赤霉素及二氫茉莉酸丙酯對抑制溫州蜜柑果實浮皮的影響
2024-01-20 01:52:32余文莉劉德春匡柳青
核農學報 2024年2期
余文莉 余 旋,2 劉德春 楊 莉 匡柳青 宋 杰 劉 勇,* 胡 威,*
(1江西農業大學農學院,江西 南昌 330045;2江西省農業技術推廣中心,江西 南昌 330046)
2021年,我國柑橘產量為5 595.6萬t[1],產業規模位居全球第一,寬皮柑橘產量也居世界首位[2]。浮皮是寬皮類柑橘果實成熟過程中普遍發生的生理性病害[3-4],嚴重影響果實品質。近年,各地采用設施延遲采收栽培技術生產完熟寬皮柑橘,經濟效益提高的同時也使浮皮問題更加突出,因此亟待研究有效控制浮皮的方法。
采前控制柑橘果實浮皮的措施主要包括葉面噴施鈣和植物生長調節劑,鈣可顯著降低溫州蜜柑果實浮皮指數[5-6]。張秋明等[7]研究表明,醋酸鈣、赤霉素(gibberellic acid,GA3)均能有效降低溫州蜜柑果實浮皮率,以GA3效果最佳,并且果皮膜脂過氧化程度降低。Garcia-Luis 等[8]及Pozo 等[9]也證實GA3可顯著降低溫州蜜柑果實浮皮發生率。但單獨使用鈣抑制果實浮皮的效果并不理想[10-11],而GA3會使果實轉色困難[8-9]。研究表明,外源蕓苔素內酯(brassinolide,BR)能促進鈣吸收與轉運并增加其在細胞質中的積累[12-13],因此,BR與鈣組合使用對抑制浮皮的效果值得進一步研究。此外,二氫茉莉酸丙酯(prohydrojasmon,PDJ)能顯著改善梨和葡萄等果實的著色[14-15],因此,將GA3與PDJ 聯合噴施可有效降低溫州蜜柑果實浮皮,同時減輕對果實著色的影響[16],但著色難問題依然難以避免,仍需結合其他植物激素進行調節[17]。綜上,鈣劑、GA3與PDJ聯合使用的方法仍需進一步優化。
研究認為,浮皮的發生機制與果皮衰老相關,涉及到果皮細胞形態和結構的變化,即細胞壁物質果膠、纖維素及半纖維素等的降解導致細胞間連接松散[3,18]。浮皮果中參與果皮代謝的相關酶類活性發生改變,如果膠甲酯酶(pectin methyl enzyme,PME)、纖維素酶(cellulase,Cx)等的活性顯著高于正常果[19-20];此外,浮皮果中抗氧化相關酶類活性顯著下降;乙烯會加劇浮皮的產生,而抑制乙烯的合成則能降低浮皮的發生率[21-22],同時生長素(indoleacetic acid,IAA)、細胞分裂素(cytokinin,CTK)以及GA3也影響著浮皮的發生;浮皮果中,參與以上代謝相關的基因表達量也與正常果實有顯著差異[23]。然而,GA3和PDJ 處理是否影響果皮細胞壁物質的降解從而抑制果實浮皮鮮見報道。
因此,本研究以設施完熟栽培溫州蜜柑宮川(CitrusunshiuMarc.)為試驗材料,通過噴施不同濃度和不同次數的鈣劑、GA3和PDJ,分析其在減輕浮皮現象中產生的作用,從而研究適宜的寬皮柑橘果實浮皮調控措施,同時探究 GA3和PDJ 抑制浮皮發生的生理及分子機制,以期為進一步提高溫州蜜柑品質提供實踐和理論依據。
1 材料與方法
1.1 植物材料與處理
以設施完熟栽培的溫州蜜柑宮川為試驗材料,選取生長健壯、長勢均衡的結果樹為試驗樹。各處理設置見表1,并設置清水對照。選取果實生長狀況相同且處于同一受光方向的樹枝做好標記,每枝約40~50 個果實,每組設置3 個重復,用于記錄成熟期果實浮皮指數的動態變化。在果實開始出現浮皮現象后,即花后天數(Days after flower,DAF)204(DAF 204)每隔15 d統計浮皮指數,并選擇大小、生長狀況一致且無機械損傷的果實用于后續試驗。
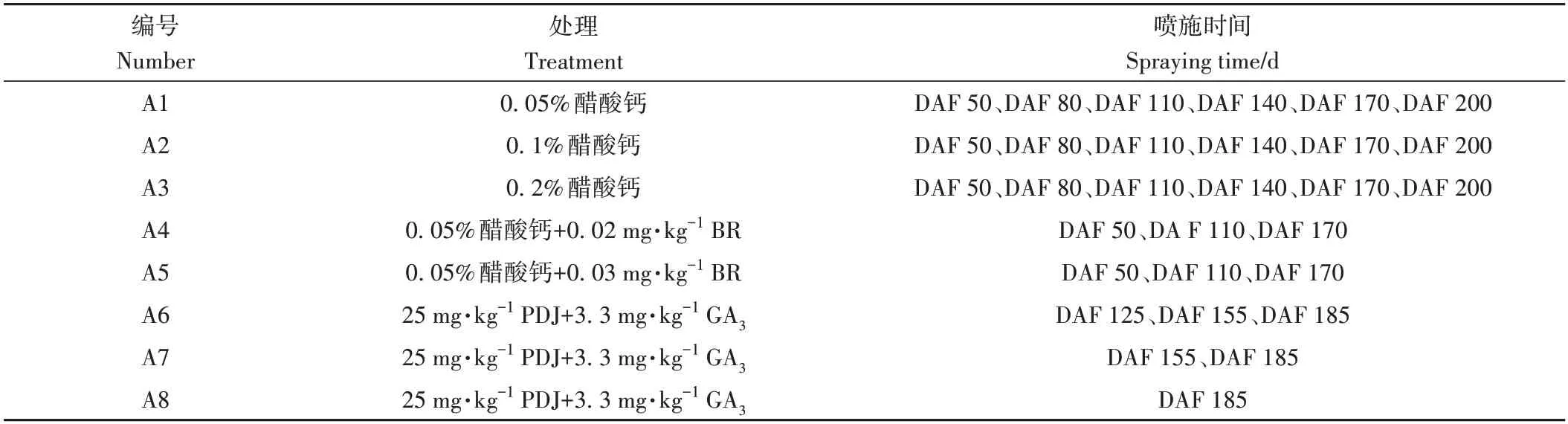
表1 試驗設計Table 1 Experimental design
1.2 試驗方法
1.2.1 浮皮指數的測定 浮皮程度分為4級,浮皮無(0)、浮皮輕(1為僅果柄端浮皮)、浮皮中(2為果柄端與兩側都浮皮)、浮皮重(3為全果浮皮),通過指頭觸感判斷每個果實的浮皮程度,最后計算果實的浮皮指數:
浮皮指數=[Σ(浮皮級果數×代表級值)/(調查總果數×最高代表值)]×100。
1.2.2 果實細胞結構觀察 果實采摘后分離果皮,參考邱麗等[24]的方法制作果皮石蠟切片,最后用IX73型倒置顯微鏡(日本OLYMPUS 公司)觀察切片后的果皮細胞結構。
1.2.3 果實品質的測定 將收集的果實樣品用電子天平測定單果重,游標卡尺測定果實橫縱經和果皮厚度。用TA.XT ExpressC 型質構儀(英國Stable Micro Systems 公司)測定果皮穿刺強度,取最大值為果皮硬度。用MINOLTA CR-400 型色彩色差計(日本柯尼卡美能達公司)測定果皮色差,記錄L*、a*、b*值,并計算色差指數(color contribution index,CCI)=1 000a*/(L*×b*)。用PAL-1 型手持式糖度計(日本Atago 公司)測定可溶性固形物含量;用酸堿滴定法測定可滴定酸含量;用2,6-二氯靛酚氧化滴定法測定維生素C(vitamin C,Vc)含量;用蒽酮比色法測定總糖含量。
1.2.4 果膠含量的測定 參考馬麗等[25]的方法進行測定。
1.2.5 纖維素含量的測定(1)將干燥的果皮樣品充分研細,稱取0.1 g 樣品放入10 mL 刻度離心管中。用3%的十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)加滿,沸水浴1 h 讓其充分水解。于10 000 r·min-1離心10 min,殘渣用蒸餾水和丙酮(分析純)洗滌3 次,離心分離后去除上清液。(2)原離心管中用2 mol·L-1HCl加滿,沸水浴50 min,冷卻后離心10 min,收集上清液定容至100 mL,得到半纖維素溶液,用地衣酚法測定其含量。(3)殘渣用丙酮洗兩次,在60 ℃烘箱中烘干過夜。加入3 mL 的72% H2SO4于35 ℃水解1 h,加熱完畢后將離心管用蒸餾水加滿,100 ℃加熱1 h。冷卻后離心10 min,取上清液定容至100 mL,得到纖維素溶液,用蒽酮比色法測定其含量。
1.2.6 PME、多聚半乳糖醛酸酶(polygalacturonase,PG)、Cx 活性的測定 采用植物多聚半乳糖醛酸酶(PG)ELISA 檢測試劑盒、植物果膠甲酯酶(PME)ELISA 檢測試劑盒和植物纖維素酶(Cellulase)ELISA檢測試劑盒(上海優選生物科技有限公司)測定果皮樣品中的PME、PG、Cx 活性的變化,每組設置3 個生物學重復,所有步驟嚴格按照試劑盒說明書進行操作。
1.2.7 內源激素含量的測定 內源激素含量委托武漢邁特維爾生物科技有限公司測定。從處理樹上隨機選取果實采摘后擦凈分離果皮,標記整理并置于-80 ℃液氮速凍待用。
1.3 基因表達分析
使用植物總RNA 快速抽提試劑盒(上海生工生物工程股份有限公司)提取果皮總RNA,用反轉錄試劑盒(上海生工生物工程股份有限公司)反轉錄成cDNA。
實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)所用引物使用Primer 5.0 軟件設計,內參基因Actin及各個基因的引物序列見表2。使用Bio-Rad CFX96-PCR 實時熒光定量PCR 儀(美國Bio-Rad公司)進行基因表達量分析,采用20 μL 反應體系:TB Green?Premix Ex Taq 10 μL,10 μmol·L-1正反向引物各0.5 μL,cDNA 模板1 μL,RNase-free H2O 8 μL。反應程序為:95 ℃預變性10 min;95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸40 s,40 個循環。3 次生物學重復,采用2-ΔΔCt法計算基因相對表達量。
1.4 數據分析
利用Excel 2019 軟件進行數據整理分析以及DPS 9.01 軟件進行方差分析(P<0.05),用Origin 2019b 軟件作圖。
2 結果與分析
2.1 不同處理對果實浮皮的影響
本研究比較了采前使用不同濃度醋酸鈣、醋酸鈣加BR,以及GA3加PDJ 噴施不同次數對完熟栽培溫州蜜柑果實浮皮指數的影響。由表3 可知,除A8 處理外,其他處理均可抑制果實浮皮指數的增加,單獨噴施0.05%和0.1%質量濃度醋酸鈣(A1、A2處理)時,其浮皮指數直至花后264 d(DAF 264)仍顯著低于CK,而噴施0.2%醋酸鈣(A3 處理)時,其浮皮指數則在DAF 234 后與CK 無顯著差異,表明醋酸鈣濃度的增加并不能進一步降低果實浮皮指數。同時,0.05%醋酸鈣加BR 聯合噴施時(A4、A5 處理),其浮皮指數在DAF 234后與CK 無顯著差異,并且BR 濃度的增加會導致浮皮指數的上升。而GA3結合PDJ 采前噴施3 次(A6 處理)的處理果實浮皮指數直至完熟栽培溫州蜜柑全部采摘上市時(DAF 279)均顯著低于CK,而噴施2 次(A7 處理)處理的浮皮指數在DAF 219、DAF 234、DAF 264、DAF 279 同樣顯著低于CK,說明采前噴施2 次或3 次3.3 mg·L-1GA3以及25 mg·L-1PDJ 均能有效降低完熟栽培溫州蜜柑果實浮皮指數。
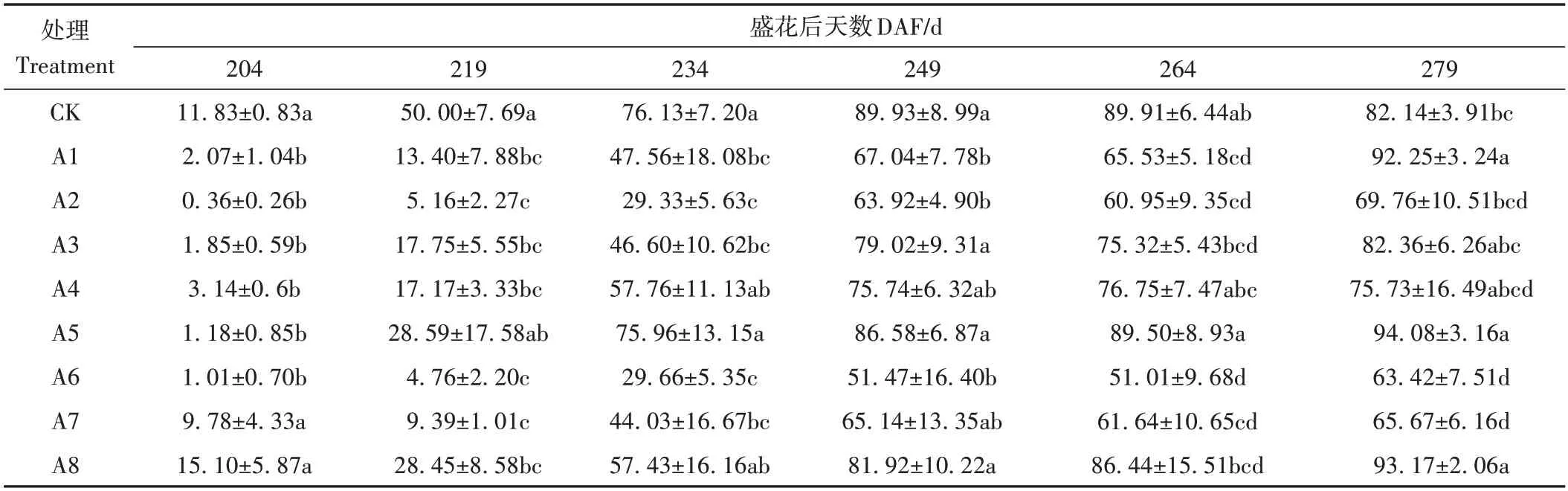
表3 不同處理在不同時期的果實浮皮指數Table 3 Fruit puffing index of different treatments in different stage
2.2 不同處理對果實品質的影響
各處理于完熟栽培果實品質最佳時期[26](DAF 264)進行取樣分析其內外在品質。由表4 可知,CK 單果質量為123.30 g,各處理單果質量均高于CK,但均無顯著差異;同時,各處理果實橫縱徑也與CK 均無顯著差異;除A8 處理外,其他各處理果皮厚度低于CK,但并無顯著差異。此外,A2、A3、A6處理CCI值均顯著低于CK,其他處理CCI值則與CK無顯著差異。
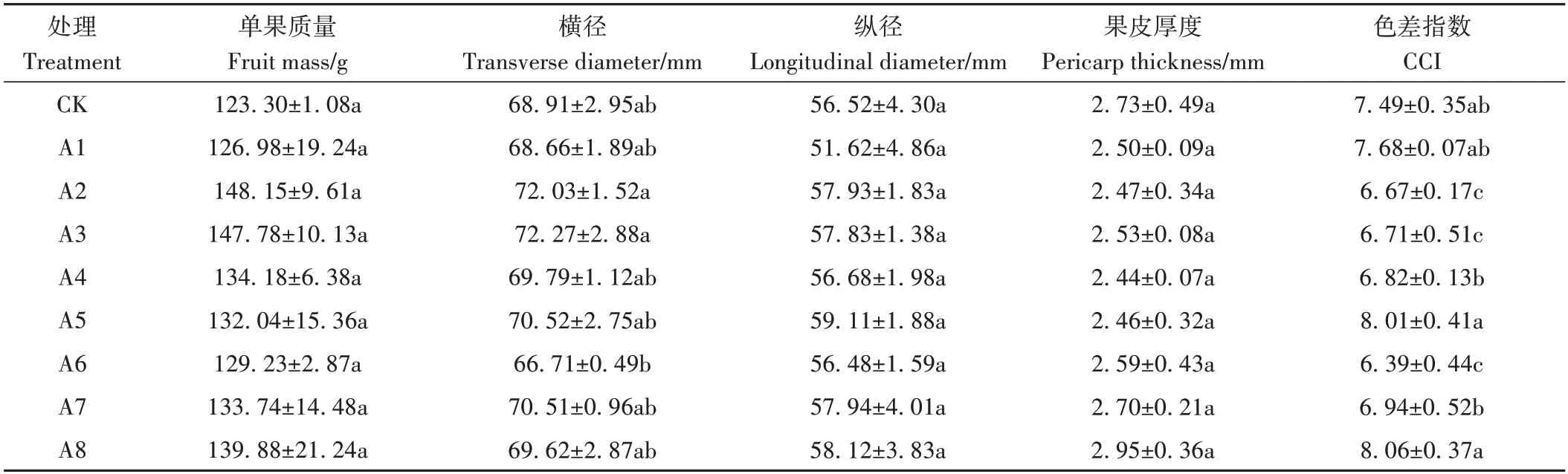
表4 不同處理對果實外觀品質的影響Table 4 Effect of different treatments on fruit appearance quality
由表5 可知,各處理果實可溶性固形物及可溶性糖含量與CK 相比均無顯著差異,而A4、A7 處理則較CK顯著增加了果實可滴定酸含量,其他處理果實可滴定酸含量雖與CK 無顯著差異,但均在不同程度上增加了果實可滴定酸含量。另外,A7處理顯著增加了果實Vc含量,其他處理則與CK差異不顯著。
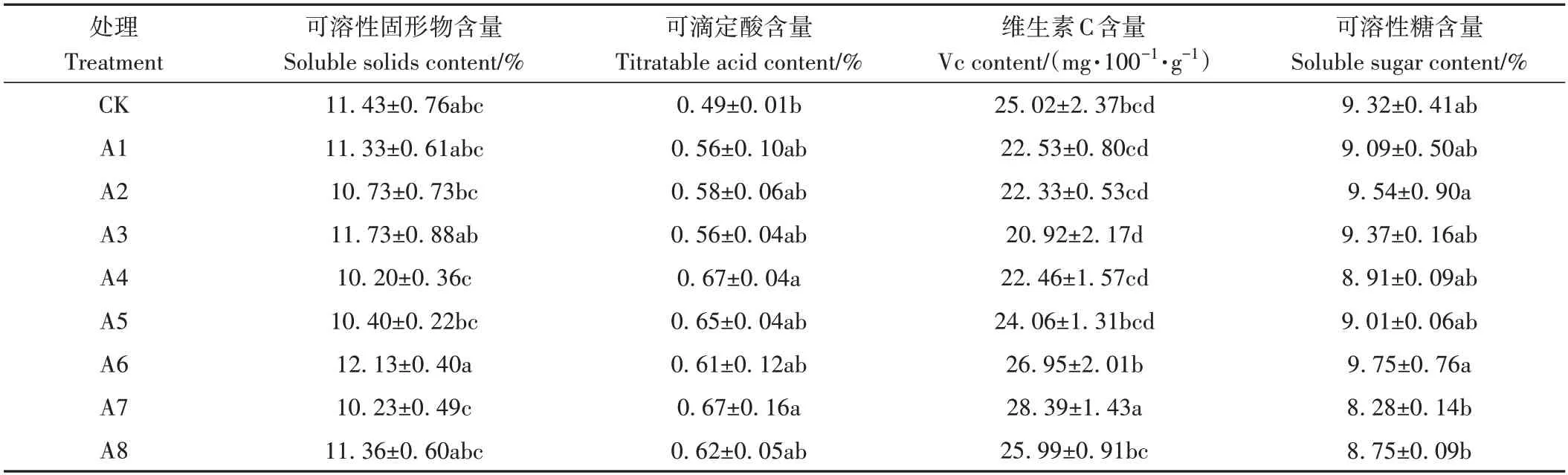
表5 不同處理對果實內在品質的影響Table 5 Effect of different treatments on the intrinsic quality of the fruit
2.3 GA3和PDJ處理后果皮解剖結構及形態觀察
以浮皮指數最低的A6處理果實為研究對象,觀察GA3和PDJ 處理后果皮解剖結構及細胞形態與CK浮皮果果皮的差異。由圖1 可知,果實縱切面可明顯觀察到CK 果皮與果肉出現較大間隙,果肉和果皮已經分離,而GA3和PDJ 處理后的果皮和果肉間隙小,未出現明顯的分離。對果皮進行石蠟切片觀察,浮皮果(圖2-A)的果皮細胞間隙大,細胞排列分散且無規律分布,大小不一,形狀呈不規則圓形或長橢圓形,油胞直徑較大、形狀不一。而GA3和PDJ處理后(圖2-B)的果皮組織細胞間隙小,大小相近,形狀較為規則呈圓形或近橢圓形,排列較為整齊致密,油胞直徑較小,邊緣整齊分布薄壁組織細胞。
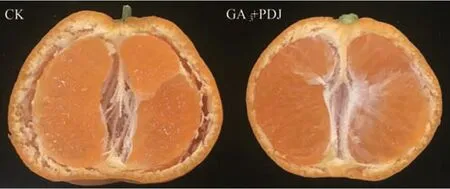
圖1 GA3和PDJ處理后果實浮皮狀態Fig.1 Puffing state of the fruit after GA3 and PDJ-treat
2.4 GA3和PDJ處理對果皮硬度及鈣含量的影響
由圖3-A 可知,在花后204~279 d,隨著時間的延長,果皮硬度整體呈下降趨勢,并且6個時期中除DAF 264時 PDJ和GA3處理后的果皮硬度顯著增加外,其余時期均極顯著高于CK,其硬度分別增加了42.0%、46.0%、48.3%、49.8%、42.9%和59.0%,除花后264 d,增加的幅度隨時間延長而逐漸增大。同時,分別檢測CK 果實出現明顯浮皮前時期(FPQ,DAF 204)、出現明顯浮皮后時期(FPH,DAF 264)以及GA3和PDJ 處理后與CK 同時期果皮中鈣元素含量,由圖3-B 可知,在FPQ 時期的CK 和GA3+PDJ 處理鈣元素含量分別為42.86、31.60 mg·L-1,CK 高于GA3+PDJ 處理。而在FPH 時期鈣元素含量分別為35.91、36.07 mg·L-1,CK與GA3+PDJ處理之間并無顯著差異。隨著取樣時期的延長,CK 鈣元素含量呈下降趨勢,而GA3+PDJ 處理鈣元素含量則較穩定。
2.5 GA3和PDJ處理對果皮果膠、纖維素含量的影響
本研究分析了GA3+PDJ 處理和CK 果皮中細胞壁物質含量的差異,由圖4-A 可知,GA3和PDJ 處理在FPQ 時期的可溶性果膠含量與CK 無顯著差異,而CK在FPH時期可溶性果膠含量為22.48 mg·g-1,GA3和PDJ處理為15.36 mg·g-1,極顯著低于CK。GA3和PDJ 處理在FPQ時期的原果膠含量顯著高于CK,在FPH時期極顯著高于CK(圖4-B)。兩個時期半纖維素(圖4-C)含量均極顯著高于CK,分別比CK 高38.83%和28.79%。兩個時期原果膠含量分別比CK高20.16%和41.06%。GA3和PDJ 處理在FPQ 時期纖維素(圖4-D)含量極顯著低于CK,在FPH時期則與CK無顯著差異,但在浮皮出現前后CK 的纖維素含量下降的幅度高于GA3和PDJ 處理,CK 纖維素含量下降45%,而GA3和PDJ 處理僅下降31.7%。這表明GA3和PDJ 可能是通過抑制果皮原果膠和半纖維素的降解來阻止細胞壁的崩解從而抑制浮皮的發生。
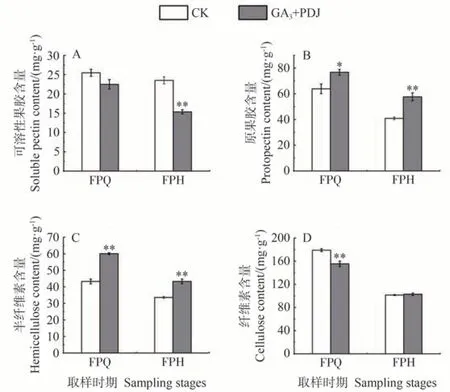
圖4 GA3和PDJ處理對果皮細胞壁物質的影響Fig.4 Effect of GA3 and PDJ-treat on peel cell wall composition
2.6 GA3 和PDJ 處理對果皮細胞壁水解酶活性的影響
本研究檢測了參與果皮細胞壁降解相關水解酶類活性,結果顯示在FPQ 時期,GA3和PDJ 處理與CK 的PME 活性分別為4.39 和3.26 U·g-1,在FPH 時期分別為4.67 和6.02 U·g-1,GA3和PDJ 處理在FPQ 時期顯著高于CK,而在FPH 極顯著低于CK,其PME 活性保持相對穩定,而CK 果皮中PME 活性在FPH 則急劇上升(圖5-A)。在FPH 時期的PG 活性較FPQ 時期有上升趨勢,但GA3和PDJ 處理和CK 之間并無顯著差異(圖5-B)。GA3和PDJ 處理果皮中Cx 活性在FPH 時期極顯著低于CK,在浮皮出現前后時期雖有上升趨勢但相比CK 上升幅度不大,而CK 的Cx 活性則上升2.3 倍以上(圖5-C)。可見,GA3和PDJ 對PME 和Cx 活性抑制作用較強,而對PG活性作用不明顯。
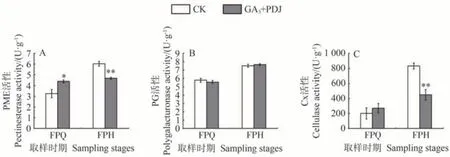
圖5 GA3和PDJ處理對果皮細胞壁水解酶活性的影響Fig.5 Effect of GA3 and PDJ-treat on the cell wall hydrolase activity of the pericarp
2.7 GA3和PDJ處理對果皮中激素含量的影響
由圖6-A 可知,在FPQ 和FPH 兩個時期,GA3和PDJ 處理后的果皮IAA 含量均極顯著高于CK,兩者均隨時間的延長呈下降趨勢。GA3和PDJ 處理果皮GA3含量在FPQ 時期極顯著高于CK,而在FPH 時期則極顯著低于CK(圖6-B)。GA3+PDJ 處理脫落酸(abscisic acid,ABA)含量在FPQ 時期與CK 無顯著差異,而在FPH 時期則極顯著高于CK,GA3+PDJ 處理和CK 的ABA含量分別為180.48和152.73 ng·g-1(圖6-C)。果皮中玉米素核苷(trans-Zeatin-riboside,ZR)含量則與GA3表現出相同趨勢,在FPQ 時期表現為GA3+PDJ 處理極顯著高于CK,而到FPH 時期反而極顯著低于CK(圖6-D)。乙烯合成前體1-氨基環丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)含量在FPQ 和FPH 兩個時期均表現為GA3+PDJ 處理極顯著低于CK,其含量分別只有CK 的38%和32%左右(圖6-E)。由此表明,GA3加PDJ 處理可顯著抑制果皮中IAA的減少和乙烯的合成。
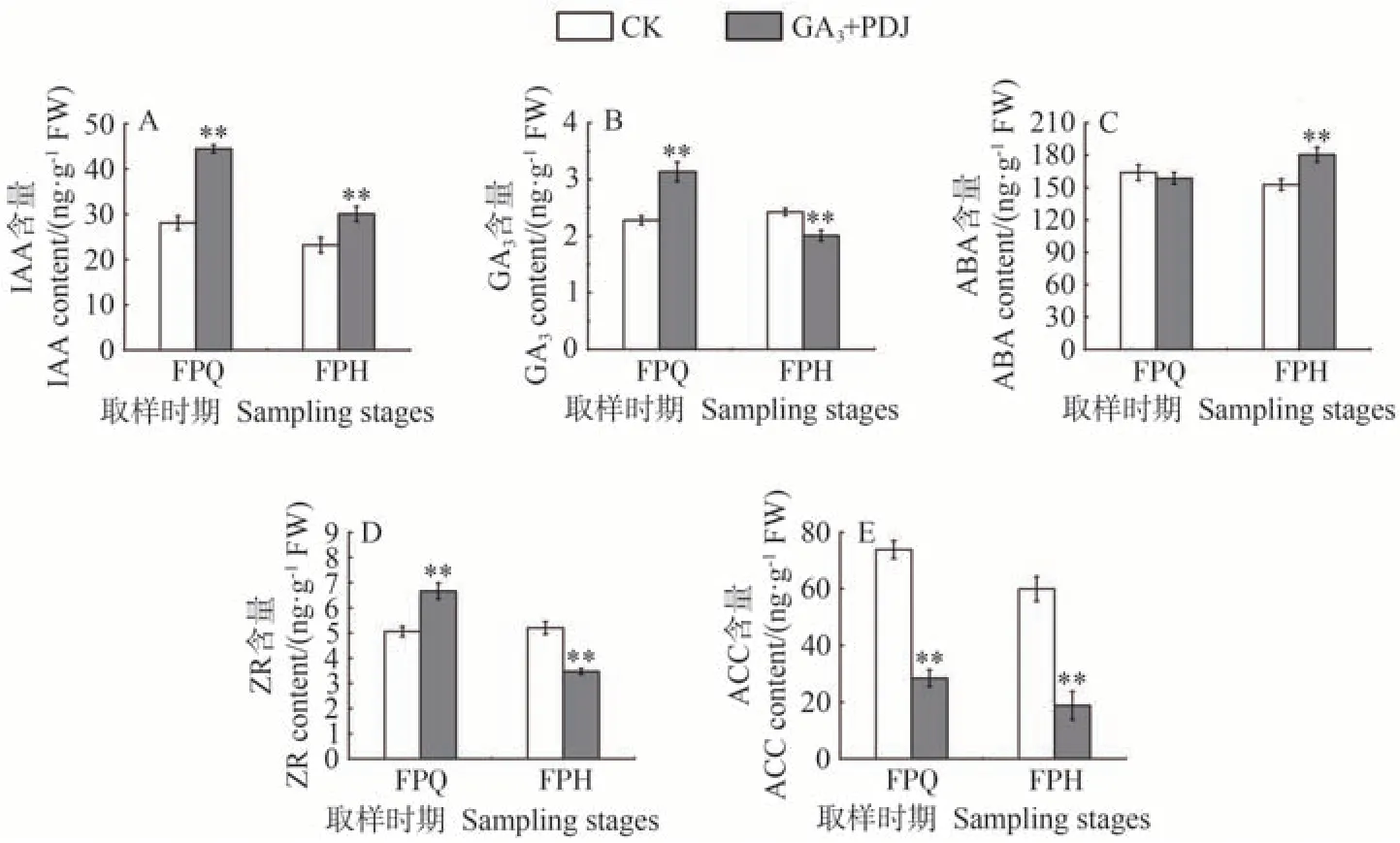
圖6 GA3和PDJ處理對果皮激素的影響Fig.6 Effect of GA3 and PDJ-treat on pericarp hormones
2.8 GA3和PDJ處理后浮皮相關基因表達分析
GA3和PDJ 處理后果皮中生長素原初響應基因CitIAA27表達量在FPQ 時期極顯著高于CK,在FPH時期顯著高于CK,分別比CK 高77.04%和30.52%(圖7-A)。乙烯合成關鍵酶基因CitACO表達量則在FPQ時期顯著低于CK,較CK低21.74%;在FPH時期極顯著低于CK,較CK 低88.07%(圖7-B)。乙烯信號響應蛋白基因CitETR1表達量在FPQ 和FPH 時期均極顯著高于CK,分別比CK高49.79%和343.92%(圖7-C)。纖維素酶基因CitCx與CitACO表達結果類似,在FPQ 時期顯著低于CK,在FPH 時期極顯著低于CK(圖7-D)。多聚半乳糖醛酸酶基因CitPG的相對表達量雖在兩個時期高于CK 但并無顯著差異(圖7-E)。果膠酯酶基因CitPME表達量在FPQ 和FPH 時期均極顯著低于CK(圖7-F)。經GA3和PDJ處理后,果皮中過氧化物酶基因CitPOD12表達量也均較CK 極顯著降低(圖7-G),表明果皮中膜脂過氧化水平降低。
3 討論
Sato 等[16]報道了GA3和PDJ 處理能顯著抑制露天栽培溫州蜜柑果實浮皮。同時,Ma 等[17]研究表明,GA3和PDJ處理雖能抑制果實浮皮但果實著色困難,需結合ABA 和萘乙酸(naphthalene-acetic acid,NAA)處理才能使果實正常著色。本研究中,在采前噴施2 或3 次GA3和PDJ 均能顯著抑制完熟栽培溫州蜜柑果實浮皮,延長果實掛果時間且對果實內在品質無顯著影響,而噴施2 次同樣對果實CCI 無顯著影響,因此本研究在前人的基礎上優化了GA3與PDJ 的使用方法。在南豐蜜橘幼果期噴施鈣能顯著抑制浮皮[27]。本試驗中不同鈣處理均能有效抑制完熟栽培溫州蜜柑果實浮皮的發生,但綜合而言噴鈣抑制浮皮效果并沒有GA3和PDJ處理好。此外,有研究顯示BR 能促進植物對鈣的吸收與運輸[12-13]。然而,本試驗中鈣結合BR 共同噴施溫州蜜柑,浮皮指數反而相對更高,Morillon 等[28]的研究認為BR 能增強細胞壁松弛度,同時誘導一些細胞壁修飾酶的表達,使細胞壁發生降解,由此推測BR 的這一特性雖能促進鈣的吸收但反而使果實浮皮更嚴重。同時,也說明鈣并不是決定果實浮皮的唯一因素,GA3加PDJ處理后鈣元素在浮皮果和正常果果皮中含量并無顯著差異也證實了上述推測。綜上所述,采前噴施2次GA3加PDJ抑制柑橘果實浮皮效果最佳。
Shiraishi 等[18]通過透射電鏡觀察果皮細胞壁超微結構,結果顯示浮皮果細胞間隙變大,細胞壁出現部分溶解,纖維含量減少,細胞間連接松散。本研究中GA3與PDJ 處理后的果皮石蠟切片顯示,細胞結構相比于CK 排列更加規則緊密,果皮油胞體積小,說明該處理能夠抑制果皮細胞結構的崩解,與前人研究結果相似[18]。前人研究發現,外施GA3后,浮皮果果皮中原果膠含量降低,細胞壁PG 和Cx 活性升高,細胞膜脂過氧化作用增強,果實衰老加快,有效降低了PE 活性和浮皮率[19,29]。Huai等[30]研究發現,葉面噴鈣也能抑制細胞壁物質的降解,提高果皮硬度從而降低柑橘皺皮果發生率。本研究中,GA3與PDJ 處理極顯著增加了果皮中原果膠和半纖維素含量,并且纖維素含量下降的幅度也低于CK,相應的細胞壁水解酶類(PME、Cx)活性及控制相關酶合成的基因表達量也極顯著低于CK,與前人研究結果類似[30],即GA3與PDJ 處理延緩了細胞壁結構的崩解,從而抑制果實浮皮的發生。此外,研究顯示,通過抑制果皮膜脂過氧化程度加重,降低果皮中POD 活性,能使南豐蜜橘果實浮皮空間率顯著降低[27],說明膜脂過氧化化程度也與果實浮皮密切相關。本研究同樣發現,GA3與PDJ 處理后果皮中CitPOD12基因表達量極顯著低于CK,表明果皮細胞膜脂過氧化程度降低,從而抑制了果實浮皮指數的增加。
乙烯可加劇柑橘果實浮皮,而抑制乙烯的生物合成則能降低浮皮的產生[21-22]。植物對乙烯敏感性的高低可通過乙烯受體基因表達水平的調控來實現,按照負調控模型的觀點,受體表達水平越高,植物響應乙烯的敏感性越差[31]。擬南芥[Arabidopsisthaliana(L.)Heynh.]受NaCl脅迫時,乙烯受體蛋白ETR1的減少導致了擬南芥對乙烯敏感性增強,反之ETR1 蛋白的增加會導致植物對乙烯敏感性減弱[32]。與之相似,在本研究中,GA3與PDJ 處理在FPQ 和FPH 時期均極顯著降低了乙烯合成前體ACC 含量,并且乙烯合成基因CitACO表達量也顯著(FPQ)和極顯著(FPH)低于CK,CitETR1基因表達量均極顯著高于CK,說明GA3與PDJ處理抑制果實乙烯的合成,同時使果實對乙烯的敏感性降低,進而抑制果實浮皮的發生。生長素對柑橘果實后熟衰老有抑制作用,在果實完熟過程中IAA 含量會持續下降。果實枯水通常也被認為是浮皮中的一種,枯水果實果皮中IAA含量顯著低于正常果,枯水果實果皮衰老程度高于正常果[33]。本研究同樣發現,經GA3與PDJ處理后,果皮中IAA 含量極顯著高于CK,同時果皮中生長素原初反應基因CitIAA27表達水平極顯著(FPQ)和顯著(FPH)高于CK。在Aux/IAA 家族基因中,部分成員在生長素處理早期被誘導表達,生長素消耗后逐漸恢復[34],園藝作物黃瓜中的CsIAA27基因也受生長素誘導表達[35]。顯然,本研究中GA3與PDJ 處理完熟栽培的溫州蜜柑,可以延緩果皮中IAA 含量下降,而較高濃度的生長素誘導了果皮中CitIAA27基因的表達,進而延緩了果皮衰老從而抑制浮皮的發生。
4 結論
本研究優化了GA3與PDJ 在抑制完熟栽培溫州蜜柑果實浮皮中的使用方法,即在DAF 155 和DAF 185分別噴施3.3 mg·kg-1GA3與25 mg·kg-1PDJ 可顯著抑制果實浮皮的發生,同時對果實著色未產生不利影響。明確了GA3與PDJ 抑制柑橘果實浮皮發生的生理及分子機制,即通過降低果皮中乙烯合成以及阻止IAA 的降解從而延緩果皮衰老,下調CitCx和CitPME等基因的表達降低PME和Cx活性,從而抑制果皮中細胞壁物質原果膠、纖維素及半纖維素的降解,防止果皮細胞壁的崩解,同時降低果皮細胞膜脂過氧化程度,從而抑制浮皮發生。
