葡萄屬植物遺傳轉化影響因素研究進展
2023-07-13 21:55:22張?zhí)飓h趙若岑
山西果樹 2023年4期
關鍵詞:影響因素
張?zhí)飓h 趙若岑
摘 要:葡萄作為世界上重要的經(jīng)濟類果樹作物之一,在釀酒、鮮食、制干、制汁、制罐、砧木等方面的應用十分廣泛,為人類創(chuàng)造了明顯的社會效益、經(jīng)濟效益和生態(tài)效益。自美國學者1990年在沙地葡萄(Vitis rupestris)中首次成功獲得遺傳轉化植株,葡萄屬植物遺傳轉化發(fā)展迅猛。多年研究證實利用分子育種建立完整、高效的葡萄遺傳轉化體系對葡萄基因改良有著至關重要的作用。闡述了葡萄屬植物遺傳轉化的4個階段,并對遺傳轉化的影響因素進行具體分析,針對這些影響因素進行資料收集和對比研究,總結葡萄遺傳轉化體系研究現(xiàn)狀并提出展望,以期對葡萄基因改良研究提供參考。
關鍵詞:葡萄;遺傳轉化;影響因素
文章編號:2096-8108(2023)04-0067-06? 中圖分類號:S663.1 ?文獻標識碼:A
Research Progress on Influencing Factors of Genetic Transformation of Grape Plants
ZHANG? Tianyue,ZHAO? Ruocen
(Faculty of Food and Wine, Ningxia University, Yinchuan Ningxia 750021,China)
Abstract: Grape is one of the most important fruit in the world, which is widely used in wine making, fresh food, drying, juice making, pot making and stock making, and has created considerable social, economic and ecological benefits for human beings. Since American scholars first obtained genetically modified plants in Vitis rupestris in 1990, the genetic transformation of grape plants has developed rapidly. Years of research have confirmed that the use of molecular breeding to establish a complete and efficient grape genetic transformation system plays a vital role in grape genetic improvement. In this paper, the four stages of genetic transformation of grape plants are elaborated, and the influencing factors of genetic transformation are analyzed in detail. Data collection and comparative study of these influencing factors are carried out, and the research status of grape genetic transformation system is summarized and prospects are proposed in order to provide reference for grape genetic improvement research.
Keywords:grape; genetic transformation; influencing factors
葡萄作為世界上重要的經(jīng)濟類果樹作物之一,在釀酒、鮮食、制干、制汁、制罐、砧木等方面的應用十分廣泛,為人類創(chuàng)造了明顯的社會效益、經(jīng)濟效益和生態(tài)效益[1-2]。由于葡萄屬植物染色體高度雜合,遺傳背景復雜,采用常規(guī)雜交育種獲得葡萄新品種周期長,工作繁瑣,已不適應當今產(chǎn)業(yè)高速發(fā)展的需求。與傳統(tǒng)的育種技術相比,分子育種可以克服偶然性大、育種年限長和后代數(shù)量龐大不易篩選等缺點,能夠精確改進目前應用品種的不足,實現(xiàn)多個優(yōu)良基因(性狀)的聚合。因此,利用分子育種建立完整、高效的葡萄遺傳轉化體系對葡萄屬植物品種改良有著至關重要的作用。
歐洲葡萄( Vitis vinifera L.)具有較高的經(jīng)濟價值,品質優(yōu)良且栽培廣泛,但其抗逆性較差,改良優(yōu)質葡萄品種抗逆性是葡萄育種的目標之一[3]。近年來葡萄轉基因研究主要采取農(nóng)桿菌介導法進行,基因槍轉化法的應用復雜且花費高,使用次數(shù)相對較少。美國學者MullinsMG等人自1990年首次在沙地葡萄(Vitis rupestris)中獲得轉基因植株,Scorza R和Franks tricia以‘無核白( Vitis vinifera L.‘Thompson seedless)和‘薩娜( Vitis vinifera L.‘Sultana)為材料,轉外殼蛋白基因和uidA/nptⅡ基因的植株已被分別獲得[4-6]。21世紀初,周鵬等[7]人在葡萄中導入人胰島素樣生長因子IGF-I基因,孫仲序等[8]人將魯貝葡萄花絲所謂外植體誘導體胚,最終獲得表達甜菜堿醛脫氫酶基因的葡萄,鮑睿等[9]人建立了歐洲葡萄無核白遺傳轉化體系,張瑩[10]于2021年初步建立‘霞多麗種子遺傳轉化體系,葡萄屬植物基因遺傳轉化技術日益廣泛應用于葡萄屬植物的研究。
多年研究證實利用分子育種建立完整、高效的葡萄遺傳轉化體系對葡萄基因改良有著至關重要的作用。隨著分子育種技術的不斷精進,研究者通過器官發(fā)生(Organogenesis)和胚狀體發(fā)生(Somatic embryogenesis)兩大途徑建立了葡萄再生體系。遺傳轉化體系是以再生體系為基礎建立的。基因型、受體類型、培養(yǎng)基類型和轉化方式等都是遺傳轉化體系的重要影響因素,本文針對這些影響因素進行資料收集和對比研究,分析葡萄遺傳轉化體系研究現(xiàn)狀并提出展望,以期對葡萄基因改良研究提供參考。
1 葡萄屬植物遺傳轉化體系的建立
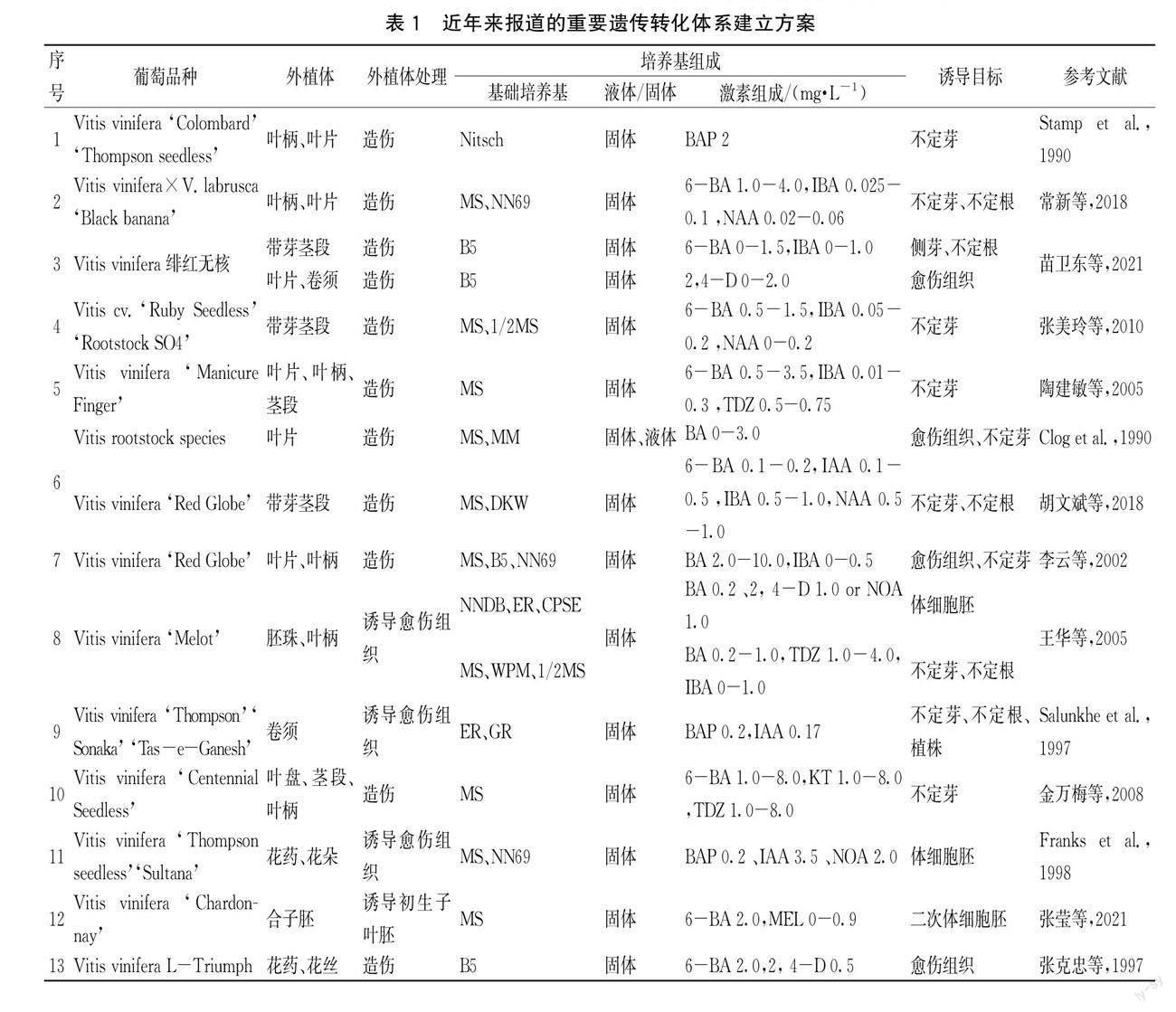
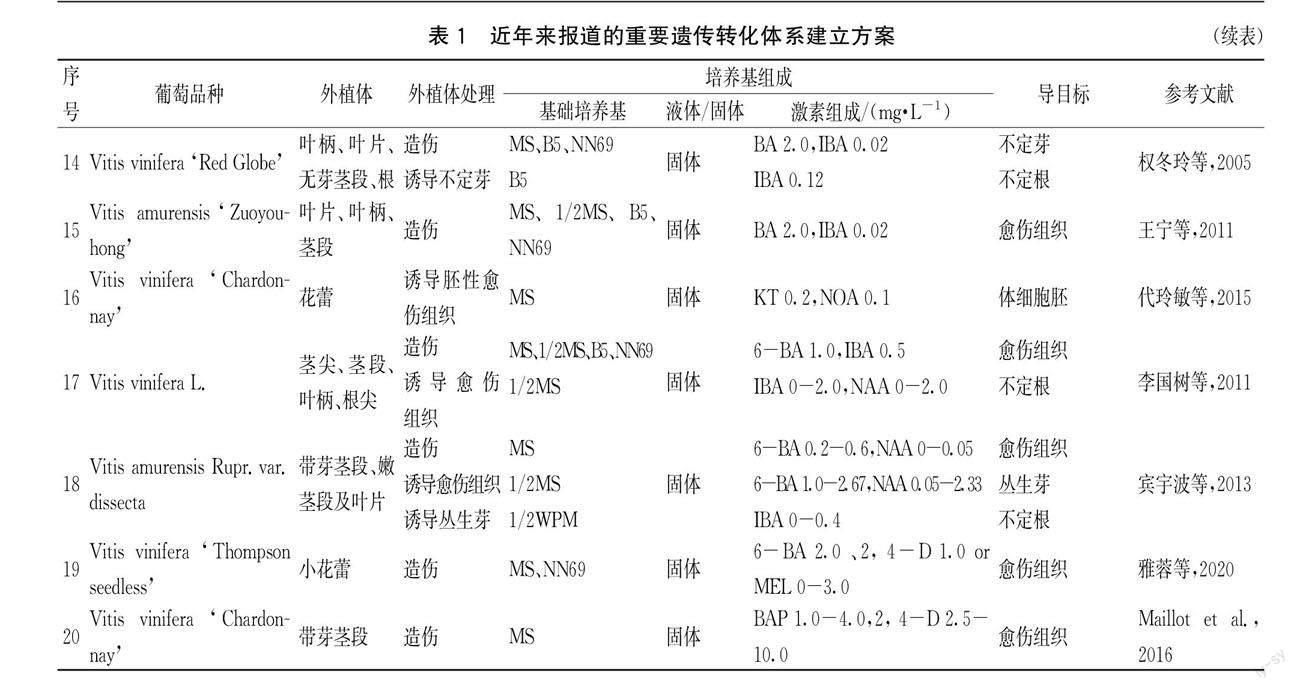
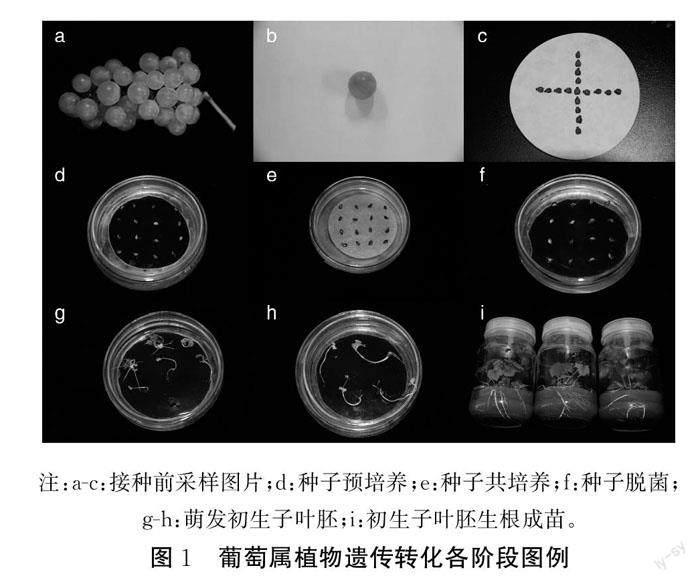
葡萄屬植物遺傳轉化可以分為以下4個階段:第1階段是遺傳轉化受體系統(tǒng)的建立。這1階段主要是準備用于遺傳轉化的受體,主要受到培養(yǎng)途徑和培養(yǎng)條件的影響;第2階段是目的基因的導入。借助各種不同的方法將目的基因高效準確的導入受體,該階段主要受到目的基因和轉化方法的影響;第3階段是受體生長與植株發(fā)育。遺傳轉化的最終目的是培養(yǎng)轉基因植株。這一階段需要給受體提供適宜的生長環(huán)境,使受體繼續(xù)生長直至成苗,并且將沒有成功導入基因的受體篩選出來;第4階段是目的基因的分析與檢測。根據(jù)目的基因攜帶的報告基因,對導入的目的基因進行檢測,檢驗遺傳轉化是否成功。近年來有重要推進意義的遺傳轉化報道詳見表1,各階段圖片展示詳見圖1。
2 不同因素對葡萄屬植物遺傳轉化的影響
2.1 受體類型
遺傳轉化受體系統(tǒng)主要包括器官發(fā)生和胚狀體發(fā)生兩大類。近年來,可用于遺傳轉化受體誘導方法及條件的研究報道甚多,詳見表2。器官發(fā)生是指在一定誘導條件下,由離體的細胞、組織經(jīng)分裂、增殖以及再分化,形成根、芽等器官的過程。葉器官和根器官是用于器官發(fā)生途徑的主要外植體。由表2可以得出,不同品種器官發(fā)生的誘導率差異較大,不同外植體誘導率沒有普遍規(guī)律,且不同品種之間差異較為明顯。胚狀體發(fā)生是指具有胚芽、胚根和胚軸的胚狀結構經(jīng)培養(yǎng)細胞誘導分化后,再生長為一個完整植株的過程[11]。Mauro M C和Scorza R證明了胚性細胞比葉片、葉柄和莖段等有較高的再生能力[5,12]。目前用于遺傳轉化的有胚性愈傷組織、原胚團和體細胞胚3種類型的胚性組織[13-14],其中胚性愈傷組織和體細胞胚的報道較多。與莖段,葉片相比,將胚性愈傷組織用作農(nóng)桿菌侵染轉化外植體的優(yōu)勢在于轉化胚性愈傷組織能夠高頻的進行再生植株的轉化,通過篩選可以避免嵌合體轉化,降低假陽性,從而獲得真正意義上的轉基因植株。
2.2 培養(yǎng)條件
無論是器官發(fā)生途徑還是胚狀體發(fā)生途徑,培養(yǎng)條件都是影響外植體生長發(fā)育的關鍵因素。對于某一基因型葡萄的特定外植體而言,必須在某一特定的培養(yǎng)基中和培養(yǎng)條件下才能再生出植株[15]。大量研究表明,培養(yǎng)外植體的基礎培養(yǎng)基和激素種類及濃度是最重要的影響因素。不同外植體的最適培養(yǎng)基不同,所以,根據(jù)不同的外植體,我們要選用最適合其生長的基礎培養(yǎng)基。對于器官發(fā)生途徑,誘導愈傷組織最適基礎培養(yǎng)基為MS和B5,誘導不定芽的最適培養(yǎng)基與品種和外植體類型有很大關系,常用的有MS、NN69和B5培養(yǎng)基。對于胚狀體發(fā)生途徑,誘導體胚常用的基礎培養(yǎng)基有NN69、MS、1/2MS、PIV、MC和改良培養(yǎng)基。不同外植體所使用的基礎培養(yǎng)基一般不盡相同,含芽莖段和花絲一般接種于1/2 MS基礎培養(yǎng)基,而葉柄、葉片和卷須一般接種于NN69基礎培養(yǎng)基[16]。有研究表明,在胚狀體形成階段,有利于胚狀體生長發(fā)育的是低濃度生長素和中低濃度細胞分裂素,其他外源激素則不再需要[17]。
2.3 轉化方法
遺傳轉化體系的建立可以依靠不同的轉化方式導入基因,而轉化方式不同,對于不同受體材料的轉化率也有所不同。目前,主要用于葡萄基因轉化的方式有農(nóng)桿菌介導法和基因槍法。
2.3.1 基因槍法
基因槍法是通過一種類似槍結構的裝置,將表面附有外源遺傳物質的金屬顆粒射入受體,其表面吸附的外源遺傳物質也會隨之進入細胞[18]。該方法轉化效率高、受體細胞類型限制少、操作簡單且可控度高,但儀器費用過高,花費較多。研究發(fā)現(xiàn),通過葡萄懸浮細胞均可實現(xiàn)轉化,分別獲得了基因轉化植株和愈傷組織[19-20]。
2.3.2 農(nóng)桿菌介導法
農(nóng)桿菌介導法是導入目的基因的主要方法,以其費用低、重復性好、基因沉默現(xiàn)象少、轉化周期短以及能轉化較大片段等優(yōu)點而備受科學工作者的青睞。農(nóng)桿菌株系和菌液濃度對于研究有較大影響,而菌株本身致病力的強弱和外界誘導致病力表達的因素也同樣是影響葡萄屬植物遺傳轉化的關鍵因素[15]。對于農(nóng)桿菌的種系,大量研究顯示,與GV3101、LBA4404等菌株相比,EHA系列的菌株具有更好的轉化效果。趙亭亭[21]研究EHA105、GV3101和LBA4404共3種不同的農(nóng)桿菌菌株對葡萄遺傳轉化的影響時發(fā)現(xiàn)EHA105和GV3101的轉化效率顯著高于LBA4404,且二者無明顯差異,但農(nóng)桿菌EHA105更易于活化且具有更廣泛的適用性。對于菌液濃度,研究人員發(fā)現(xiàn)低濃度的農(nóng)桿菌菌液可以降低植物性毒素帶來的作用[22]。Tao J在研究苜蓿時發(fā)現(xiàn)菌液濃度(OD600)0.3~0.5時,GUS 的陽性愈傷率明顯超出其他濃度[23]。農(nóng)桿菌介導法也常與基因槍法結合使用來提高轉化效率。Bouanama B和Bidney D將微粒轟擊和農(nóng)桿菌介導轉化結合應用,發(fā)現(xiàn)比常規(guī)基因槍轉化的轉化率提高至少100倍[24-25]。
2.4 抗生素濃度
對于運用農(nóng)桿菌介導法進行目的基因導入的受體材料,篩選時所用的抗生素的種類、濃度對葡萄遺傳轉化能否成功至關重要。對于抗生素的選擇,卡那霉素在遺傳轉化中篩選效果比其他的抗生素效果好,在較低濃度下仍然可以有較好的篩選力,而巴龍霉素容易對外植體有過多抑制導致組織無法生長[26]。但后來的研究者認為巴龍霉素可將無效用細胞快速消滅,避免卡那霉素長續(xù)性抑制細胞生長[27]。抗生素的種類由載體和農(nóng)桿菌菌株決定,使用濃度因物種而異,濃度過低會導致出現(xiàn)嵌合體及假轉化體;濃度過高,毒性也會過高,使植物細胞無法生存,死細胞也會抑制臨近活細胞的生長[28]。因此,進行預試驗確定抗生素使用濃度十分重要。經(jīng)研究發(fā)現(xiàn)對于葡萄種子,當卡那霉素為30 mg/L就可起到篩選作用,70 mg/L卡那霉素濃度篩選效果最為明顯[29]。當受體生長出子葉胚時,應及時將其轉移到成苗培養(yǎng)基中,并黑暗環(huán)境下培養(yǎng)[30]。
3 遺傳轉化中存在的問題及展望
3.1 胚性愈傷誘導困難且嵌合體較多
據(jù)介紹,目前研究人員多選擇胚性組織作為遺傳轉化的受體,但胚性愈傷組織的誘導難度較大,過程繁瑣復雜,誘導率極低,使遺傳轉化效率大大降低[16]。直接將器官作為外植體進行再生,可以有效提高再生頻率,但很容易在遺傳轉化時發(fā)生嵌合體,而且容易發(fā)生變異,很難保證與母株基因型轉化后的植物基因型保持一致[31]。因此,探索更有利于遺傳轉化的再生受體系統(tǒng)和研究各獨立品種的最佳受體和再生方案是今后葡萄遺傳轉化研究的重點之一,如此便能進一步提高胚性愈傷組織的誘導率,有效降低器官再生嵌合體出現(xiàn)的幾率。
3.2 基因沉默
基因沉默在葡萄基因工程的研究中是一個普遍存在的現(xiàn)象。共抑制、位置效應、DNA甲基化、反式失活、重復序列等都會引起基因的沉默,基因被成功導入植株卻不表達,嚴重制約了葡萄基因工程的發(fā)展,難以讓試驗與生產(chǎn)實踐相結合[32]。基因沉默現(xiàn)象的發(fā)生與轉基因條件和人為因素也密切相關,所以,深入研究引起沉默的機理,調(diào)控轉基因過程的各個條件,有利于葡萄屬植物基因工程研究的發(fā)展。
3.3 潛在生態(tài)病害
轉基因技術在8年前就成為了全球的熱點話題,盡管轉基因技術在提高作物產(chǎn)量和質量、改善作物抗逆性等方面發(fā)揮作用,但在一定程度上也危害了人體。研究顯示,插入外源基因會導致生物體內(nèi)基因組不穩(wěn)定,出現(xiàn)各種不良變異,甚至會出現(xiàn)危害人類性狀的現(xiàn)象[32]。就轉基因而言,它會產(chǎn)生新的有害生物類型,外源基因逃逸對其他植物的造成基因污染,引發(fā)潛在生態(tài)危害,如產(chǎn)生新的毒性或者過敏原等[32]。因此,應加強轉基因植物特異表達的研究,對于尚未完全成熟的成果不應急于應用到生產(chǎn)實際當中,盡可能避免轉基因植物對人體與生態(tài)環(huán)境傷害。
葡萄屬植物遺傳轉化體系已經(jīng)發(fā)展了30年之久,但仍存在很多亟待解決的問題。經(jīng)過研究人員幾十年共同努力,釀酒葡萄‘黑比諾基因組測序已初步完成;與葡萄果實品質相關優(yōu)良抗病、抗蟲、抗寒、抗旱、抗鹽堿等基因也被不斷發(fā)掘[33];許多栽培品種和砧木品種已建立較為成熟的再生體系;抗病蟲和抗毒素等基因的成功轉化,初步建立遺傳轉化體系,葡萄育種工作取得不可小覷的進展。但是,目前葡萄屬植物遺傳轉化效率較低,嚴重制約了葡萄基因工程的發(fā)展。未來,我們應聚焦提高遺傳轉化效率和簡化轉化過程兩大內(nèi)容,盡快解決葡萄屬植物遺傳轉化效率低的問題。近年來,葡萄基因工程的研究發(fā)展勢頭正盛,相信在未來會有更多優(yōu)良品質、抗逆性強的葡萄品種出世,基因工程的研究定能帶領葡萄產(chǎn)業(yè)走向更光明的未來。
中文致謝
參考文獻
[1] BISSON L F, WATERHOUSE A L, EBELER S E, et al.The present and future of the international wine industry[J]. Nature, 2002, 418(6898): 696-699.
[2] 賀普超,羅國光.葡萄學[M].中國農(nóng)業(yè)出版社, 1999.
[3] VIDAL JR, GOMEZ C, CUTANDA MC, et al. Use of gene transfer technology for functional studies in grapevine[J]. Australian Journal of Grape and Wine Research,2010(16):138-151.
[4] MULLINS MG, TANG FA &FACCIOTTI D.Agrobacterium-mediated genetic transformation of grapevines: transgenic plants of Vitis rupestris Scheele and buds of Vitis vinifera L.[J].Nature Biotechnology,1990(8):1041-1045.
[5] SCORZA R, CORDTS JM, GRAY DJ, et al. Producing Transgenic ‘Thompson Seedless grape (Vitis vinifera L.) plants[J]. Journal of the American Society for Horticultural Science, 1996, 121(4):616-619.
[6] FRANKS TRICIA, HE DING GANG, THOMAS MARK. Regeneration of transgenic shape Vitis vinifera L. Sultana plants: genotypic and phenotypic analysis[J]. Molecular Breeding, 1998, 4(4) : 321-333.
[7] 周鵬,王躍進,賀普超.人胰島素樣生長因子-I基因轉化葡萄的研究[J].熱帶作物學報,2002(1):54-61.
[8] 孫仲序,陳受宜,王建設,等.農(nóng)桿菌介導BADH基因轉化葡萄的研究[J].果樹學報,2003(2):89-92.
[9] 鮑睿.中國野生毛葡萄VqRSGT基因的遺傳轉化與功能研究[D].西北農(nóng)林科技大學,2017.
[10] 張瑩.基于‘霞多麗葡萄花蕾與種子的體細胞胚再生體系建立及農(nóng)桿菌介導基因轉化的研究[D].寧夏大學,2021.
[11] 劉昱.葡萄體細胞胚的誘導及VvACS1基因對葡萄的遺傳轉化[D].河南農(nóng)業(yè)大學,2018.
[12] MAURO M C, TOUTAINA S, WALTERB B. High efficiency regeneration of grapevine plants transformed with the GFLV coat protein gene[J].Plant Science, 1995,112(1):97-106.
[13] DHEKNEY S A, LI Z T, DUTT M, et al. Agrobacterium-mediated transformation of embryogenic cultures and plant regeneration in Vitis rotundifolia Michx. (Muscadine grape)[J].Plant Cell Rep., 2008,27(5): 865-872.
[14] FAN C H, PU N, WANG X, et al. Agrobacterium-mediated genetic transformation of grapevine (Vitis vinifera L.) with a novel stilbene synthase gene from Chinese wild Vitis pseudoreticulata[J].Plant Cell Tiss. Org.,2008, 92(2): 197-206.
[15] 李云,馮慧,田硯亭.葡萄再生系統(tǒng)研究進展[J].生物技術通報,2000(2):28-31.
[16] 劉昱,付濛濛,安棟梁,等.葡萄體細胞胚發(fā)生及遺傳轉化體系研究進展[J].分子植物育種,2018,16(18):6068-6079.
[17] 譚超.葡萄再生體系建立的影響因素[J].農(nóng)家之友(理論版),2011(1):26-27.
[18] SANFORD J C,KLEIN T M,WOLF E D, et al. Delivery of substances into cells and tissues using a particle bombardment process[J].Particul Sci Technol,1987(5):27-37.
[19] HEBERTD.Optimization of biolistic transformation of embryogenicgrape cell suspensions[J].Plant Cell Rep,1993(12):585-589.
[20] SCORZA R,CORDTS JM,RAMMING DM, et al. Transformation of grape (Vitis vinifera L.) zygotic derived somatic embryos and rsgeneration of transgenic plants[J].Plant Cell Reports,1995,14(9):589-592.
[21] 趙亭亭.山葡萄高效遺傳轉化愈傷體系的建立[D].中國科學院武漢植物園,2017.
[22] IOCCO P,F(xiàn)RANKS T, THOMAS M R.Genetic transformation of major wine grape cultivars of Vitis Vinifera L.[J]. Transgenic Research,2002,10(2):105-112.
[23] TAO J, LING L. Genetic transformation of Torenia fournieri L.mediated by Agrobaterium rhizogenes[J]. South African Journal of Botany,2006(72):211-216.
[24] BOUAMAMA B, MLIKI A, GHORBEL A. Efficiency of coupling biolistic and Agrobacterium in genetic transformation of Tunisian autochthonous grapes[J]. Agricoltura Mediterranea,2000,130(3-4):223-227.
[25] BIDNEY D, SCELONGE C, M ART ICH J, et al. Microprojectile bombardment of plant tissues increases transformation frequency by Agrobacterium tumef aciens[J].Plant Mol.Biol.,1992(18):301-314.
[26] KIKKERT J, THOMAS M, REISCH B. Grapevine genetic engineering[J]. Molecular Biology & Biotechnology of the Grapevine.2001:393-410.
[27] WANG Q, PING L, HANANIA U, et al. Improvement of Agrobacterium-mediated transformation efficiency and transgenic plant regeneration of Vitis vinifera L. by optimizing selection regimes and utilizing cryopreserved cell suspensions[J]. Plant Science, 2005, 168(2):565-571.
[28] PENA L, CERVERA M, JUAREZ J,et al. Agrobacterium-mediated tranformation of sweet orange and regeneration of transgenic plants[J].Plant Cell Rep,1995(14):616-619.
[29] 張瑩,雅蓉,徐偉榮,等.褪黑素在霞多麗葡萄種子體細胞胚誘導發(fā)生中的作用[J].果樹學報,2021,38(6):922-933.
[30] 雅蓉,徐偉榮,張瑩,等.褪黑素對‘無核白葡萄體細胞胚的誘導作用[J].園藝學報,2020,47(5):953-962.
[31] 張麗,王敬東,宋玉霞.農(nóng)桿菌介導的葡萄轉化研究進展[J].生物技術通報,2012(2):14-19.
[32] 董志剛,馬小河,趙旗峰,等.葡萄遺傳轉化體系研究進展與展望[J].山西農(nóng)業(yè)科學,2013,41(11):1264-1269.
[33] SCORZA R,CORDTS JM,RAMMING DM, et al. Transformation of grape (Vitis vinifera L.) zygotic derived somatic embryos and rsgeneration of transgenic plants[J].Plant Cell Reports,1995,14(9):589-592.
[36] 莊智敏. 農(nóng)桿菌介導AtNHX1基因轉化葡萄砧木99R的研究[D].南京農(nóng)業(yè)大學,2006.
[37] 楊麗娜,劉捷,莊智敏,等.根癌農(nóng)桿菌介導Barnase基因轉化‘美人指葡萄的研究[J].南京農(nóng)業(yè)大學學報,2009,32(1):31-35.
[38] 趙鳳霞,王正平,高相彬,等.農(nóng)桿菌介導的葡萄胚性與非胚性愈傷組織遺傳轉化過程中脅迫響應基因的表達(英文)[J].果樹學報,2011,28(6):964-971,1149.
[39] 梁宏偉,陳發(fā)菊,王玉兵,等.根癌農(nóng)桿菌介導hGLP1基因轉化摩爾多瓦葡萄的研究[J].西北農(nóng)業(yè)學報,2013,22(2):131-135.
[40] 張修銘.中國野生華東葡萄VpCN基因轉化歐洲葡萄無核白的研究[D].西北農(nóng)林科技大學,2017.
[41] 賈慧.葡萄抗白粉病VvBAK1和VvLecRK1 CRISPR/Cas9載體構建及轉化葡萄的研究[D].西北農(nóng)林科技大學,2019.
[42] STAMP J A, COLBY S M, Meredith C P. Improved shoot organogenesis from leaves of grape[J].Journal of the American Society for Horticultural Science,1990,115(6):1038-1042.
[43] 常新,梁晉軍,劉振華,等.黑香蕉葡萄不同器官發(fā)生途徑再生體系的優(yōu)化[J].中國農(nóng)業(yè)大學學報,2018,23(9):43-52.
[44] 苗衛(wèi)東,井朋偉,王萌,等.克瑞森無核葡萄不同器官的離體培養(yǎng)[J].山東農(nóng)業(yè)科學,2021,53(7):27-33.
[45] 張美玲,楊國順,石雪暉,等.紅寶石無核和SO4單芽莖段的增殖與成苗培養(yǎng)[J].中外葡萄與葡萄酒,2010(7):28-30+30+33.
[46] 陶建敏,莊智敏,章鎮(zhèn),等.美人指葡萄不定芽離體誘導再生植株的研究[J].果樹學報,2005(5):551-553,599.
[47] CLOG E, BASS P, WALTER B. Plant regeneration by organogenesis in Vitis, rootstock species.[J].Plant Cell Reports, 1990,8(12):726-728.
[48] 胡文斌,張少飛,孫娜.隴南紅提葡萄莖段組織培養(yǎng)研究[J].陜西林業(yè)科技,2018,46(1):6-9.
[49] 李云,馮慧,田硯亭.‘紅地球葡萄葉片、葉柄不定芽再生體系的建立[J].園藝學報,2002(1):60-62.
[50] 王華,盧江,馬春花,等.釀酒葡萄“梅爾諾”再生系統(tǒng)建立的研究[J].西北植物學報,2005(5):858-863.
[51] SALUNKHE C K, RAO P S, MHATRE M. Induction of somatic embryogenesis and plantlets in tendrils of Vitisvinifera L.[J].Plant Cell Reports, 1997,17(1):65-67.
[52] 金萬梅,董靜,閆愛玲,等.葡萄器官離體再生和遺傳轉化體系的建立[J].園藝學報,2008(1):27-32.
[53] 張克忠,鮑雪珍,白永延,等.蘇云金桿菌內(nèi)毒素蛋白基因轉入葡萄胚性愈傷組織細胞及轉基因植株再生的研究[J].試驗生物學報,1997,30(3):303-308.
[54] 權冬玲,常永義.葡萄不同器官直接再生不定芽研究[J].甘肅農(nóng)業(yè)大學學報,2005(2):173-177.
[55] 王寧,宗成文,湯曉艷,等.山葡萄“左優(yōu)紅”離體器官再生不定芽的研究[J].遼寧林業(yè)科技,2011(1):27-29.
[56] 代玲敏,周起,楊亞州,等.歐洲葡萄‘霞多麗體細胞胚再生途徑建立與轉化華東葡萄芪合成酶基因[A].中國園藝學會.中國園藝學會2015年學術年會論文摘要集[C].中國園藝學會:中國園藝學會,2015.
[57] 李國樹,邱璐,徐成東,等.葡萄組織培養(yǎng)快速繁殖體系研究[J].北方園藝,2011(9):134-136.
[58] 賓宇波,沙海峰,任建武,等.百花山葡萄組織培養(yǎng)和快速繁殖[J].西北林學院學報,2013,28(6):99-102.
[59] MAILLOT PASCALE, DEGLENE-BENBRAHIM LAURENCE, WALTER BRENAID. Efficient somatic embryogenesis from meristematic explants in grapevine (Vitis vinifera L.) cv. Chardonnay: an improved protocol[J].Trees, 2016,30(4): 1377-1387.
收稿日期:2023-05-10中文收稿日期
基金項目:基于釀酒葡萄種子受體材料的遺傳轉化體系建立項目(G2021107490021)。中文作者簡介
第一作者簡介:張?zhí)飓h(2001-),女,本科在讀,主要從事葡萄組織培養(yǎng)及栽培技術研究。中文作者簡介
猜你喜歡
現(xiàn)代經(jīng)濟信息(2016年19期)2016-10-20 18:46:44
現(xiàn)代經(jīng)濟信息(2016年19期)2016-10-20 18:12:28
現(xiàn)代經(jīng)濟信息(2016年19期)2016-10-20 16:20:30
中國科技博覽(2016年19期)2016-10-19 13:33:22
中國科技博覽(2016年18期)2016-10-19 10:49:54
中國科技博覽(2016年18期)2016-10-19 08:16:45
中國科技博覽(2016年18期)2016-10-19 06:39:44
中國市場(2016年36期)2016-10-19 03:54:01
中國市場(2016年35期)2016-10-19 02:30:10
商(2016年27期)2016-10-17 07:09:07
