大負(fù)荷運(yùn)動(dòng)后骨骼肌核心時(shí)鐘基因Bmal1/Clock、Cry1/Per1的節(jié)律性振蕩變化趨勢(shì)
2023-08-07 01:11:52丁海麗傅澤鋌夏雨
中國(guó)體育科技 2023年7期
丁海麗,傅澤鋌,夏雨
生命的各種行為、生理和代謝功能都具有生物節(jié)律。生物節(jié)律是指生命活動(dòng)按一定時(shí)間順序、周而復(fù)始地發(fā)生變化的節(jié)律,主要由生物鐘和外界環(huán)境信號(hào)共同調(diào)控(Dibner et al., 2010)。生物鐘本質(zhì)上是一組構(gòu)成自主節(jié)律振蕩的時(shí)鐘基因,其周期性特異表達(dá)形成的“轉(zhuǎn)錄—翻譯—反饋”環(huán)路(transcriptional-translational feedback loop,TTFL)是生物節(jié)律核心分子機(jī)制(Harfmann et al., 2015;Takahashi, 2017)。系列研究表明,哺乳動(dòng)物時(shí)鐘基因變異與腸道菌群穩(wěn)態(tài)失衡、腎功能異常、呼吸道過(guò)敏性疾病、骨代謝疾病等密切相關(guān)(程馮麗 等,2021;劉程程 等,2021;熊勇紅 等,2020;余明芳 等,2020;Thaiss et al., 2014;Voigt et al., 2016)。人體的生物節(jié)律尤其是晝夜節(jié)律能夠影響運(yùn)動(dòng)能力及運(yùn)動(dòng)表現(xiàn),運(yùn)動(dòng)也能對(duì)生物節(jié)律產(chǎn)生反作用。骨骼肌作為運(yùn)動(dòng)的動(dòng)力結(jié)構(gòu)和直接效應(yīng)器官,可受到時(shí)鐘基因調(diào)節(jié)參與各項(xiàng)生理功能與活動(dòng)(Mayeuf-Louchart et al., 2015)。而運(yùn)動(dòng)作為一種授時(shí)因子(zeitgeber)能夠影響骨骼肌節(jié)律變化,同時(shí)也受到骨骼肌固有時(shí)鐘的調(diào)控。因此,明確運(yùn)動(dòng)訓(xùn)練后骨骼肌時(shí)鐘基因的變化趨勢(shì)、在觀測(cè)競(jìng)技體育及運(yùn)動(dòng)促進(jìn)健康的時(shí)間生物學(xué)效應(yīng)、探討擇時(shí)訓(xùn)練的內(nèi)在機(jī)制、改善生物鐘失調(diào)導(dǎo)致的節(jié)律紊亂等方面具有重要意義。
在生物體內(nèi),生物鐘通過(guò)時(shí)鐘基因反饋回路自主產(chǎn)生晝夜節(jié)律。研究發(fā)現(xiàn),大腦/肌肉芳香經(jīng)受體核轉(zhuǎn)位因子樣蛋白1(brain and muscle ARNT-like1, Bmal1)、晝夜運(yùn)動(dòng)輸出周期蛋白(circadian locomoter output cycles protein kaput, Clock)、Period(Per)及Cryptochrome(Cry)等基因?qū)Σ溉轭?lèi)動(dòng)物的生物鐘調(diào)控起著重要的作用(Keleher et al., 2014)。哺乳動(dòng)物體內(nèi)的生物時(shí)鐘主要由2個(gè)轉(zhuǎn)錄翻譯反饋環(huán)路組成,包括核心環(huán)路和穩(wěn)定環(huán)路。核心環(huán)路的關(guān)鍵結(jié)構(gòu)是轉(zhuǎn)錄因子Bmal1和Clock產(chǎn)物形成的異二聚體(Crnko et al., 2019),Bmal1/Clock在TTFL中起正向調(diào)控作用,能夠與Per和Cry等其他時(shí)鐘基因的啟動(dòng)子序列結(jié)合,激活其特異性表達(dá)并形成Per/Cry復(fù)合物,經(jīng)蛋白質(zhì)定向轉(zhuǎn)運(yùn)返回細(xì)胞核,可反抑制Bmal1/Clock活性,從而以負(fù)反饋方式調(diào)控自身基因的轉(zhuǎn)錄(Curtis et al.,2014)。
運(yùn)動(dòng)表現(xiàn)能力與時(shí)間生物學(xué)因素密切相關(guān),時(shí)鐘基因是導(dǎo)致運(yùn)動(dòng)表現(xiàn)差異的原因之一,Cry1/2作為過(guò)氧化物酶體增殖物激活受體δ(peroxisome proliferator-activator receptor,PPARδ)共抑制因子可調(diào)控運(yùn)動(dòng)能力(Jordan et al.,2017);Per1/2雙敲除小鼠在擇時(shí)運(yùn)動(dòng)中并未顯示出運(yùn)動(dòng)持續(xù)時(shí)間和血糖水平的差異(Ezagouri et al., 2019);肌肉特異性Bmal1基因敲除可致小鼠肌纖維類(lèi)型向I型轉(zhuǎn)變(Vitale et al., 2019)。運(yùn)動(dòng)還可以調(diào)控骨骼肌分子時(shí)鐘。研究發(fā)現(xiàn)計(jì)劃性運(yùn)動(dòng)可影響骨骼肌自身節(jié)律表達(dá),使時(shí)鐘基因相位前移2~3個(gè)小時(shí),而不影響中樞節(jié)律變化(Harfmann et al., 2015);去除嚙齒類(lèi)動(dòng)物骨骼肌運(yùn)動(dòng)神經(jīng)元會(huì)導(dǎo)致晝夜節(jié)律轉(zhuǎn)錄活性失調(diào)(Nakao et al., 2015);盡管這些研究初步驗(yàn)證了運(yùn)動(dòng)與骨骼肌生物節(jié)律的相互關(guān)系,但運(yùn)動(dòng)訓(xùn)練過(guò)程中骨骼肌時(shí)鐘基因分子機(jī)制尚未被完全認(rèn)識(shí)。基于此,本研究采用一次下坡跑建立大負(fù)荷運(yùn)動(dòng)大鼠模型,檢測(cè)骨骼肌組織Bmal1、Clock 、Cry1、Per1表達(dá),分析其節(jié)律振蕩變化趨勢(shì),旨在探討大負(fù)荷運(yùn)動(dòng)訓(xùn)練與時(shí)鐘基因以及晝夜節(jié)律的關(guān)系,為運(yùn)動(dòng)表現(xiàn)、運(yùn)動(dòng)能力的外周生物節(jié)律機(jī)制提供實(shí)驗(yàn)依據(jù)。
1 研究對(duì)象與方法
1.1 研究對(duì)象
8周齡Sprague-Dawley(SD)大鼠208只,體質(zhì)量(283.17±4.06)g,由成都達(dá)碩生物科技有限公司提供,在成都體育學(xué)院SPF級(jí)動(dòng)物實(shí)驗(yàn)室分籠飼養(yǎng),室內(nèi)溫度為22~25 ℃,濕度為60%~70%,自由飲食、飲水。實(shí)驗(yàn)室光照與黑暗時(shí)間12 h交替,光照起始時(shí)間(zeitgeber time, ZT)為8∶00,光照結(jié)束時(shí)間(ZT12)為20∶00。實(shí)驗(yàn)中動(dòng)物處置符合國(guó)家科學(xué)技術(shù)部2006年頒布的《關(guān)于善待實(shí)驗(yàn)動(dòng)物的指導(dǎo)性意見(jiàn)》相關(guān)規(guī)定,獲得成都體育學(xué)院倫理委員會(huì)批準(zhǔn)[(2021)34號(hào)]。
1.2 實(shí)驗(yàn)分組
大鼠實(shí)驗(yàn)前適應(yīng)性喂養(yǎng)1周后隨機(jī)分為空白對(duì)照(Control,C)組和運(yùn)動(dòng)(Exercise,E)組,根據(jù)運(yùn)動(dòng)72 h內(nèi)不同取材時(shí)相(ZT0、ZT6、ZT12、ZT18、ZT24、ZT30、ZT36、ZT42、ZT48、ZT54、ZT60、ZT66、ZT72),C組分為C0、C6、C12、C18、C24、C30、C36、C42、C48、C54、C60、C66、C72共13個(gè)亞組;E組分為E0、E6、E12、E18、E24、E30、E36、E42、E48、E54、E60、E66、E72共13個(gè)亞組。
1.3 運(yùn)動(dòng)方案
通過(guò)一次性下坡跑建立大鼠大負(fù)荷運(yùn)動(dòng)模型。E組經(jīng)3天適應(yīng)性喂養(yǎng)后,先采取2天適應(yīng)性運(yùn)動(dòng),具體運(yùn)動(dòng)方案:第1天:跑臺(tái)坡度0°,運(yùn)動(dòng)速度16 m/min,運(yùn)動(dòng)時(shí)間5 min;第2天:跑臺(tái)坡度0°,運(yùn)動(dòng)速度16 m/min,運(yùn)動(dòng)時(shí)間10 min;第3天不運(yùn)動(dòng)。同時(shí)C組正常喂養(yǎng)。適應(yīng)性訓(xùn)練完成后第4天清晨于6∶30開(kāi)始運(yùn)動(dòng),坡度-16°,速度16 m/min,運(yùn)動(dòng)時(shí)間90 min,跑完即刻與ZT0(08∶00)時(shí)相相同。
1.4 取材與樣本采集
按照不同時(shí)間進(jìn)行分批取材。實(shí)驗(yàn)大鼠稱(chēng)重后腹腔注射10%水合氯醛(3.5 mL/kg)麻醉,腹主動(dòng)脈取血。用有齒鑷夾起大鼠腹股溝處皮膚,剝離多余結(jié)締組織,暴露完整肌肉,并將皮膚分離至跟腱部位,用止血鉗夾住大鼠足部,將跟腱翻折暴露,手術(shù)刀劃斷跟腱并用齒鑷提起,用手術(shù)刀劃開(kāi)外層腓腸肌暴露出深層比目魚(yú)肌,冰冷生理鹽水漂洗,濾紙拭干,剔除結(jié)締組織。所有時(shí)相取右側(cè)組織置于液氮迅速冷凍后-80 ℃冰箱保存,待測(cè)RT-PCR實(shí)驗(yàn);每24 h取左側(cè)2 mm×3 mm組織塊投入2.5%戊二醛固定液,制備電鏡樣品。
1.5 透射電鏡觀察肌纖維超微結(jié)構(gòu)
采用透射電鏡(型號(hào)JEM-1400PLUS,日本電子JEOL生產(chǎn))觀察比目魚(yú)肌的肌原纖維特征,包括肌小節(jié)排列順序、肌節(jié)明/暗帶分布情況、線粒體改變情況。將骨骼肌切段后進(jìn)行樣品預(yù)固定(3%戊二醛),接著再固定(1%四氧化鋨),經(jīng)丙酮逐級(jí)脫水,按比例設(shè)置脫水劑的濃度(在100%濃度中更換3次),處理后的骨骼肌樣品使用脫水劑,后通過(guò)環(huán)氧樹(shù)脂包埋,制備約50 nm切片后,漂浮于刀槽液面上,再撈至銅網(wǎng),分別通過(guò)醋酸鈾和枸櫞酸鉛雙染色10~15 min和1~2 min,透射電鏡觀察采圖。
1.6 實(shí)時(shí)熒光定量PCR檢測(cè)核心時(shí)鐘基因mRNA表達(dá)
將100 mg組織在液氮中磨碎,加入1 mL已預(yù)冷的TRIzol,依次采用氯仿、異丙醇、75%乙醇經(jīng)反復(fù)離心分離提純樣本總RNA,經(jīng)超微量紫外分光光度計(jì)和瓊脂糖凝膠電泳檢測(cè)濃度和純度后,將RNA逆轉(zhuǎn)錄合成cDNA。
采用PIKORed 96(ThermoFisher生產(chǎn),美國(guó))實(shí)時(shí)熒光定量PCR測(cè)定Bmal1、Clock、Cry1、Per1的mRNA表達(dá)水平,PCR反應(yīng)體系與反應(yīng)條件見(jiàn)表1、表2。所有引物均交由上海生工生物工程技術(shù)服務(wù)有限公司設(shè)計(jì)合成,并以ULTRAPAGE純化,引物序列見(jiàn)表3。使用Thermo Scientific PikoReal軟件分析PCR過(guò)程各檢測(cè)樣本的CT(Threshold cycle)值。通過(guò)2-△△CT計(jì)算X相對(duì)mRNA表達(dá)水平。

表1 PCR反應(yīng)體系Table 1 PCR Reaction System

表2 PCR反應(yīng)條件Table 2 PCR Reaction Conditions
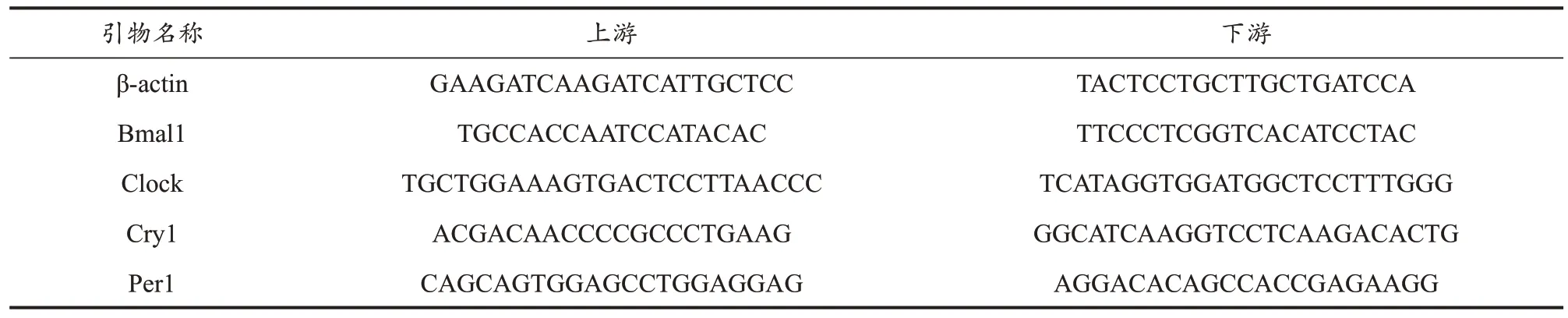
表3 PCR引物序列Table 3 PCR Primer Sequence
1.7 數(shù)據(jù)統(tǒng)計(jì)與分析
所得數(shù)據(jù)以平均數(shù)±標(biāo)準(zhǔn)差(M±SD)表示。使用GraphPad Prism 8.0統(tǒng)計(jì)軟件進(jìn)行數(shù)據(jù)處理。對(duì)所得數(shù)據(jù)進(jìn)行正態(tài)性檢驗(yàn),比較符合正態(tài)分布的兩組獨(dú)立樣本時(shí)采用Student’st檢驗(yàn),方差齊性時(shí)采用LSD檢驗(yàn),方差不齊時(shí)用Welch’s校正檢驗(yàn),P<0.05表示差異具有顯著性。
應(yīng)用余弦分析軟件circacompare (R package) 獲取擬合余弦曲線參數(shù),其擬合的余弦函數(shù)方程為Y=Mesor+Amplitude*cos(time_Radians-φ),其中Mesor為基線/中值;Amplitude為節(jié)律震蕩的振幅;time_Radians為時(shí)間所對(duì)應(yīng)的弧度值;φ為峰值相位。
2 研究結(jié)果
2.1 各組骨骼肌核心時(shí)鐘基因表達(dá)的節(jié)律分析
C組擬合余弦函數(shù)曲線顯示,Bmal1 mRNA于ZT0.25處于峰值,隨后下降到ZT12.25為谷值,然后回升至ZT24完成一個(gè)周期節(jié)律振蕩;Clock mRNA于ZT5.43處于峰值,隨后下降到ZT17.43為谷值,然后回升至ZT24完成一個(gè)周期節(jié)律振蕩;Cry1 mRNA于ZT1.20處于谷值,隨后上升到ZT13.20為峰值,然后回落至ZT24完成一個(gè)周期節(jié)律振蕩;Per1 mRNA于ZT0.42處于谷值,隨后上升到ZT12.42為峰值,然后回落至ZT24完成一個(gè)周期節(jié)律振蕩。在之后2天中, Bmal1、Clock、Cry1、Per1 mRNA的變化趨勢(shì)與0~24 h相似,72 h內(nèi)共呈現(xiàn)3個(gè)完整近日節(jié)律(圖1~4,表4)。
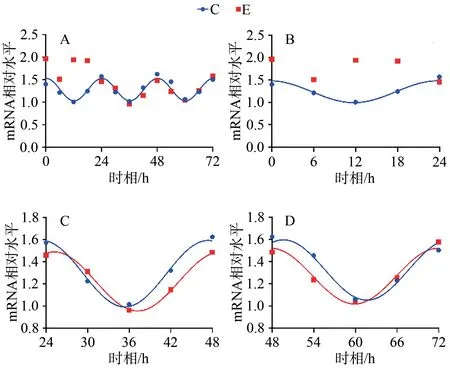
圖1 Bmal1 mRNA 表達(dá)及擬合余弦函數(shù)節(jié)律性變化Figure 1. Bmal1 mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
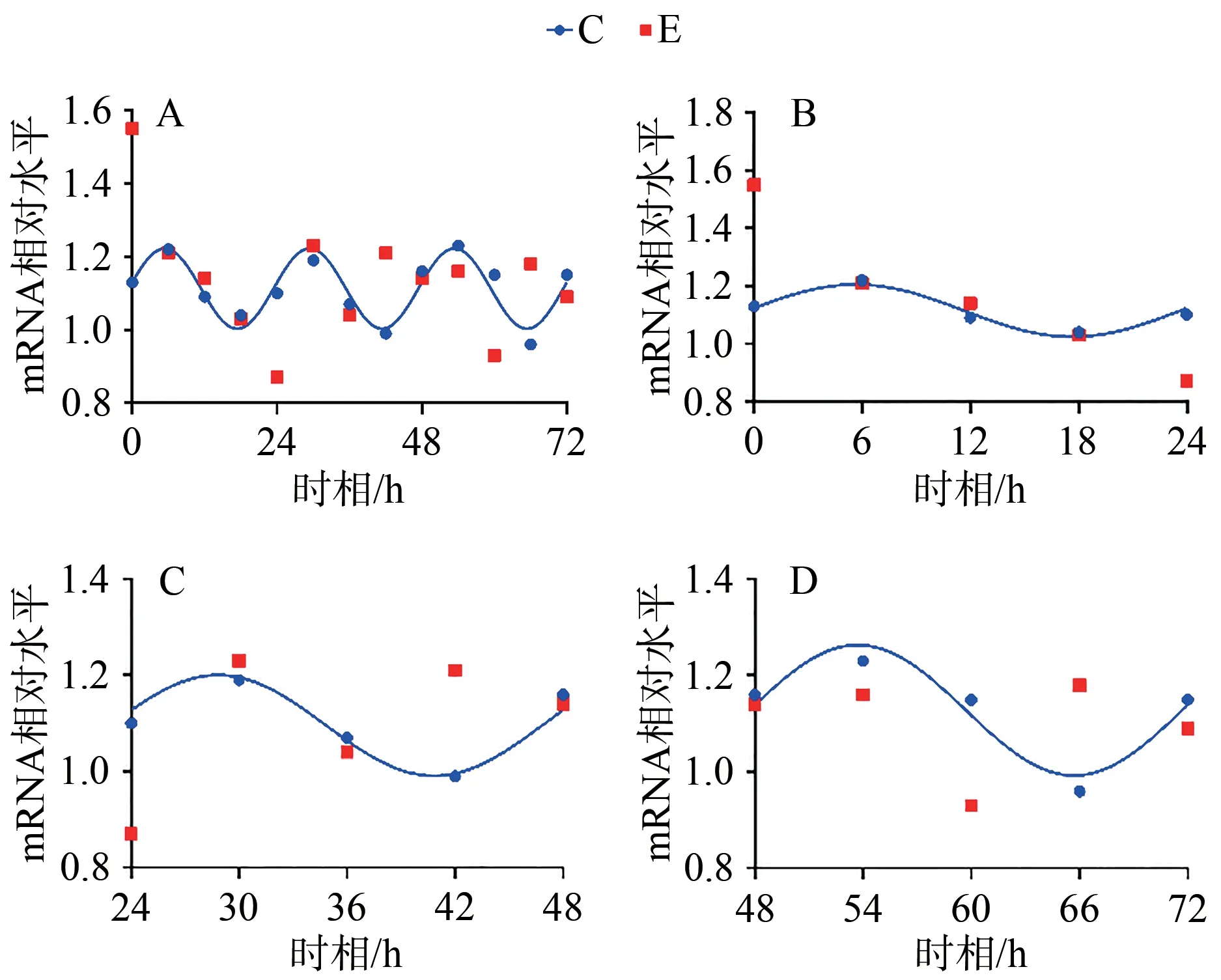
圖2 Clock mRNA 表達(dá)及擬合余弦函數(shù)節(jié)律性變化Figure 2. Clock mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
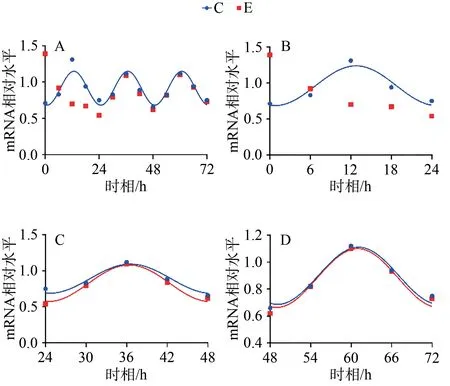
圖3 Cry1 mRNA 表達(dá)及擬合余弦函數(shù)節(jié)律性變化Figure 3. Cry1 mRNA Expression and the Rrhythmic Changes of the Fitted Cosine Function
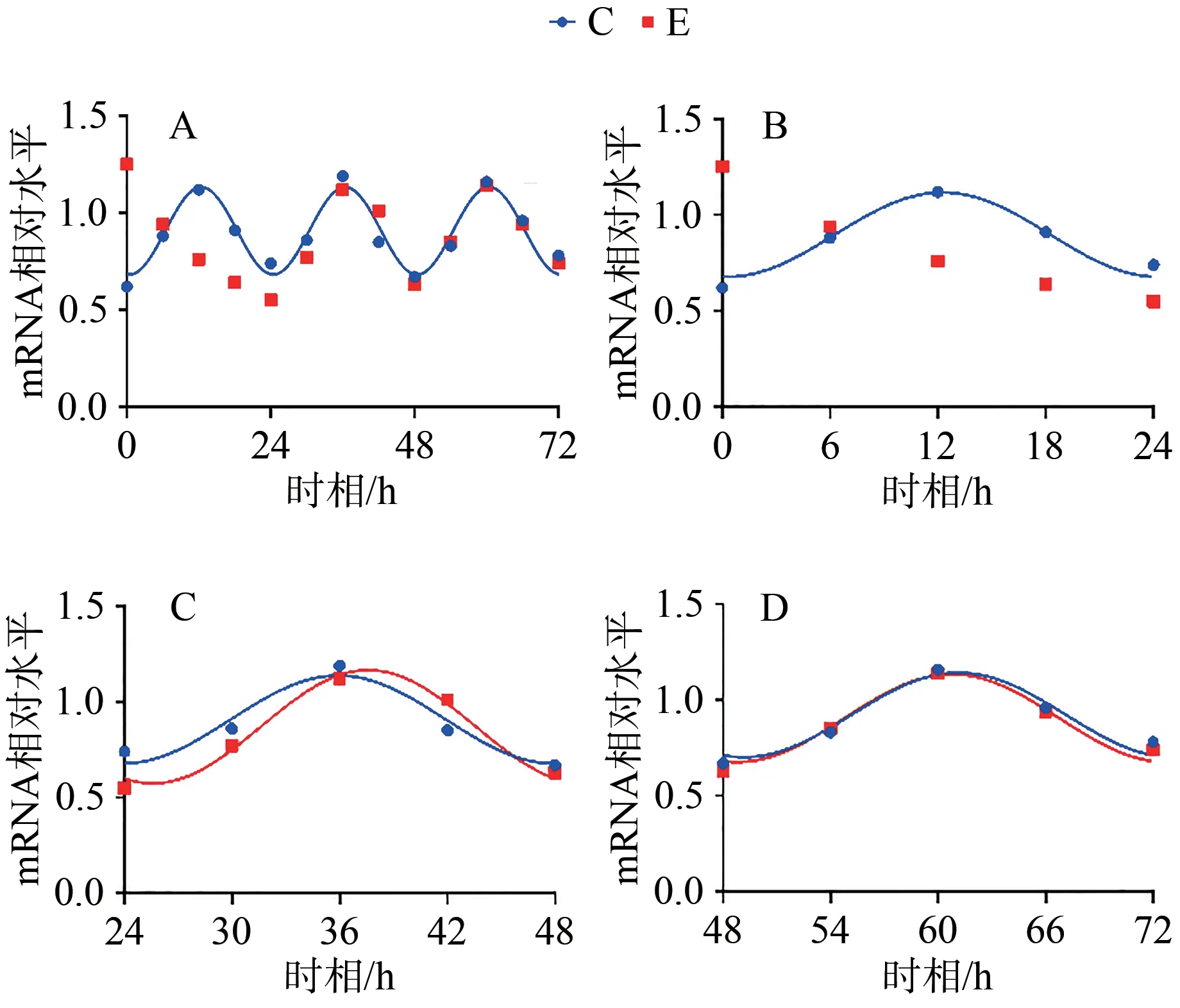
圖4 Per1 mRNA 表達(dá)及擬合余弦函數(shù)節(jié)律性變化Figure 4. Per1 mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
E組擬合余弦函數(shù)曲線顯示,Bmal1 mRNA在0~24 h近日節(jié)律消失,24~48 h與48~72 h恢復(fù)近日節(jié)律;Clock mRNA在0~72 h近日節(jié)律消失;Cry1 mRNA在0~24 h近日節(jié)律消失,24~48 h恢復(fù)近日節(jié)律;Per1 mRNA在0~24 h近日節(jié)律消失,24~48 h恢復(fù)近日節(jié)律(圖1~4,表4)。
2.2 各組對(duì)應(yīng)時(shí)相骨骼肌核心時(shí)鐘基因表達(dá)的比較分析
統(tǒng)計(jì)結(jié)果顯示,E組Bmal1 mRNA呈現(xiàn)先升高,后趨于正常的狀態(tài),在0、6、12和18 h顯著高于C組對(duì)應(yīng)時(shí)相(P<0.05);Clock mRNA變化趨勢(shì)與Bmal1 mRNA相似,在0 h差異顯著(P<0.05);Cry1 mRNA在0、12 h顯著低于C組對(duì)應(yīng)時(shí)相(P<0.05);Per1 mRNA變化趨勢(shì)與Cry1 mRNA基本相同,在0、12 h顯著低于C組對(duì)應(yīng)時(shí)相(P<0.05;表4,圖5)。
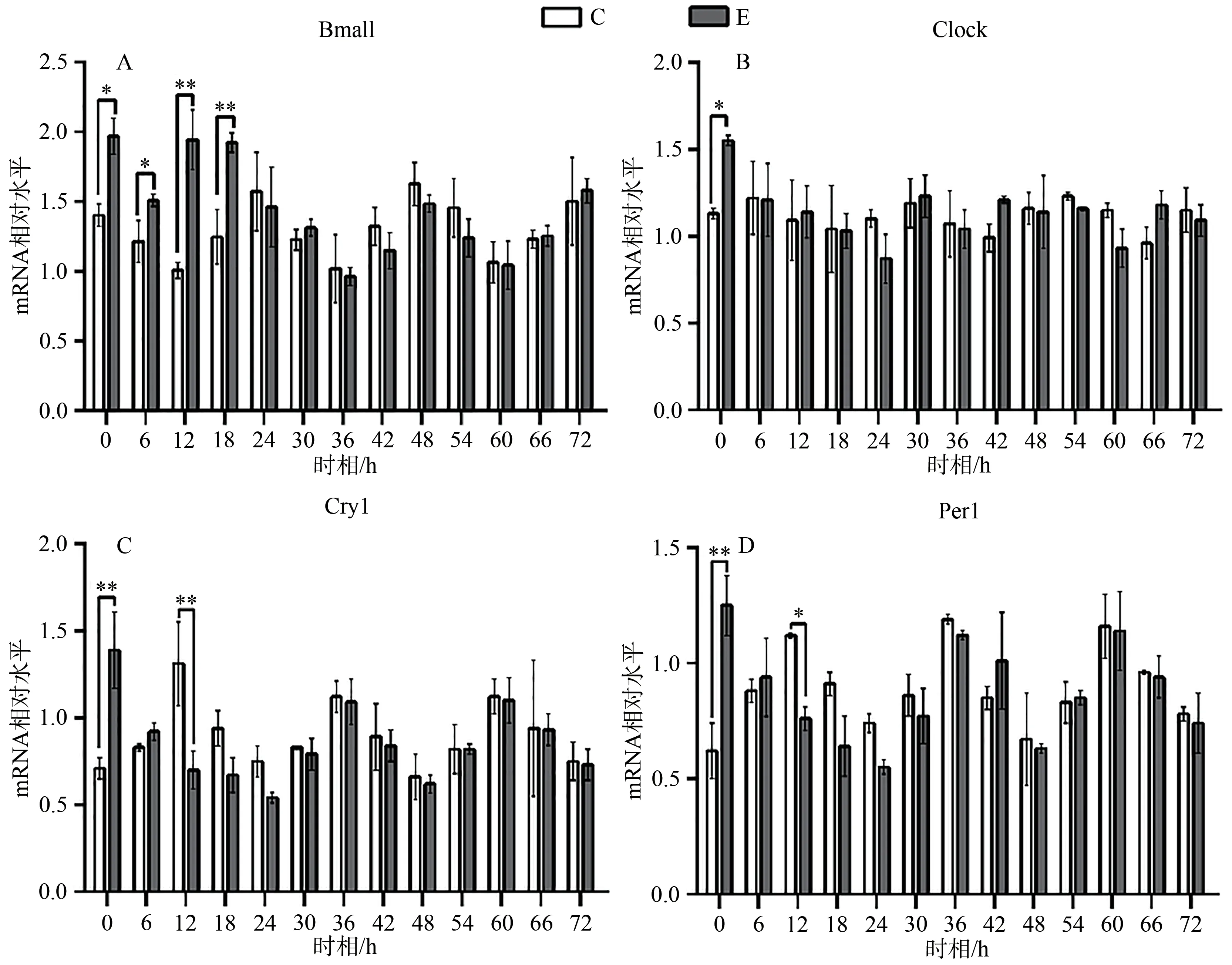
圖5 E組與C組同一時(shí)相Bmal1、Clock、Cry1、Per1 mRNA的相對(duì)表達(dá)變化Figure 5. Relative Expression Changes of Bmal1, Clock, Cry1 and Per1 mRNA in Group E and Group C at the Same Timepoint
2.3 各組骨骼肌纖維超微結(jié)構(gòu)變化
透射電鏡下觀察肌纖維超微結(jié)構(gòu)(圖6)發(fā)現(xiàn),C組在0、24、48和72 h各時(shí)相肌原纖維分布均勻,A帶與I帶分界清楚、排列規(guī)則,Z線清晰,線粒體排列整齊、形態(tài)完整。E組0、24和48 h出現(xiàn)不同程度的Z線加寬、斷裂,A帶與I帶分界不清,線粒體腫脹并聚集在肌膜下,72 h雖有所緩解,但仍未完全恢復(fù)。

圖6 各組大鼠骨骼肌纖維超微結(jié)構(gòu)Figure 6. Ultrastructure of Skeletal Muscle Fibers in Each Group of Rats
3 分析討論
晝夜時(shí)間(circadian time,CT)0時(shí),Bmal1和Clock形成異二聚體,作用于Cry、Per等靶基因的 E 反應(yīng)元件(Ebox,CACGTG)并啟動(dòng)轉(zhuǎn)錄過(guò)程,Cry基因的表達(dá)產(chǎn)物和Per基因的表達(dá)產(chǎn)物可形成復(fù)合物,在細(xì)胞核中不斷積蓄并反饋抑制Bmall/Clock異二聚體驅(qū)動(dòng)的E-box表達(dá)。CT12—CT24期間,Per和Cry的mRNA水平下降,現(xiàn)有Per/Cry復(fù)合物降解,以上過(guò)程進(jìn)行約24 h周期循環(huán),形成了生物鐘調(diào)控網(wǎng)絡(luò)的TTFL(黃增浩 等,2020)。本研究對(duì)未施加干預(yù)的對(duì)照組SD大鼠骨骼肌Bmal1和Clock、Per和Cry的mRNA進(jìn)行檢測(cè),并獲取擬合余弦曲線參數(shù)分析其變化趨勢(shì),發(fā)現(xiàn)72 h內(nèi)上述基因均呈現(xiàn)3個(gè)完整近日節(jié)律周期,即晝夜節(jié)律振蕩。進(jìn)一步分析其振蕩曲線的峰值與谷值,發(fā)現(xiàn)Bmal1在CT0—CT12期間由波峰降至波谷,在CT12—CT24期間回升至波峰,Clock的表達(dá)趨勢(shì)與Bmal1基本一致,而Per1和Cry1的擬合曲線則與Bmal1和Clock呈現(xiàn)反向振蕩,以上4個(gè)基因周期性特異表達(dá)形成的振蕩規(guī)律與生物節(jié)律核心分子機(jī)制——TTFL相吻合。經(jīng)轉(zhuǎn)錄組學(xué)研究發(fā)現(xiàn),骨骼肌中3.4%~16.0%的基因是以節(jié)律性方式表達(dá),其表達(dá)的Baml1、Rev-erbα、Clock等時(shí)鐘基因不僅可維持晝夜節(jié)律穩(wěn)定調(diào)節(jié),而且通過(guò)激活相關(guān)信號(hào)轉(zhuǎn)導(dǎo)通路或其他作用途徑,參與調(diào)控骨骼肌機(jī)能(Perrin et al., 2018)。本研究基于骨骼肌時(shí)鐘基因特異性節(jié)律表達(dá),主要檢測(cè)和分析了Bmal1/Clock、Cry1/Per1 mRNA晝夜變化趨勢(shì),實(shí)驗(yàn)結(jié)果進(jìn)一步佐證了骨骼肌組織作為外周時(shí)鐘的關(guān)鍵作用,表明骨骼肌表達(dá)的核心時(shí)鐘基因參與維持晝夜節(jié)律的穩(wěn)定輸出。
骨骼肌不僅作為外周組織表達(dá)時(shí)鐘基因維持生物節(jié)律,還受到時(shí)鐘基因調(diào)控參與各項(xiàng)生理活動(dòng)。運(yùn)動(dòng)表現(xiàn)、運(yùn)動(dòng)能力與時(shí)間密切相關(guān)(黃華生 等, 2016; Faull et al.,2015),有研究發(fā)現(xiàn)跨時(shí)區(qū)產(chǎn)生的時(shí)差變化可影響運(yùn)動(dòng)表現(xiàn)能力,與向西跨經(jīng)度旅行相比,向東跨經(jīng)度旅行可使人體在間歇性短跑運(yùn)動(dòng)中表現(xiàn)下降、疲勞感更強(qiáng)(Fowler et al.,2017)。時(shí)鐘基因是運(yùn)動(dòng)表現(xiàn)差異的原因之一,人體實(shí)驗(yàn)研究發(fā)現(xiàn),以股四頭肌為測(cè)試對(duì)象,受試者在進(jìn)行45 min抗阻訓(xùn)練后6 h和18 h,Bmal1、Cry1和Per2的mRNA表達(dá)顯著升高(Zambon et al., 2003);70 min、70% V.O2max強(qiáng)度的自行車(chē)運(yùn)動(dòng)后4 h和8 h,股外側(cè)肌Bmal1 mRNA表達(dá)顯著升高,而Per1 mRNA表達(dá)顯著降低(Popov et al., 2018);15 min 80% V.O2max強(qiáng)度的自行車(chē)運(yùn)動(dòng)后60 min,股外側(cè)肌中Per1和Per2的表達(dá)增加1.5倍,而B(niǎo)mal1和Cry1沒(méi)有變化(Small et al., 2020)。可見(jiàn),不同形式、不同強(qiáng)度以及不同時(shí)間的一次運(yùn)動(dòng)后,人體骨骼肌中核心生物鐘基因的表達(dá)變化并不一致(嚴(yán)露 等,2021)。深入了解運(yùn)動(dòng)與骨骼肌時(shí)鐘基因節(jié)律振蕩的關(guān)系,有利于深入探究肌肉運(yùn)動(dòng)能力的時(shí)間調(diào)節(jié)機(jī)制、擇時(shí)運(yùn)動(dòng)如何增強(qiáng)運(yùn)動(dòng)效果以及過(guò)度訓(xùn)練如何影響機(jī)能狀態(tài)。本研究采用可誘發(fā)骨骼肌微損傷的一次離心運(yùn)動(dòng)經(jīng)典模型,試圖明確大負(fù)荷運(yùn)動(dòng)能否影響骨骼肌時(shí)鐘基因固有節(jié)律表達(dá)。實(shí)驗(yàn)結(jié)果發(fā)現(xiàn),Bmal1、Cry1、Per1在運(yùn)動(dòng)后第1天(0~24 h)均出現(xiàn)近日節(jié)律消失,第2天(24~48 h)和第3天(48~72 h)基本恢復(fù),相較而言,Clock最為敏感,72 h內(nèi)的3個(gè)近日節(jié)律均受到破壞,表明一次大負(fù)荷離心運(yùn)動(dòng)可誘發(fā)骨骼肌核心時(shí)鐘基因節(jié)律振蕩紊亂,但因具體分子鐘的不同,其周期性變化呈現(xiàn)特有的振幅和相位表達(dá)。
晝夜節(jié)律時(shí)鐘基因大都存在亞型而顯示功能冗余,如Cry與Per家族中Cry1、Cry2、Per1、Per2,若敲除單個(gè)基因并不會(huì)出現(xiàn)明顯表型變化,而敲除Cry1/2、Per1/Per2雙重基因才能破壞相應(yīng)基因功能(Aoyama et al., 2017)。Bmal1作為一種非冗余時(shí)鐘基因,敲除后可導(dǎo)致生物鐘完全紊亂(Schiaffino et al., 2016)。Bmal1/Clock異二聚體、Cry/Per復(fù)合物是TTEL核心環(huán)路的關(guān)鍵結(jié)構(gòu),因此,需要在整體分析其作用效應(yīng)的同時(shí),進(jìn)一步分析各分子的具體變化。本研究中,一次大負(fù)荷運(yùn)動(dòng)后Bmal1在0~24 h近日節(jié)律消失,且無(wú)論是光照還是運(yùn)動(dòng)刺激后,均在CT0左右即出現(xiàn)顯著變化,而Clock在0~72 h內(nèi)3個(gè)近日節(jié)律全部消失,推測(cè)Bmal1可能作為始動(dòng)因子,首先影響B(tài)mal1/Clock異二聚體結(jié)構(gòu),進(jìn)而調(diào)控Cry/Per復(fù)合物,形成核心環(huán)路4個(gè)分子鐘的周期性表達(dá)變異,在此過(guò)程中,Clock可能具有對(duì)運(yùn)動(dòng)更為敏感的特異性。然而,大負(fù)荷運(yùn)動(dòng)后時(shí)鐘基因出現(xiàn)上述節(jié)律振蕩紊亂的具體作用機(jī)制及其帶來(lái)的后續(xù)效應(yīng),仍需要進(jìn)一步探究。
一次大負(fù)荷運(yùn)動(dòng)模型可誘發(fā)運(yùn)動(dòng)性骨骼肌損傷,有研究認(rèn)為骨骼肌質(zhì)量與肌纖維結(jié)構(gòu)、功能受到時(shí)鐘基因調(diào)控。Clock-/-小鼠可致骨骼肌肌絲結(jié)構(gòu)破壞,單個(gè)肌纖維肌力水平下降(Andrews et al., 2010);Bmal1-/-小鼠骨骼肌粗細(xì)肌絲超微結(jié)構(gòu)破壞,線粒體腫脹、體積減小(Dyar et al.,2016),但具體途徑尚不明確,是否和生物節(jié)律中斷或時(shí)鐘基因振蕩紊亂相關(guān),還是中樞或其他外周組織的繼發(fā)效應(yīng)所致有待進(jìn)一步探索。本研究在每個(gè)近日節(jié)律周期始末觀察肌纖維超微結(jié)構(gòu),同時(shí)分析比較E組與C組對(duì)應(yīng)時(shí)相基因表達(dá)差異,結(jié)果發(fā)現(xiàn),在近日節(jié)律表達(dá)均受到破壞的0~24 h,E組相較C組出現(xiàn)顯著變化,肌纖維超微結(jié)構(gòu)損傷在0、24 h也較為明顯,24~48 h時(shí)Clock節(jié)律未恢復(fù)且Bmal1節(jié)律發(fā)生偏移,而48 h時(shí)相的肌纖維超微結(jié)構(gòu)仍有損傷現(xiàn)象,在48~72 h,隨著大部分核心時(shí)鐘基因近日節(jié)律的恢復(fù),肌纖維超微結(jié)構(gòu)損傷也明顯減輕,表明時(shí)鐘基因以及骨骼肌生物節(jié)律可參與運(yùn)動(dòng)誘發(fā)的肌纖維結(jié)構(gòu)改變,機(jī)體生物鐘與外界生物鐘失去同步性有可能加速運(yùn)動(dòng)性骨骼肌損傷的發(fā)展。綜上,本研究通過(guò)檢測(cè)Bmal1/Clock、Cry1/Per1的mRNA表達(dá),分析時(shí)相變化及其與肌纖維結(jié)構(gòu)損傷的關(guān)系,初步闡釋了大負(fù)荷運(yùn)動(dòng)后骨骼肌核心時(shí)鐘基因的節(jié)律性振蕩趨勢(shì),為運(yùn)動(dòng)員對(duì)抗或減輕大運(yùn)動(dòng)量訓(xùn)練后時(shí)鐘基因變化對(duì)機(jī)體的影響,科學(xué)合理安排訓(xùn)練計(jì)劃并預(yù)防運(yùn)動(dòng)損傷,以及應(yīng)對(duì)跨時(shí)區(qū)、洲際參賽引起的晝夜節(jié)律紊亂等問(wèn)題提供了實(shí)驗(yàn)依據(jù)與參考。
4 結(jié)論
大鼠骨骼肌核心時(shí)鐘基因Bmal1/Clock、Cry1/Per1呈現(xiàn)晝夜節(jié)律振蕩趨勢(shì),一次大負(fù)荷運(yùn)動(dòng)可誘發(fā)上述時(shí)鐘基因節(jié)律紊亂,其時(shí)相變化與骨骼肌纖維超微結(jié)構(gòu)損傷相一致。

- 中國(guó)體育科技的其它文章
- 合并軟骨損傷的前交叉韌帶損傷特征的研究進(jìn)展
- 加快建設(shè)體育強(qiáng)國(guó)背景下行為經(jīng)濟(jì)學(xué)促進(jìn)決策優(yōu)化研究
- Notch信號(hào)通路介導(dǎo)運(yùn)動(dòng)對(duì)抑郁的調(diào)控機(jī)制
——基于整合生物學(xué)視角 - 加壓訓(xùn)練對(duì)中老年人下肢肌肉力量及肥大效應(yīng)的Meta分析
- 睡眠時(shí)長(zhǎng)與中國(guó)6~19歲兒童青少年肥胖風(fēng)險(xiǎn)的關(guān)系:基于系統(tǒng)綜述與Meta分析
- 基于速度的力量訓(xùn)練中不同速度丟失閾值(10% vs 20%)對(duì)大學(xué)生運(yùn)動(dòng)員下肢爆發(fā)力的影響