光照對樹番茄幼苗碳、氮、磷、鉀元素積累與分配的影響
2024-04-24 13:54:48李樹萍李世民郭煥仙孫麗娟
西南農(nóng)業(yè)學(xué)報 2024年2期
李樹萍,李世民,郭煥仙,孫麗娟,董 瓊
(西南林業(yè)大學(xué)林學(xué)院/西南地區(qū)生物多樣性保育國家林業(yè)和草原局重點實驗室, 昆明 650224)
【研究意義】樹番茄(CyphomandrabetaceaSendt.)為茄科(Solanaceae)樹番茄屬(Cyphomandra)植物[1]。樹番茄生長迅速,成樹快,植株高大,分枝整齊,可以觀花賞果,可點綴美化庭園和綠化街道,經(jīng)濟(jì)和實用價值高,其蛋白質(zhì)含量高于一般果蔬,大多數(shù)的微量元素、維生素含量、氨基酸總量高于普通番茄[2],是一種頗具開發(fā)前景的植物資源[3]。樹番茄在幼苗期葉片容易受強(qiáng)光損傷和病蟲危害[4],生長受到抑制。研究不同遮陰條件下植物的形態(tài)結(jié)構(gòu)、元素的積累與分配情況為深入了解植物的適應(yīng)性提供重要意義。【前人研究進(jìn)展】光照是植物生長必不可少的條件之一,植物對光能的利用能力對其生長非常重要[5]。為了適應(yīng)不同光照環(huán)境,植物會通過調(diào)整形態(tài)結(jié)構(gòu)和生理生態(tài)特性來適應(yīng)一定范圍內(nèi)的光強(qiáng)環(huán)境[6]。在不同光照環(huán)境下,植物調(diào)整各部分的分配比例[7],把生物量分配給最需要的構(gòu)件[8],碳(C)、氮(N)、磷(P)和鉀(K)4種元素在植物生長、生理代謝及應(yīng)對環(huán)境變化時發(fā)揮重要作用,不但能反映出植物對營養(yǎng)的利用效率,而且反映其對生存環(huán)境的養(yǎng)分限制[9]。植物在自然光照和遮陰環(huán)境下,植株生長與礦質(zhì)元素的分配利用狀況有差異[1]。【本研究切入點】樹番茄幼苗強(qiáng)光下葉片容易損傷,而成年樹則表現(xiàn)出喜光特性。目前,云南德宏、西雙版納、保山等地都有栽培樹番茄,給當(dāng)?shù)剞r(nóng)民帶來了較大的經(jīng)濟(jì)收益。云南德宏通過探索樹番茄林下種植模式,在一定程度上可以減少樹番茄幼苗期的強(qiáng)光暴曬以及病蟲害的發(fā)生,但關(guān)于樹番茄幼苗在弱光下能否較好地生長及生理響應(yīng)尚缺乏系統(tǒng)研究。【擬解決的關(guān)鍵問題】通過研究樹番茄幼苗在不同遮光條件下的生理響應(yīng)機(jī)制,為其林下種植和速生豐產(chǎn)提供理論依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
試驗地點位于云南省昆明市西南林業(yè)大學(xué)樹木園內(nèi)(102°45′ E,25°03′ N),年平均氣溫16.5 ℃,年均降雨量700~1100 mm,海拔1954 m。
1.2 試驗材料
2021年4月選取長勢一致、生長健康的樹番茄幼苗開展試驗。苗期管理正常澆水除草。遮陰網(wǎng)為黑色遮陰網(wǎng)(包括2、3、6針),購買于市面;采用亮度檢測儀(SW-6023)測定遮光率。
1.3 試驗設(shè)計
試驗通過遮陰網(wǎng)來模擬4個不同的光照水平,每個光照水平為1個處理,共4個處理,即N1:無遮陰(100%NR,NR為自然光照,無遮陰網(wǎng));N2:輕度遮陰(75%NR,2針遮陰網(wǎng));N3:中度遮陰(50%NR,3針遮陰網(wǎng));N4:重度遮陰(25%NR,6針遮陰網(wǎng)),每個處理10株幼苗,重復(fù)3次,共120株幼苗。
1.4 測定指標(biāo)及方法
1.4.1 形態(tài)指標(biāo)測定 試驗開始于2021年4月,周期5個月,待試驗結(jié)束,測定所有苗木的苗高、地徑。
1.4.2 生物量的測量 試驗結(jié)束時將生長整齊一致的樹番茄苗木(每個處理選取4株)帶回實驗室,葉、莖、粗根(直徑≥2 mm)和細(xì)根(直徑<2 mm)分別置于烘箱中于105 ℃殺青30 min,80 ℃烘烤72 h至恒重,記錄每個處理組生物量。
1.4.3 C、N、P、K含量測定 參照鮑士旦[10]的方法,采用重鉻酸鉀氧化法測定全C含量,試驗樣品在濃硫酸-過氧化氫消解后過濾保存。全N采用奈氏比色法,在420 nm波長下測定;全P采用釩鉬黃比色法,在450 nm波長下測定;全K采用原子吸收分光光度法測定。
1.4.4 相關(guān)的指標(biāo)計算 相關(guān)指標(biāo)計算公式如下:根冠比=地下生物量/地上生物量,器官元素積累量=器官元素含量×器官生物量,器官養(yǎng)分分配比率(%)=器官養(yǎng)分積累量/植株總養(yǎng)分積累量×100。
2 結(jié)果與分析
2.1 不同光照處理對樹番茄幼苗苗高和地徑的影響
由圖1可見,不同遮光處理對樹番茄幼苗苗高和地徑影響顯著(P<0.05)。隨著遮光強(qiáng)度的增加,樹番茄幼苗苗高和地徑呈先增加后減小趨勢,在N3處理下樹番茄幼苗的苗高和地徑達(dá)到最大,全光照處理下,樹番茄幼苗苗高和地徑均小于其他遮陰處理,與N1處理相比,N2、N3、N4處理苗高分別增加7.94%、18.76%、6.43%,地徑分別增加1.30%、17.25%、7.80%。
2.2 不同光照強(qiáng)度對樹番茄幼苗生物量及其分配的影響
由圖2可見,不同光照處理對樹番茄幼苗各器官生物量、總生物量及根冠比影響顯著(P<0.05)。隨著光照程度減少,樹番茄幼苗的葉、莖、粗根生物量及總生物量先增加后減少,在N2處理下,樹番茄幼苗葉、莖生物量和總生物量達(dá)到最大值,分別為3.62、6.31、13.24 g;粗根生物量在N3處理下最大,較全光照處理增加40.17%。細(xì)根生物量和根冠比表現(xiàn)出相同的趨勢,隨著遮陰程度的增加,細(xì)根生物量和根冠比不斷減小,全光照下為最大值,其他處理較全光照處理細(xì)根生物量分別減少2.88%、29.81%、52.88%。
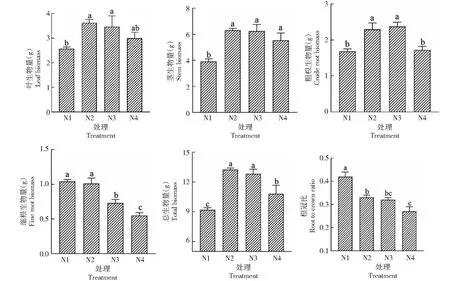
圖2 樹番茄幼苗各器官生物量及根冠比Fig.2 Biomass of each organ and root-crown ratio of C.betacea Sendt seedlings
如圖3所示,不同光照對樹番茄幼苗各器官生物量比影響顯著(P<0.05)。隨著遮陰程度增加,樹番茄幼苗葉生物量比先減少后增加,莖生物量比增加,細(xì)根生物量比不斷減少。較全光照處理下細(xì)根生物量比在各個處理下分別減少32.12%、49.42%、53.77%,莖生物量增加12.38%、14.74%、20.23%。遮陰處理影響樹番茄各器官生物量的分配比例,從而影響其生長。遮陰促進(jìn)樹番茄地上生物量的分配比例,減少地下生物量分配比例。
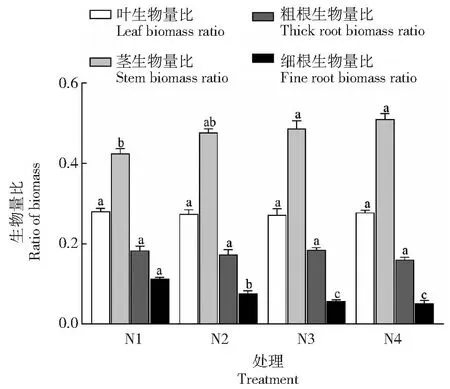
圖3 樹番茄各器官生物量比Fig.3 Biomass ratio of C.betacea Sendtes organs
2.3 不同光照強(qiáng)度對樹番茄幼苗C、N、P、K含量的影響
由表1可知,不同遮陰處理下,樹番茄幼苗各器官C、N、P、K含量不同。全光照處理下樹番茄幼苗各器官C含量差異不顯著(P>0.05),N2、N3、N4處理之間有顯著差異(P<0.05)。隨著光照程度減少,樹番茄幼苗的葉C含量先減少后增加,N2處理下最小。莖和粗根則隨光照強(qiáng)度減少先增加后減少,細(xì)根則趨勢不同,隨遮陰程度的增加不斷減少,在全光照下C含量最大。不同遮陰處理下樹番茄幼苗葉、細(xì)根N含量大于莖和粗根,表現(xiàn)為葉>細(xì)根>粗根>莖。樹番茄幼苗各器官P含量隨著遮陰程度的增加先增加后減少,在N2處理下達(dá)到最大值,葉片P含量在N4處理下最小,其他處理較N4分別增加24.20%、53.71%、17.77%。不同遮陰處理下,樹番茄幼苗K含量差異顯著(P<0.05)。不同器官K含量表現(xiàn)為葉>莖>細(xì)根>粗根。隨著遮陰程度增加,樹番茄幼苗的葉、莖、細(xì)根K含量呈上升趨勢,在N4處理下達(dá)到最大值。
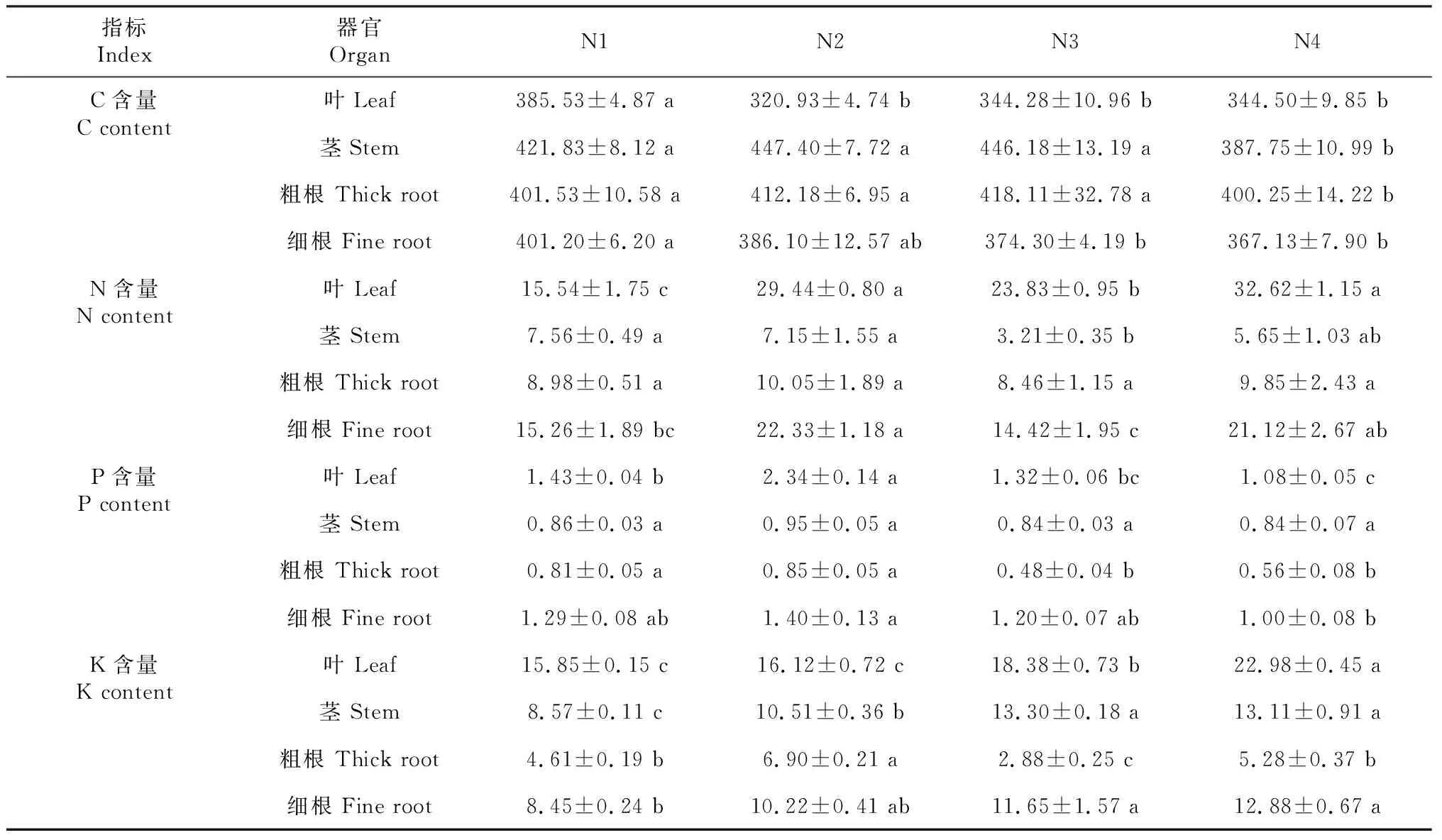
表1 樹番茄幼苗各器官C、N、P、K含量Table 1 The C, N, P and K content of C.betacea Sendt seedlings in all organs
2.4 不同光照強(qiáng)度對樹番茄幼苗C、N、P、K積累量的影響
由表2可知,樹番茄幼苗葉、莖的C、N、P、K積累量在不同遮陰處理下均大于粗根、細(xì)根。隨著遮陰程度增加,樹番茄幼苗的葉、莖、粗根C積累量先增加后減少,細(xì)根C積累量不斷減少。葉和粗根C積累量在N3處理下達(dá)到最大值,分別為1195.39、993.64 mg。樹番茄幼苗的葉、莖、粗根、細(xì)根N積累量在N2處理下達(dá)到最大值,與N1處理相比分別增加62.15%、33.93%、34.70%、31.71%。樹番茄幼苗的葉、莖、粗根、細(xì)根P含量隨著遮陰程度的增加先增后減,均在N2處理下達(dá)到最大值,在N4處理下最小,且葉、莖P含量大于粗根和細(xì)根,表現(xiàn)為葉>莖>粗根>細(xì)根。樹番茄幼苗葉K含量隨遮陰程度增加不斷上升,莖、粗根和細(xì)根總體上先增后減,粗根和細(xì)根在N2處理下達(dá)到最大值,而莖則在N3處理下最大。
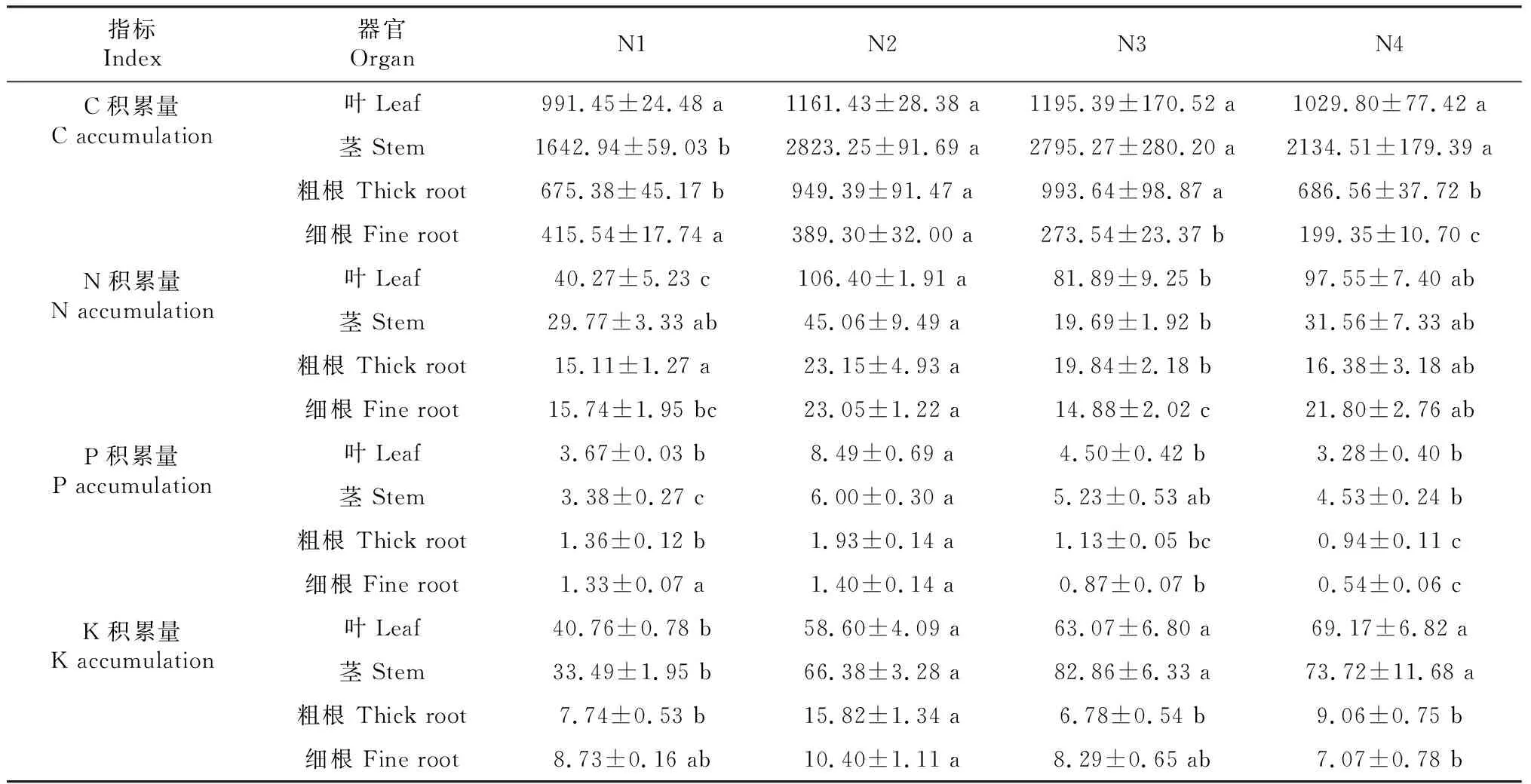
表2 樹番茄幼苗各器官C、N、P、K積累量Table 2 The accumulation of C, N, P and K in various organs of C.betacea Sendt seedlings
2.5 不同光照強(qiáng)度對樹番茄幼苗C、N、P、K分配比例的影響
如表3所示,遮陰處理增加了C、N、P、K向葉分配比例,減少了向粗根、細(xì)根的分配比例。隨著遮陰程度的增加,樹番茄幼苗莖C、K分配比例變化趨勢相同,在N3處理下達(dá)到最大值。莖N比例分配隨遮陰程度增加不斷減小,葉的變化趨勢與之相反,隨遮陰程度增加而增加,在N3處理下最大。莖P分配比例不斷增加,葉P分配比例先增大后減小,在N2處理下最大。
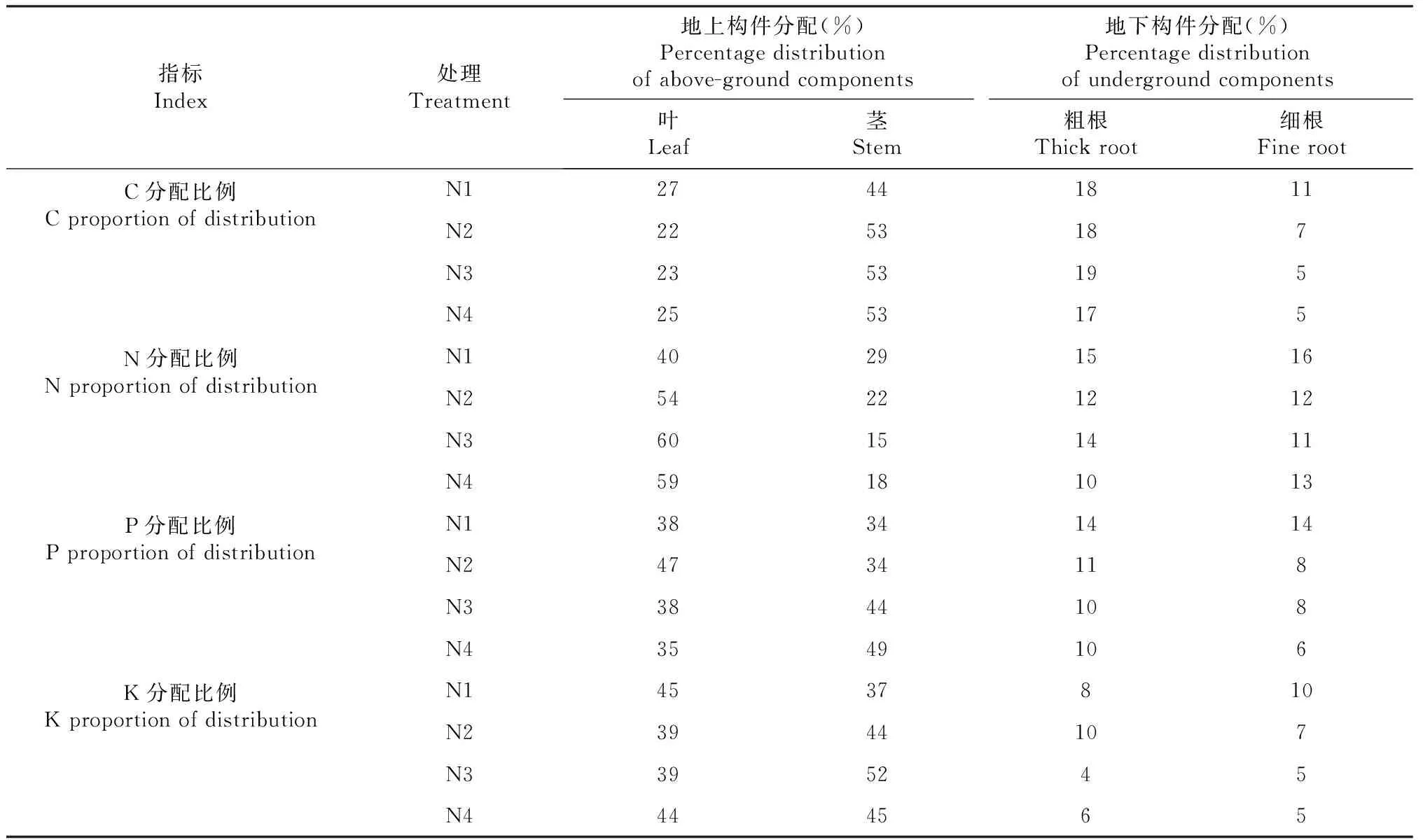
表3 樹番茄幼苗各器官C、N、P、K分配情況Table 3 The distribution of C, N, P and K in various organs of C.betacea Sendt seedlings
2.6 不同光照強(qiáng)度對樹番茄幼苗C、N、P、K相關(guān)性的影響
從表4可知,葉P含量和粗根P、K含量與細(xì)根生物量呈極顯著正相關(guān)(P<0.01,下同),葉K含量與莖K含量呈極顯著正相關(guān),莖N含量與粗根P含量呈極顯著正相關(guān),莖K含量與粗根P含量、細(xì)根生物量呈極顯著負(fù)相關(guān),粗根P含量與粗根K含量呈極顯著正相關(guān),莖生物量與粗根生物量呈極顯著正相關(guān),葉N含量與葉、莖、細(xì)根K含量和莖生物量顯著正相關(guān)(P<0.05,下同),葉P含量與莖C含量和莖、細(xì)根P含量呈顯著正相關(guān),莖C含量與細(xì)根P含量和粗根生物量呈顯著正相關(guān),莖N含量與莖K含量呈顯著負(fù)相關(guān),粗根C、N含量和細(xì)根N、P含量與各器官生物量、C、N、P、K含量無顯著相關(guān)(P>0.05)。

表4 樹番茄幼苗各器官C、N、P、K含量與生物量相關(guān)性Table 4 Correlation between C, N, P and K contents and biomass of various organs of C.betacea Sendt seedlings
3 討 論
光照的強(qiáng)弱直接影響植物的生長發(fā)育[11]。苗高和地徑是反映植物生長狀況的一項重要指標(biāo)[12]。植物通過改變生長策略來適應(yīng)光照條件變化時的環(huán)境。本研究中,遮陰能夠顯著提高樹番茄幼苗苗高、地徑生長量,50%NR處理下達(dá)到最大值。韋美華等[13]對楓香(Liquidambarformosana)幼苗研究發(fā)現(xiàn)在中度遮陰處理下苗高及地徑生長量均達(dá)到最大。樹番茄幼苗在中等光照強(qiáng)度下生長較好,過強(qiáng)或者過弱的光照則會對樹番茄幼苗的生長造成一定影響。全光照下,光照強(qiáng)度超出樹番茄所能利用的光照范圍,且樹番茄幼苗缺乏完善的光能保護(hù)機(jī)制,從而限制苗高和地徑的增長[6],而遮陰程度過大,樹番茄幼苗生長所需的光照不足,不能滿足其生長需求。生物量可以反映植物生長和生產(chǎn)力,改變生物量可以幫助植物適應(yīng)光照環(huán)境[14]。本研究中,隨光照強(qiáng)度減弱,樹番茄幼苗葉、莖生物量先增加后減少,葉、莖生物量在中度遮陰下最大,而細(xì)根生物量則逐漸減少,與其他處理相比,粗根生物量在重度遮陰下顯著降低,表明樹番茄幼苗地上部分生物量在適度遮陰下得到了累積,而地下生物量的累積減少,這與郭海霞等[15]研究發(fā)現(xiàn)適宜的遮陰能增加暗紫貝母(Fritillariaunibracteata)地上生物量的結(jié)果一致。植物通過改變生物量分配模式以適應(yīng)不同光照強(qiáng)度[13]。本研究發(fā)現(xiàn),遮陰后樹番茄幼苗為了競爭光照資源,增強(qiáng)縱向生長,將更多的生物量分配到莖上;弱光環(huán)境下,影響樹番茄幼苗生長的主要器官是莖,作為限制器官的細(xì)根對樹番茄幼苗生長的作用較小,因此樹番茄幼苗為了保持莖的生長而犧牲細(xì)根,徐飛等[6]對麻櫟的研究結(jié)果也與此相同。另外,植物通過調(diào)節(jié)根冠比以適應(yīng)不同的環(huán)境條件[16]。
本研究中,隨著光照程度減少,樹番茄幼苗根冠比逐漸減小,樹番茄幼苗為了適應(yīng)遮陰環(huán)境,捕獲更多的光能,提高對光能的吸收與利用,增加了地上部分構(gòu)件的生長,減少對地下部分的投入,更利于樹番茄幼苗構(gòu)建支持結(jié)構(gòu)和運輸結(jié)構(gòu)。
植物體內(nèi)C、N、P、K元素水平在一定程度上反映植物在不同生長階段的營養(yǎng)狀況以及對環(huán)境的適應(yīng)能力[17]。在植物的生長發(fā)育過程中,所能獲得的營養(yǎng)資源是有限的,為提高自身的適應(yīng)性,需要對有限的資源進(jìn)行分配。為了更好地適應(yīng)環(huán)境,植物會改變自身對資源的利用和分配[18]。本研究中,不同遮陰程度下樹番茄幼苗各器官C含量相對穩(wěn)定,葉片的C含量低于莖,葉片C含量用于葉的C同化。在不同遮陰程度下,葉片需要進(jìn)行光合作用以及呼吸和蒸騰作用,需要更多的N、P來支持合成光合色素等物質(zhì),而莖含有大量的木質(zhì)素和纖維素[19],有支持水分和營養(yǎng)物質(zhì)的運輸作用,不會儲存過多的N、P,因此植物會將更多的N、P分配給光合器官而不是莖。本研究中,隨著遮陰程度的增加,樹番茄幼苗各器官K含量總體上不斷增加,葉片K含量最高。K含量對植物的抗逆性有一定作用,還能提高植物的競爭力[20]。葉片含鉀量的增加在一定程度上有助于維持遮陰條件下的光合作用。充足的葉片鉀含量可改善植物功能和耐陰樹木對過度輻射的抵抗力,同時能促進(jìn)植物葉綠素合成,提高植物碳水化合物向經(jīng)濟(jì)器官的運輸效率[21]。
本研究中,樹番茄幼苗葉和莖C、N、P、K積累量在不同遮陰處理下均大于粗根、細(xì)根。葉片進(jìn)行光合作用時需要大量的N、P支持,提高光合速率,導(dǎo)致分配到葉片中的N、P高于其他器官,而莖中的N含量主要用于莖的吸收和運輸物質(zhì)[22],因此,葉和莖對上述元素積累量相對較高。K元素參與植物的各種生理與生化過程。植物的K含量與其自身的抗旱、抗病害能力密切相關(guān)。樹番茄在不同遮陰程度下K積累量增加,說明樹番茄會選擇性地吸收更多的K來應(yīng)對環(huán)境的變化。
4 結(jié) 論
不同光照強(qiáng)度對樹番茄幼苗苗高和地徑生長、元素含量以及各器官生物量均有顯著影響。50%遮陰可使樹番茄幼苗苗高、地徑、生物量、養(yǎng)分含量和積累量均有所增加,但過度遮陰會在一定程度上抑制幼苗生長。遮陰處理增加了C、N、P、K向葉、莖的分配比例,減少了向粗根、細(xì)根的分配比例。因此,室外栽培、林下推廣種植以及園林應(yīng)用中選擇50%遮陰環(huán)境有助于樹番茄生物量及C、N、P、K含量和積累量的增加。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
