China[J].Environmental science & technology,2012,46(24):13136-13143.
[15] YU X H,XUE J C,YAO H,et al.Occurrence and estrogenic potency of eight bisphenol analogs in sewage sludge from the U.S. EPA targeted national sewage sludge survey[J].Journal of hazardous materials,2015,299:733-739.
[16] SUN X L,PENG J Y,WANG M H,et al.Determination of nine bisphenols in sewage and sludge using dummy molecularly imprinted solid-phase extraction coupled with liquid chromatography tandem mass spectrometry[J].Journal of chromatography A,2018,1552:10-16.
[17] ZHANG H F,ZHANG Y P,LI J B,et al.Occurrence and exposure assessment of bisphenol analogues in source water and drinking water in China[J].Science of the total environment,2019,655:607-613.
[18] MANDRAH K,SATYANARAYANA G N V,ROY S K.A dispersive liquid-liquid microextraction based on solidification of floating organic droplet followed by injector port silylation coupled with gas chromatography-tandem mass spectrometry for the determination of nine bisphenols in bottled carbonated beverages[J].Journal of chromatography A,2017,1528:10-17.
[19] YAMASAKI K,TAKEYOSHI M,SAWAKI M,et al.Immature rat uterotrophic assay of 18 chemicals and Hershberger assay of 30 chemicals[J].Toxicology,2003,183(1/2/3):93-115.
[20] YAMASAKI K,TAKEYOSHI M,YAKABE Y,et al.Comparison of reporter gene assay and immature rat uterotrophic assay of twenty-three chemicals[J].Toxicology,2002,170(1/2):21-30.
[21] MATSUSHIMA A,LIU X H,OKADA H,et al.Bisphenol AF is a full agonist for the estrogen receptor ERα but a highly specific antagonist for ERβ[J].Environmental health perspectives,2010,118(9):1267-1272.
[22] KITAMURA S,SUZUKI T,SANOH S,et al.Comparative study of the endocrine-disrupting activity of bisphenol A and 19 related compounds[J].Toxicological sciences,2005,84(2):249-259.
[23] GU J,WANG H Y,ZHOU L J,et al.Oxidative stress in bisphenol AF-induced cardiotoxicity in zebrafish and the protective role of N-acetyl N-cysteine[J/OL].Science of the total environment,2020,731[2021-11-15].https://doi.org/10.1016/j.scitotenv.2020.139190.
[24] 陳亞文.六氟雙酚A暴露對成魚期斑馬魚的毒性效應[D].海口:海南大學,2016.
[25] 唐天樂.六氟雙酚A對斑馬魚(Danio rerio)甲狀腺軸及認知能力的干擾效應研究[D].海口:海南大學,2016.
[26] 楊洋.雙酚AF暴露對胚胎/幼魚期斑馬魚的毒性效應及甲狀腺內分泌系統的干擾效應[D].海口: 海南大學,2016.
[27] 吳迪.雙酚AF對小鼠血睪屏障和精子發生的影響及其機制研究[D].武漢:華中農業大學,2020.
[28] YOU C,JIA C X,PAN G.Effect of salinity and sediment characteristics on the sorption and desorption of perfluorooctane sulfonate at sediment-water interface[J].Environmental pollution,2010,158(5):1343-1347.
[29] 黃乾生,陳亞檞,方超,等.鹽度影響全氟辛烷磺酸對海水青鳉(Oryzias melastigma)的毒性[J].科學通報,2013,58(2):151-157.
[30] 楊洋,陳亞文,唐天樂,等.雙酚AF暴露對胚胎期和幼魚期斑馬魚的毒性效應[J].環境科學研究,2015,28(8):1219-1226.
[31] YANG X X,LIU Y C,LI J,et al.Exposure to bisphenol AF disrupts sex hormone levels and vitellogenin expression in zebrafish[J].Environmental toxicology,2016,31(3):285-294.
[32] 董忠典,龍水生,黃承勤,等.一種快速鑒定弓背青鳉遺傳性別的方法[J].廣東海洋大學學報,2018,38(3):25-29.
[33] 黃順楷,郭昱嵩,汪淳,等.弓背青鳉3種雌激素受體基因的克隆及其表達分析[J].廣東海洋大學學報,2019,39(2):8-19.
[34] 張海瑞,王中鐸,黃順楷,等.弓背青鳉的胚胎發育及自發熒光觀察[J].廣東海洋大學學報,2019,39(2):38-44.
[35] 董忠典,黎學友,廖健,等.雌、雄弓背青鳉(Oryzias curvinotus)肝臟轉錄組比較分析[J].海洋與湖沼,2020,51(5):1203-1213.
[36] DONG Z D,LI X Y,YAO Z B,et al.Oryzias curvinotus in Sanya does not contain the male sex-determining gene dmy[J].Animals,2021,11(5):1-13.
[37] DONG Z D,LI X Y,HUANG S K,et al.Vitellogenins and choriogenins are biomarkers for monitoring Oryzias curvinotus juveniles exposed to 17 β-estradiol[J].Comparative biochemistry and physiology part C:Toxicology & pharmacology,2020,236:1-10.
[38] DONG Z D,CHEN P S,ZHANG N,et al.Evaluation of reference genes for quantitative real-time PCR analysis of gene expression in Hainan medaka (Oryzias curvinotus)[J].Gene reports,2019,14:94-99.
[39] MU X Y,HUANG Y,LI X X,et al.Developmental effects and estrogenicity of bisphenol A alternatives in a zebrafish embryo model[J].Environmental science & technology,2018,52(5):3222-3231.
[40] 郭婧穎,劉建超,李帥衡,等.雙酚AF對大型溞生殖、生長等生態行為的影響[J].中國環境科學,2019,39(10):4394-4400.
[41] MONDON J A,HOWITT J,TOSIANO M,et al.A simple osmium post-fixation paraffin-embedment technique to identify lipid accumulation in fish liver using medaka (Oryzias latipes) eggs and eleutheroembryos as lipid rich models[J].Marine pollution bulletin,2011,63(5/6/7/8/9/10/11/12):86-90.
[42] ZHANG X Y,WEN H S,WANG H L,et al.RNA-Seq analysis of salinity stress-responsive transcriptome in the liver of spotted sea bass (Lateolabrax maculatus)[J].PLoS One,2017,12(3):1-18.
[43] NI X M,WAN L,LIANG P P,et al.The acute toxic effects of hexavalent chromium on the liver of marine medaka (Oryzias melastigma) [J].Comparative biochemistry and physiology,part C:Toxicology & pharmacology,2020,231:1-11.
[44] HOSHIKAWA Y,FURUKAWA S,IRIE K,et al.Sequential histological changes in the liver of medaka exposed to methylazoxymethaol acetate[J].Journal of toxicologic pathology,2020,33(4):219-226.
[45] YANG Y J,YIN J,YANG Y,et al.Determination of bisphenol AF (BPAF) in tissues,serum,urine and feces of orally dosed rats by ultra-high-pressure liquid chromatography-electrospray tandem mass spectrometry[J].Journal of chromatography B,2012,901:93-97.
[46] LIU Y,WANG J S,WEI Y H,et al.Induction of time-dependent oxidative stress and related transcriptional effects of perfluorododecanoic acid in zebrafish liver[J].Aquatic toxicology,2008,89(4):242-250.
[47] BJORK J A,LAU C,CHANG S C,et al.Perfluorooctane sulfonate-induced changes in fetal rat liver gene expression[J].Toxicology,2008,251(1/2/3):8-20.
[48] CHEN X P,LI V W,YU R M K,et al.Choriogenin mRNA as a sensitive molecular biomarker for estrogenic chemicals in developing brackish medaka (Oryzias melastigma)[J].Ecotoxicology & environmental safety,2008,71(1):200-208.
[49] LEE C,NA J G,LEE K C,et al.Choriogenin mRNA induction in male medaka,Oryzias latipes as a biomarker of endocrine disruption[J].Aquatic toxicology,2002,61(3/4):233-241.
[50] LAI K P,LI J W,WANG S Y,et al.Tissue-specific transcriptome assemblies of the marine medaka Oryzias melastigma and comparative analysis with the freshwater medaka Oryzias latipes[J].BMC genomics,2015,16(1):1-13.
[51] TUCKER D K,BOUKNIGHT S H,BRAR S S,et al.Evaluation of prenatal exposure to bisphenol analogues on development and long-term health of the mammary gland in female mice[J].Environmental health perspectives,2018,126(8):1-17.
[52] HU M C,HSU H J,GUO I C,et al.Function of Cyp11a1 in animal models[J].Molecular and cellular endocrinology,2004,215(1/2):95-100.
[53] SI Y F,DING Y X,HE F,et al.DNA methylation level of cyp19a1a and Foxl2 gene related to their expression patterns and reproduction traits during ovary development stages of Japanese flounder (Paralichthys olivaceus)[J].Gene,2016,575(2):321-330.
[54] KOBAYASHI Y,HORIGUCHI R,MIURA S,et al.Sex-and tissue-specific expression of P450 aromatase (cyp19a1a) in the yellowtail clownfish,Amphiprion clarkii[J].Comparative biochemistry and physiology part A:Molecular & integrative physiology,2010,155(2):237-244.
[55] SAWATARI E,SHIKINA S,TAKEUCHI T,et al.A novel transforming growth factor-beta superfamily member expressed in gonadal somatic cells enhances primordial germ cell and spermatogonial proliferation in rainbow trout (Oncorhynchus mykiss)[J].Developmental biology,2007,301(1):266-275.
[56] SHIBATA Y,PAUL-PRASANTH B,SUZUKI A,et al.Expression of gonadal soma derived factor (GSDF) is spatially and temporally correlated with early testicular differentiation in medaka[J].Gene expression patterns,2010,10(6):283-289.
[57] JIANG D N,YANG H H,LI M H,et al.gsdf is a downstream gene of dmrt1 that functions in the male sex determination pathway of the Nile tilapia[J].Molecular reproduction and development,2016,83(6):497-508.
