ATP5B在結(jié)直腸癌組織中的表達(dá)及意義
2023-11-25 12:55:50李時(shí)孟岳斯琦
中國實(shí)驗(yàn)診斷學(xué) 2023年11期
林 英,李時(shí)孟,岳斯琦,楊 琦*
(1.北京中醫(yī)藥大學(xué)東直門醫(yī)院 檢驗(yàn)科,北京100700;2.吉林大學(xué)中日聯(lián)誼醫(yī)院 檢驗(yàn)科,吉林 長春130033)
結(jié)直腸癌是全球第三大癌癥死因,每年有超過185萬病例被發(fā)現(xiàn),85萬人死亡[1]。結(jié)直腸癌初期起病隱匿,發(fā)展緩慢,多數(shù)病例是在癌癥晚期才被確診,有效的診斷及發(fā)病機(jī)制的研究對腫瘤的預(yù)后有重要意義[2]。本研究通過二維色譜質(zhì)譜聯(lián)用技術(shù)、免疫印跡實(shí)驗(yàn)、免疫組化實(shí)驗(yàn)來探究ATP5B蛋白在結(jié)直腸癌組織與癌旁組織的表達(dá)及意義,為結(jié)直腸癌的臨床診斷及發(fā)病機(jī)制的研究提供新的方向。
1 材料與方法
1.1 研究對象
選用2021年4月至2022年10月在吉林大學(xué)中日聯(lián)誼醫(yī)院初次就診且未經(jīng)處理的結(jié)直腸癌患者組織及其配對的癌旁正常組織,癌組織選取癌灶中心區(qū)域,癌旁組織來自于距離癌組織7 cm以上的正常黏膜。共計(jì)41例。完成采集后用冷的PBS進(jìn)行沖洗,液氮速凍,保存至-80°冰箱,本研究通過吉林大學(xué)中日聯(lián)誼醫(yī)院倫理委員會審批(20220126004)。
1.2 儀器與試劑
EASY-nLC 1200高效液相色譜儀購自美國Agilent公司,LTQXL離子阱質(zhì)譜儀購自美國Thermo Fisher Scientific公司,電泳儀,ATP5B單抗購自美國Affinity公司,兔抗人ATPB5多克隆抗體購自英國Abcum公司、HRP-羊抗兔 IgG購自美國CST公司,β-actin多抗、BCA蛋白質(zhì)定量試劑盒購自sab公司,DNA酶,RNA酶購自美國西格瑪公司。組化試劑盒DAB顯色劑購自中國Servicebio公司。無水乙醇、二甲苯購自致遠(yuǎn)化學(xué)試劑有限公司。
1.3 方法
1.3.1蛋白提取及樣品制備 將樣品從-80°冰箱中取出并倒入預(yù)冷的研缽中將其研磨為粉末狀,用刮勺將粉末轉(zhuǎn)移至加入適量蛋白裂解液的ep管中。充分混勻后冰上靜置30 min。在ep管中加入適量的DNA酶和RNA酶后放入4°離心機(jī)(12 000 rpm)離心10 min。根據(jù)BCA試劑盒中步驟測定蛋白樣品的濃度,將樣品板放入酶標(biāo)儀中,記錄各個(gè)樣品在OD562條件下的OD值并繪制蛋白質(zhì)濃度標(biāo)準(zhǔn)曲線,在ep管中加入1/4原體積的上樣緩沖液,95°條件下煮沸5 min,室溫靜置15 min后放入-80°冰箱中待用。
1.3.2二維質(zhì)譜色譜聯(lián)用技術(shù)鑒定蛋白多肽 0.1%的甲酸蛋白加入樣品使其充分溶解,取50 μg樣品進(jìn)行上樣,通過強(qiáng)陽離子柱和反向柱對樣品進(jìn)行洗脫。經(jīng)LTQXL離子阱質(zhì)譜儀分析檢測,獲取全掃描及二級掃描處理后的質(zhì)譜數(shù)據(jù)圖。
1.3.3免疫印跡試驗(yàn) SDS-PAGE電泳,將促凝劑冷卻至室溫后按照比例加入到分離膠中充分混勻,1.0 mm玻璃板中加入4.5 ml的混合溶液,迅速加入適量超純水后靜置45 min,倒棄超板內(nèi)超純水。用濾紙吸去板內(nèi)多余液體,加入配制好的濃縮膠后垂直插入電泳梳,靜置50 min。樣品經(jīng)10%分離膠、5%濃縮膠將蛋白按照不同的分子量大小進(jìn)行轉(zhuǎn)印。用快速封閉液浸潤PVDF膜10 min,倒入一抗低溫孵育過夜,PBS-T洗膜3次,每次10 min,加入二抗(1∶8 000)低溫孵育1 h。PBS-T洗膜5次,每次10 min,通過ECL發(fā)光法暗室顯影。
1.3.4免疫組化試驗(yàn) 將組織樣本烤片后脫蠟水化,將切片放入枸櫞酸鹽抗原修復(fù)緩沖液中煮沸3 min,待其冷卻至室溫后拿出切片用PBS溶液清洗3次,用濾紙吸去殘留液體后放入濕盒內(nèi)。確定組織邊緣后畫圈,在圈內(nèi)滴加3%BSA室溫封閉30 min,加入ATP5B一抗稀釋液(1∶500)后低溫孵育過夜。PBS洗片3次,每次5 min,滴加二抗室溫孵育50 min,PBS洗片5次,每次3 min,甩干后滴加DAB顯色劑并控制反應(yīng)時(shí)間,超純水沖洗切片后用蘇木素復(fù)染2 min,用鹽酸乙醇洗滌切片5次后放入PBS緩沖溶液中。脫水封片后由兩名病理科醫(yī)生在顯微鏡下進(jìn)行圖像采集與分析。
1.4 統(tǒng)計(jì)學(xué)方法
2 結(jié)果
2.1 二維色譜與質(zhì)譜聯(lián)用技術(shù)分析結(jié)果
本實(shí)驗(yàn)癌組織及癌旁組織中以圖譜數(shù)為依據(jù)判斷同一蛋白的豐度,當(dāng)兩個(gè)樣品中蛋白圖譜數(shù)比值 ≥1且蛋白圖譜數(shù)差值 ≥72 時(shí),在篩選時(shí)將其判定為差異蛋白。在此標(biāo)準(zhǔn)下檢測到NUP107、CUL48、FCG、E5RH53、ENO1、TOMM40、RPS14、FGB共8個(gè)上調(diào)蛋白,CNN1、DESM、VCL、A1AT、SYNM、KCRB、MYH14、ATP5B等8個(gè)下調(diào)蛋白。ATP5B作為其中1個(gè)下調(diào)蛋白,質(zhì)譜圖結(jié)果見圖1。

圖1 ATP5B 質(zhì)譜圖結(jié)果
2.2 免疫印跡試驗(yàn)結(jié)果
運(yùn)用Western blot方法對14對組織的ATP5B表達(dá)量進(jìn)行分析,ATP5B在結(jié)直腸癌組織中表達(dá)量低于癌組織,如圖2所示。
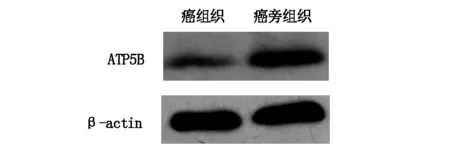
圖2 ATP5B 免疫印跡結(jié)果
2.3 免疫組化試驗(yàn)結(jié)果
結(jié)果顯示,ATP5B蛋白在癌組織中強(qiáng)陽性表達(dá)占17%(7/41),中等陽性表達(dá)占22%(9/41),弱陽性表達(dá)占61%(25/41)。在癌旁組織中,強(qiáng)陽性表達(dá)占22%(9/41),中等陽性表達(dá)占41%(17/41),弱陽性表達(dá)占37%(15/41)。在癌組織和癌旁組織中ATP5B的陽性率差異具有統(tǒng)計(jì)學(xué)意義(P<0.05),見圖3。
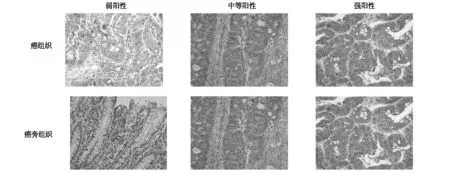
圖3 ATP5B 免疫組化結(jié)果
3 討論
癌癥的發(fā)展通常伴有能量代謝方式的變化,為癌細(xì)胞提供代謝能量并合成代謝前體產(chǎn)物是結(jié)直腸癌發(fā)生發(fā)展的主要表現(xiàn)之一[3]。線粒體是細(xì)胞能量代謝的核心細(xì)胞器,ATP合酶是調(diào)節(jié)其生物學(xué)功能的關(guān)鍵酶復(fù)合物之一[4]。ATP合成酶由兩個(gè)相連的多亞基復(fù)合物組成,其催化部分由5個(gè)不同的亞基組成:3個(gè)α亞基,3個(gè)β亞基,γ、δ 和ε亞基各1個(gè),其功能主要是在氧化磷酸化的過程中以α亞基和β亞基高度協(xié)調(diào)的構(gòu)象變化為表現(xiàn)催化ATP合成或水解[5]。ATP5B基因主要編碼與線粒體能量代謝核心相關(guān)的F1β亞基[6]。近年來,陸續(xù)有研究發(fā)現(xiàn)ATP5B與乳腺癌[7]、膠質(zhì)母細(xì)胞瘤[8]、急性粒細(xì)胞白血病[9]、腎透明細(xì)胞癌[10]等惡性腫瘤的發(fā)生發(fā)展密切相關(guān),MOTTAGHI-DASTJERDI等[11]通過生信分析推測ATP5B為胃癌發(fā)病機(jī)制的中樞基因,有研究[12]證實(shí)ATP5B在胃癌腫瘤組織中的高表達(dá)與患者的腫瘤大小、TNM分期,淋巴結(jié)轉(zhuǎn)移和預(yù)后不良呈正相關(guān),促進(jìn)了腫瘤細(xì)胞的遷移、侵襲和增殖,過表達(dá)ATP5B激活FAK/AKT/MMP2通路促進(jìn)了胃癌的生長,因此認(rèn)為其可能對胃癌診斷有一定的輔助作用。
本研究選擇通過液質(zhì)聯(lián)用技術(shù)(LC-MS)分析41例患者的結(jié)直腸癌及癌旁組織差異表達(dá)蛋白,得到了結(jié)直腸癌相關(guān)的差異表達(dá)蛋白質(zhì)圖譜,發(fā)現(xiàn)ATP5B是下調(diào)蛋白之一。利用免疫印跡試驗(yàn)和免疫組化試驗(yàn)對LC-MS的結(jié)果進(jìn)行驗(yàn)證,發(fā)現(xiàn)在結(jié)直腸癌旁組織中ATPB1的表達(dá)量顯著高于癌組織中的表達(dá)量,因此,ATP5B低表達(dá)可能與結(jié)直腸癌的發(fā)生發(fā)展存在聯(lián)系,為結(jié)直腸癌的初篩和研究提供了新的靶點(diǎn)。ATP5B更深入的生信分析及腫瘤進(jìn)展機(jī)制的研究尚不完善,需要后續(xù)實(shí)驗(yàn)進(jìn)行更全面的探討。
 中國實(shí)驗(yàn)診斷學(xué)2023年11期
中國實(shí)驗(yàn)診斷學(xué)2023年11期
- 中國實(shí)驗(yàn)診斷學(xué)的其它文章
- 線上教學(xué)中教學(xué)交互對醫(yī)學(xué)生在線學(xué)習(xí)滿意度的影響研究
- lncRNA MALAT1調(diào)節(jié)miR-106b-5p/TLR4軸對慢性阻塞性肺疾病大鼠肺組織炎性損傷的影響
- miR-196通過靶向調(diào)控CDK6表達(dá)對骨肉瘤細(xì)胞增殖的影響
- H4及A172神經(jīng)膠質(zhì)瘤細(xì)胞系高親和腺相關(guān)病毒載體的研究
- 長鏈非編碼RNA UCA1促進(jìn)人骨髓間充質(zhì)干細(xì)胞成骨分化研究
- 孔源性視網(wǎng)膜脫離硅油填充術(shù)后黃斑區(qū)微血管變化