大腸桿菌L-蘇氨酸脫氫酶的高效表達及其酶學性質分析
2023-10-21 03:10:26劉欣欣史紅玲姚倫廣唐存多
食品科學 2023年18期
關鍵詞:分析
劉欣欣,王 瑤,史紅玲,姚倫廣,王 賢,唐存多,,3,*
(1.南陽師范學院生命科學與農業工程學院,河南 南陽 473061;2.賒店老酒股份有限公司博士后創新實踐基地,河南 南陽 473300;3.河南農業大學食品科學技術學院,河南 鄭州 450002)
2,5-二甲基吡嗪(2,5-dimethylpyrazine, 2,5-DMP)是一種雜環含氮化合物,廣泛存在于各種食品中,是食品中重要的香氣化合物[1]。由于2,5-DMP具有增香作用,它作為食品工業中的食品添加劑受到了廣泛關注。天然的2,5-DMP常存在于熱處理后原料種子[2]、烤牛肉[3]、白醋[4]、酒[5-6]、醬油[7]、咖啡豆[8-9]、烤花生[10-11]、烏龍茶[12]以及發酵大豆[13]中,具有炒花生香氣和巧克力、奶油風味,是食品工業中不可缺少的調味添加劑。同時,2,5-DMP還被應用于制藥工業,是生產降糖類藥物和抗脂類藥物的重要底物,如5-甲基吡嗪-2-羧酸是合成抗脂分解藥物中間體的合成原料[14],常用于合成抗高血壓類藥物阿莫西林、降血糖血脂類藥物格列吡嗪以及抗結核藥物5-甲基吡嗪-2-羧酸甲酯等[15-16]。
L-蘇氨酸是一種必需氨基酸,可用于食品增香劑、醫藥、飼料添加等方面[17]。目前,安全綠色且廣泛使用的生產方法是微生物發酵法[18]。近年來,L-蘇氨酸的生產過熱,導致出現了產能嚴重過剩的窘境[19]。因此,L-蘇氨酸的轉產增值迫在眉睫,其途徑主要有以下幾種:一是以L-蘇氨酸為底物,經過多酶級聯反應進行脫氨和還原生成L-氨基丁酸,同時還原反應所消耗的NADH通過甲酸脫氫酶再生[20]。二是以L-蘇氨酸為底物,利用L-蘇氨酸脫氫酶(L-threonine dehydrogenase,L-TDH)使L-蘇氨酸脫氫生成L-2-氨基乙酰乙酸,經過一系列自發反應生成2,5-DMP。三是L-蘇氨酸在蘇氨酸醛縮酶的作用下直接生成甘氨酸。本研究擬挖掘鑒定出高活性的L-TDH,試圖以L-蘇氨酸為底物采用全細胞催化法生產2,5-DMP,實現L-蘇氨酸的轉產增值,力爭解決蘇氨酸產能過剩問題[12,14]。在大腸桿菌(Escherichia coli)中,2,5-DMP可以由L-蘇氨酸天然合成,全細胞催化途徑中的催化過程為L-蘇氨酸在L-TDH的作用下轉化為L-2-氨基乙酰乙酸[21],L-TDH是催化過程中的關鍵酶[22]。
已有文獻報道,來源于E.coli的L-TDH借助pET28a載體在E.coli中實現了可溶性表達,其比活力約為132 mU/mg。曹艷麗等[19]經枯草芽孢桿菌外源表達了來源不同的L-TDH,結果表明,來源于E.coliK-12的L-TDH在枯草芽孢桿菌中更有利于L-蘇氨酸合成2,5-DMP,但其L-TDH活力僅為0.024 IU,比活力為0.15 IU/mg。本研究擬以E.coli為出發菌株,從中調取L-TDH基因,再借助pACYCDuet-1質粒強化其在E.coliBL21(DE3)的表達,并對其酶學性質進行分析,旨在為提高蘇氨酸轉化為L-2-氨基乙酰乙酸效率提供參考。
1 材料與方法
1.1 材料與試劑
1.1.1 試劑
限制性內切酶BamH I和Hind III New England Biolabs(北京)公司;L-蘇氨酸 美國Sigma-Aldrich公司;2,5-DMP、2,4-二硝基氟苯 上海阿拉丁生化科技股份有限公司;NAD+日本Chem-Station公司;異丙基-β-D-硫代半乳糖苷(isopropyl-beta-D-thiogalactoside,IPTG)Master of Bioactive Molecules(北京)公司;Prime STAR HS (Premix)、DL 2000 DNA Marker、rTaqDNA聚合酶、PrimeScript? II 1st Strand cDNA Synthesis Kit、DNA連接試劑盒Ver.2.1 寶生物工程(大連)有限公司;甲醇、甲酸(均為色譜純) 天津科密歐化學試劑有限公司;其他試劑均為國產分析純。
1.1.2 菌株與質粒
本研究所涉及到的菌株和質粒如表1所示,重組質粒的構建如圖1所示。
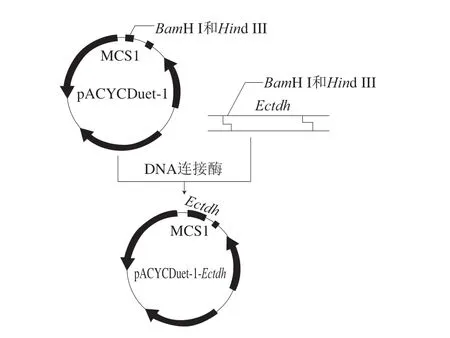
圖1 重組質粒pACYCDuet-1-Ectdh的構建Fig.1 Construction of the recombinant plasmid pACYCDuet-1-Ectdh

表1 本研究用到的菌株和質粒Table 1 Strains and plasmids used in this study
1.2 儀器與設備
核酸電泳儀、Hypersil C18柱 美國Thermo Fisher公司;UNIVERSAL Hood凝膠成像系統、PowerPacTMHC/Mini-PROTEAN?蛋白電泳系統 美國Bio-Rad公司;VCX 130型超聲破碎儀 美國Sonics and Materials公司;UV1000紫外-可見分光光度計 梅特勒-托利多科技(中國)有限公司;EC 2006型高效液相色譜(high performance liquid chromatography,HPLC)系統 大連依利特分析儀器有限公司。
1.3 方法
1.3.1 重組菌株的構建及序列分析
從EMBL’s European Bioinformatics Institute(https://www.ebi.ac.uk/)數據庫中搜索到一個E.coli來源的L-TDHEcTDH(ACI75701.1)。根據此序列設計擴增E c t d h的特異性引物E c t d h-F:CGCGGATCCGATGAAAGCGTTATCCAAACTG( 含B a mH I 酶 切 位 點) 和E c t d h- R :CCCAAGCTTTTAATCCCAGCTCAGAATAAC(含Hind III酶切位點),委托蘇州泓訊生物技術有限公司進行合成。以E.coliBL21(DE3)基因組DNA為模板,利用Ectdh-F和Ectdh-R引物進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增Ectdh的編碼基因,再利用限制性核酸內切酶BamH I和Hind III對目的基因及載體pACYCDuet-1分別進行雙酶切,然后連接轉化進入E.coliBL21(DE3)感受態細胞,經氯霉素抗性篩選和測序鑒定獲得E.coliBL21(DE3)/ pACYCDuet-1-Ectdh重組菌。將上述重組菌連續在無抗LB培養基上進行培養,然后接種至含氯霉素抗性的LB培養基上培養,觀察生長情況,通過氯霉素抗性篩選考察重組菌攜帶質粒的遺傳穩定性[23]。基于測序結果,通過Jalview對該酶進行多序列比對。
1.3.2 重組E.coli誘導
重組E.coli的誘導表達采用低溫、低濃度誘導劑的方法。分別將E.coliBL21(DE3)/pACYCDuet-1、E.coliBL21(DE3)/pACYCDuet-1-Ectdh單菌落接種至5 mL含100 μg/mL氯霉素的LB液體培養基中,37 ℃、200 r/min振蕩培養14 h,以2%接種量將過夜培養的菌液轉接至100 mL新鮮LB液體培養基中,37 ℃、200 r/min培養約2 h至菌體細胞密度(OD600nm)為1.0,加入1 mol/L IPTG至終濃度為0.1 mmol/L,16 ℃、200 r/min誘導培養20 h。將100 mL菌液于8 000 r/min、4 ℃離心5~10 min,收集菌體,將菌體用20 mmol/L Tris-HCl重懸后進行超聲破碎,破碎后收集上清液,上清液通過鎳瓊脂糖柱上進行純化,利用十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)進行分析[24]。
1.3.3 酶活力測定
誘導表達后的重組菌進行超聲破碎,離心取純化后的上清液,參照文獻[19]并進行略微改動測定重組酶活力。以L-蘇氨酸為底物測定EcTDH活力,總的1 mL反應體系包含0.76 mL Tris-HCl(76 mmol/L,pH 7.8)、0.08 mLL-蘇氨酸(8 mmol/L)、0.06 mL NAD+溶液(3 mmol/L)混勻,放入35 ℃金屬浴中預熱5 min,加入0.1 mL酶液,測定1 min內OD340nm的變化值。酶活力定義為1 min內生成1 μmol NADH所需的酶量為1 IU[25]。
1.3.4 重組酶的溫度特性
參照1.3.3節方法,分別在20~60 ℃(以5 ℃為間隔)下測定重組酶活力以獲得重組酶的最適反應溫度。將重組酶分別在35~55 ℃保溫0~120 min(以10 min為間隔),在最適反應溫度下測定各自的殘留酶活力,以冰浴保存各時間段的殘留酶活力為100%,考察重組酶的溫度穩定性。
1.3.5 重組酶的pH值特性
參照1.3.3節方法,控制在最適反應溫度下,分別在pH 6.5~10.5(以0.5為間隔)下測定重組酶活力以獲得重組酶的最適反應pH值。將重組酶分別在pH 6.5~10.5(以0.5為間隔)的緩沖液中放置在最適反應溫度下保溫1.5 h,測定各自的殘留酶活力,以未經處理的酶液為100%,考察重組酶的pH值穩定性。
1.3.6 重組酶的動力學參數
根據已報道的方法稍加修改,測定重組TDH的動力學參數,確定TDH底物的特異性[26-27],反應系統中NAD+濃度為0.5 mmol/L。最佳反應條件下,在4~14 mmol/L不同L-蘇氨酸濃度下測定重組酶的催化活性。每次測定重復3 次。同樣地,固定L-蘇氨酸濃度為10 mmol/L,將NAD+濃度設置為0.5~1.75 mmol/L,測定重組酶的催化活性,每次測定3 次,考察對NAD+的動力學參數。所有計算均由Origin 9.0執行,利用Origin 9.0軟件進行非線性擬合,得出重組酶對底物的Km、Kcat和Vmax。
1.3.7 重組E.coli全細胞轉化L-蘇氨酸合成2,5-DMP
在250 mL三角瓶中構建50 mL的反應體系,體系包括:50 mL Tris-HCl-NaCl緩沖液、85 mmol/LL-蘇氨酸、0.75 mmol/L NAD+、50 mg凍干的重組E.coli和適量玻璃珠,并于30 ℃、220 r/min振蕩反應,在0~36 h內每12 h取1 mL轉化液于60 ℃金屬浴鍋中加熱10 min終止反應,于12 000 r/min離心1 min,取上清液用0.22 μm濾膜進行過濾,最后用HPLC法檢測2,5-DMP含量,并繪制反應進程曲線。
1.3.8 底物和產物的檢測
對底物L-蘇氨酸、產物2,5-DMP進行普通C18柱的HPLC分析,HPLC色譜柱為Thermo Hypersil C18柱,檢測波長為275 nm,柱溫25 ℃,流動相A(體積分數0.1%甲酸)與流動相B(甲醇),按照表2進行梯度洗脫,流速為0.2 mL/min,運行時間10 min。底物L-蘇氨酸HPLC檢測方法通過柱前衍生測定[28-29]。

表2 HPLC梯度洗脫Table 2 HPLC gradient elution program
1.3.9 產物2,5-DMP的定量分析
將2,5-DMP標準品用超純水分別配成1.82、2.27、3.03、4.55、9.10 mmol/L的溶液,經0.22 μm微孔濾器過濾后,取10 μL進行HPLC分析,獲得各個濃度2,5-DMP的峰面積,進行線性擬合獲得回歸方程及相關系數。HPLC色譜柱為Thermo Hypersil C18柱,檢測波長275 nm,柱溫25 ℃,流動相為體積分數0.1%的甲酸和色譜級甲醇,流速0.2 mL/min,進行梯度洗脫。待檢樣品產物按照同樣的色譜條件進行HPLC分析,根據測得的峰面積可以由回歸方程計算出相應的樣品濃度。
1.3.10 常用網站及軟件
EMBL’s European Bioinformatics Institute(https://www.ebi.ac.uk/)數據庫和Basic Local Alignment Search Tool(https://blast.ncbi.nlm.nih.gov/Blast.cgi):蛋白質、基因序列的搜索;DNAMAN軟件:用于目的基因序列及限制性酶切位點分析和測序結果拼接;Origin 9.0軟件:用于數據統計及分析;Jalview軟件:用于酶的多序列比對;NCBI Autodock:用于分子對接。
1.4 數據統計與分析
圖片均用PDF-Xchange Viewer軟件進行清晰度調節,用Adobe Photoshop CS 8.0軟件進行色階、對比度調節及標注等處理。簡單的數據處理及分析均借助Origin 9.0進行。
2 結果與分析
2.1 重組E. coli的構建
以E.coli的全基因組DNA為模板,Ectdh-F和Ectdh-R為引物進行PCR擴增,擴增產物進行瓊脂糖凝膠電泳,如圖2A所示,在約1 100 bp出現了明顯的特異性條帶,PCR產物片段與預期理論長度基本一致,與已報道的來源于E.coliK-12的TDH序列相似度約為77%[19]。將PCR產物用PCR產物純化試劑盒進行純化,用限制性核酸內切酶BamH I和Hind III進行雙酶切,將酶切產物連接至經相同的限制性酶進行同樣雙酶切的pACYCDuet-1載體,轉化至E.coliBL21(DE3)感受態細胞,再經氯霉素抗性篩選和雙酶切驗證,挑取重組子,用通用引物ACYCDuetUP1 Primer和DuetDOWN1 Primer進行PCR檢測,結果如圖2B所示,片段長度約1 100 bp,大小與其預期條帶大小相符。將經PCR檢測和雙酶切驗證的重組子送至蘇州泓迅生物科技股份有限公司進行測序鑒定,獲得Ectdh基因序列,并推測出其氨基酸序列。結果表明,EcTDH的序列及在載體上的插入位置與預期的結果相符,表明重組菌E.coliBL21(DE3)/pACYCDuet-1-Ectdh已成功構建。

圖2 Ectdh重組子的PCR擴增(A)及pACYCDuet-1-Ectdh的通用引物驗證(B)Fig.2 Agarose gel electropherograms of PCR amplified Ecctdh (A) and universal primer validation for pACYCDuet-1-Ectdh (B)
2.2 EcTDH的序列分析
將上述推測出的EcTDH氨基酸序列在ProtParam(https://web.expasy.org/protparam/)上進行理論理化性質的預測,結果表明,EcTDH的理論等電點為5.81,理論分子質量為37 173.94 Da,不穩定指數II為24.46,是一個較穩定的蛋白,其在E.coli和酵母體內的半衰期分別能達到10 h和20 h以上。利用TMHMM-2.0服務器預測蛋白跨膜結構的結果顯示,EcTDH的所有氨基酸殘基均為膜外殘基,表明EcTDH理論上相當容易實現胞外分泌型表達。將EcTDH的氨基酸序列和已經報道來源不同的TDH氨基酸序列進行分析(激烈熱球菌(Pyrococcus furiosus)、堀越熱球菌(Pyrococcus horikoshii)、柯達熱球菌(Thermococcus kodakaraensis)),用Jalview軟件進行多序列比對,結果如圖3所示,已報道的文獻表明來自P.horikoshii的L-TDH氨基酸殘基半胱氨酸Cys97、Cys100、Cys103、Cys111位在L-TDH中是保守的[30-31],其對應來自E.coli的L-TDH Cys93、Cys96、Cys99、Cys107。建立L-TDH的三維結構模型如圖4所示,L-TDH對于底物L-蘇氨酸的催化中心是在107位的半胱氨酸。

圖3 EcTDH與其他3 個代表性TDH的多序列比對Fig.3 Multisequence alignment of EcTDH with three other representative TDHs
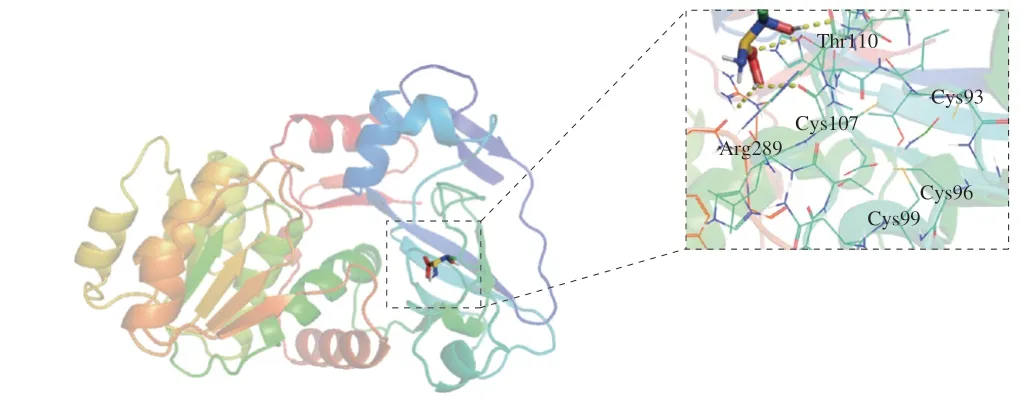
圖4 EcTDH三維結構圖Fig.4 Three-dimensional structure analysis of EcTDH
2.3 重組E. coli的誘導表達及鑒定
分別將E.coliBL21(DE3)/pACYCDuet-1、E.coliBL21(DE3)/pACYCDuet-1-Ectdh菌株按照1.3.2節方法進行誘導表達,將經誘導表達后的菌體進行離心,收集菌體重懸后進行超聲破碎,8 000 r/min、4 ℃離心15 min后收集裂解上清液。將收集裂解的上清液進行SDS-PAGE及酶活力分析。如圖5所示,E.coliBL21(DE3)/pACYCDuet-1-Ectdh裂解上清液經純化后在約37 kDa處有明顯的特異性條帶,與EcTDH的理論分子質量37 173.94 Da基本一致,表明已成功實現了EcTDH在E.coli的可溶性表達。

圖5 重組E. coli表達產物的SDS-PAGE分析Fig.5 SDS-PAGE analysis of recombinant Escherichia coli expression products
各重組子酶活力測定結果顯示,E.coliBL21(DE3)/pACYCDuet-1的酶活力為0.24 IU/mL,而E.coliBL21(DE3)/pACYCDuet-1-Ectdh的細胞裂解液中檢測到EcTDH活力可達到19.13 IU/mL,與本底細胞相比,其酶活力是E.coli本底細胞表達的79 倍,純化后的比活力可達12.77 IU/mg,表明EcTDH表現出較高的酶活力。比較已有文獻報道的不同來源的L-TDH活力結果如表3所示。可以看出,來源于E.coli的L-TDH經載體pACYCDuet-1和pET28a表達后比活力產生較大差別,主要是由于酶活力定義不同,以及酶活力測定時底物和輔酶終濃度的差異導致,它們會對表觀酶活造成較大的影響。文獻報道[32]測定酶活力體系中底物L-蘇氨酸和輔酶NAD+終濃度分別為100 mmol/L和200 mmol/L,而在本研究中終濃度分別為8 mmol/L和3 mmol/L。他們將每分鐘消耗1mol NAD+所需的酶量定義為1 U,而本研究將每分鐘產生1 μmol NADH所需的酶量定義為1 IU。

表3 不同來源L-TDH活力與比活力的對比Table 3 Specific activity of L-TDH from different sources
2.4 重組L-TDH的溫度特性
由圖6可知,在溫度為20~45 ℃范圍內,酶促反應速率隨溫度的升高而不斷提高,超過45 ℃后,反應速率逐漸下降,表明EcTDH的最適反應溫度為45 ℃,在40~45 ℃之間時具有較高酶活力。這與之前文獻報道的最適溫度一致[22]。
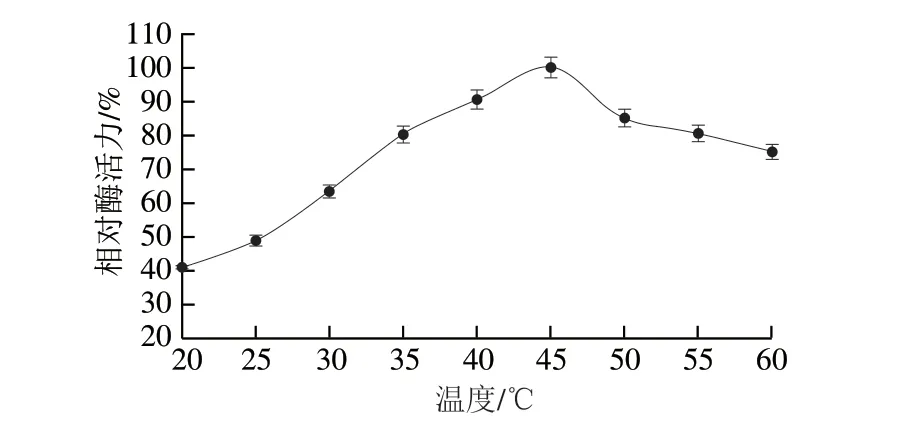
圖6 重組L-TDH的最適反應溫度Fig.6 Optimal reaction temperature for recombinant L-TDH
如圖7所示,在35 ℃和40 ℃保溫120 min后的殘留酶活力仍然高于90%,這表明它在35~40 ℃范圍內有較好的穩定性。與之前文獻報道的在35~40 ℃條件下測定的殘留酶活力相比,本研究殘留酶活提高了12.5%;在45、50、55 ℃殘留酶活力達到50%所對應的時間分別為78、40 min和20 min。

圖7 重組L-TDH溫度穩定性Fig.7 Temperature stability of recombinant L-TDH
2.5 重組L-TDH的pH值特性
緩沖液過酸過堿都會導致酶的空間結構的改變,從而使酶的活性降低,由圖8可知,重組酶的最適反應pH值在8.5~9.5之間,相對酶活力可達80%以上,EcTDH最適反應pH值為9.0。
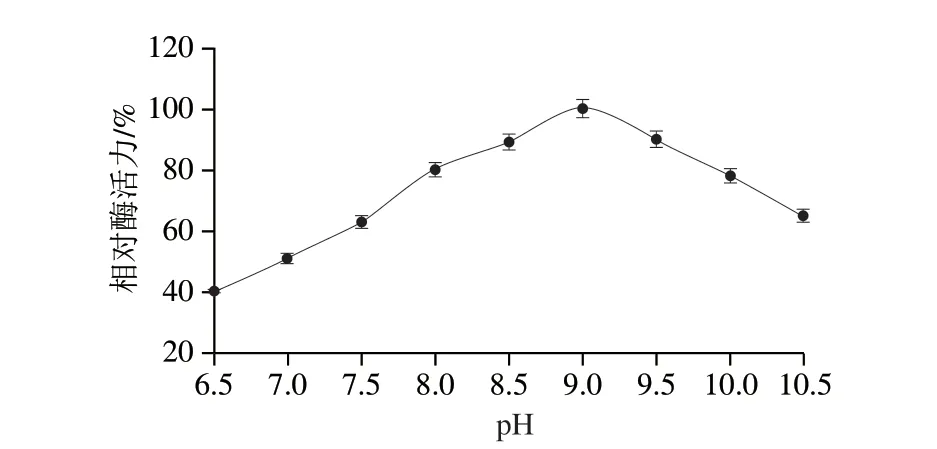
圖8 重組L-TDH的最適pH值Fig.8 Optimal reaction pH for recombinant L-TDH
如圖9所示,重組L-TDH在pH 7.5~8.5的緩沖液中保持1.5 h后殘留酶活力均能保持90%以上,表明它在pH 7.5~8.5范圍內有較好的穩定性,其pH值適應性相對較廣,在pH 8.5~10.0范圍內殘留酶活力在65%以上,而已報道的L-TDH在pH值為10時,其殘留酶活力僅達到60%,表明在pH值穩定性方面,該研究中L-TDH穩定性更好。
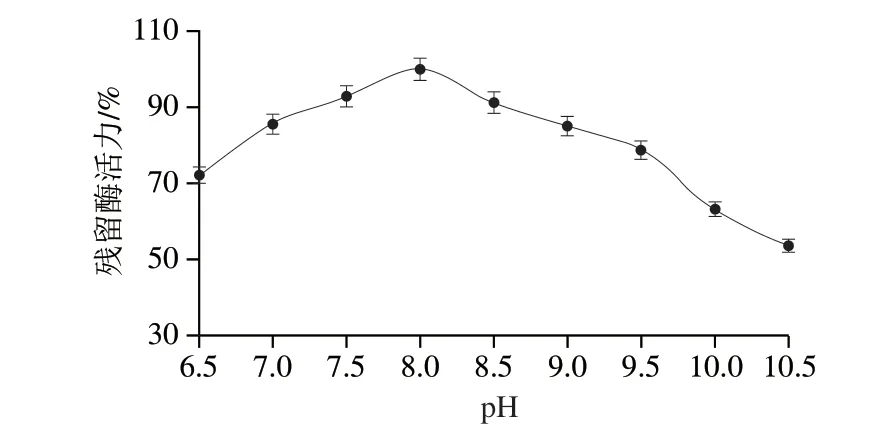
圖9 重組L-TDH的pH值穩定性Fig.9 pH stability of recombinant L-TDH
2.6 重組TDH的動力學參數
表4結果顯示,EcTDH對L-蘇氨酸的Kcat為1.13 s-1,Km為7.76 mmol/L,Vmax為1.827 μmol/(min·mg),Kcat/Km值為0.15 L/(mmol·s)。EcTDH對輔酶NAD+的Kcat為1.52 s-1,Km為1.61 mmol/L,Vmax為2.46 μmol/(min·mg),Kcat/Km值為0.94 L/(mmol·s)。對比文獻來源于Cupriavidus necator的野生型L-TDH和各位點突變型L-TDH的Km和Kcat值(表4),L-TDH因其空間結構的不同,進而影響反應過程中活性位點殘基的運動使得動力學參數產生差異。
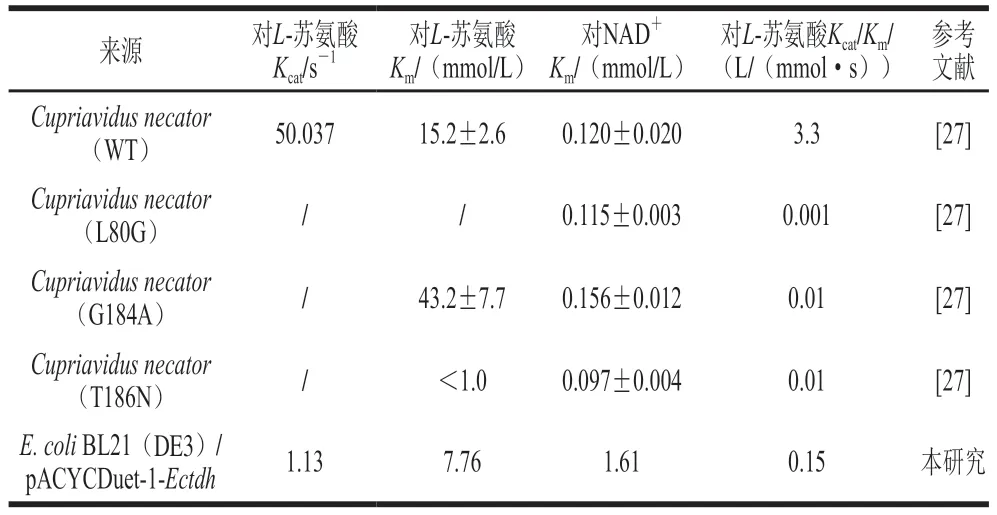
表4 不同L-TDH動力學參數的比較Table 4 Comparison of kinetic parameters of L-TDH from different sources
2.7 2,5-DMP及底物標準品的HPLC分析
按照1.3.9節方法,將分析純的2,5-DMP在Thermo Hypersil C18柱上進行HPLC分析,在該分析條件下,2,5-DMP的保留時間約為6.55 min,將2,5-DMP不同濃度對相應的峰面積進行線性擬合,獲得2,5-DMP的標準曲線,得出2,5-DMP濃度對應的回歸方程為y=2 062.4x+354.29(x為2,5-二甲基吡嗪濃度(mmol/L);y為峰面積,相關系數為0.997 8),結果表明在該HPLC分析條件及濃度范圍下,2,5-DMP濃度與峰面積呈良好線性相關。因此,利用此法可以對2,5-DMP進行精確定量分析。同樣條件下對底物L-蘇氨酸進行HPLC分析,底物的保留時間約為3.06 min。在相同的分析條件下,L-蘇氨酸的保留時間與2,5-DMP保留時間差約3.5 min,結果表明在該色譜條件下能夠實現L-蘇氨酸與2,5-DMP的良好分離,該色譜方法能夠用于催化產物的分析鑒定。
2.8 重組E. coli全細胞催化L-蘇氨酸合成2,5-DMP
按照1.3.7節方法對L-蘇氨酸進行全細胞催化的生物轉化。E.coliBL21(DE3)/pACYCDuet-1和E.coliBL21(DE3)/pACYCDuet-1-Ectdh的催化產物經過離心取樣后按照1.3.8節方法進行HPLC分析,保留時間約為6.55 min和3.06 min處有明顯的特征峰,它們與相同分析條件下2,5-DMP和L-蘇氨酸標品的保留時間一致,表明在該催化條件下,部分L-蘇氨酸已被轉化成2,5-DMP。在不同時間取樣分析、計算各時間節點的產物得率,它們的催化反應進程曲線如圖10所示。反應進行到36 h,反應基本達到平衡狀態,2,5-DMP得率基本不再隨時間的變化而變化,E.coliBL21(DE3)/pACYCDuet-1-Ectdh的產物得率約為12%,顯著高于E.coliBL21(DE3)/pACYCDuet-1的轉化水平。結果表明,強化L-TDH的表達可顯著提高2,5-DMP合成效率,但僅依靠強化TDH的表達難以實現L-蘇氨酸的高效轉化。

圖10 L-TDH對催化反應進程的影響Fig.10 Effect of L-TDH on the catalytic reaction process
3 討 論
L-TDH是以L-蘇氨酸為底物合成高值化合物2,5-DMP的關鍵酶,本研究對挖掘出來源于E.coli的L-TDH進行了高效表達及酶學性質分析。本研究從E.coli中克隆出L-TDH的基因片段,再借助pACYCDuet-1表達質粒在E.coliBL21(DE3)中實現了高水平的可溶性表達,酶活力測定結果表明,E.coliBL21(DE3)/pACYCDuet-1-Ectdh發酵液中L-TDH活力可達19.13 IU/mL,純化后重組L-TDH的比活力為12.77 IU/mg。與已報道文獻相比[19],本研究表達水平和比活力均較高,比活力的差異可能歸因于蛋白序列本身的差異,兩者的序列相似度為77%;而表達水平的差異可能主要歸因于宿主的差別,所用的E.coli表達系統比枯草芽孢桿菌表達系統具有更大表達潛力。也利用強化了TDH表達水平的重組E.coliBL21(DE3)/pACYCDuet-1-Ectdh進行了轉化L-蘇氨酸合成2,5-DMP的初步研究,它利用強化的TDH以及自身本底表達的NADH氧化酶、蘇氨酸轉運蛋白和氨基酸氧化酶等能夠協同催化2,5-DMP的合成,反應24 h后產物得率約12%,顯著高于沒有強化TDH表達的E.coliBL21(DE3)/pACYCDuet-1,這說明TDH確實在轉化L-蘇氨酸合成2,5-DMP的過程中起關鍵作用。然而,僅強化TDH表達難以實現L-蘇氨酸的高效轉化,后續研究需要參考前人的研究[32],充分利用代謝工程和酶工程的手段,繼續強化2,5-DMP積累正相關的基因表達,敲除2,5-DMP積累負相關的基因,以期進一步提高2,5-DMP的得率。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
