奶牛乳房炎相關IFNE基因的克隆、表達及功能研究
2024-04-24 14:12:56王正興羅仍卓么王晉鵬包斌武王興平
西南農業學報 2024年2期
王正興,羅仍卓么,王晉鵬,包斌武,王興平
(1.寧夏大學動物科技學院,銀川 750021;2.寧夏回族自治區反芻動物分子細胞育種重點實驗室,銀川 750021)
【研究意義】奶牛乳房炎通常是由病原體感染乳腺組織引起的炎癥反應,是奶牛最常見的疾病之一[1],可導致奶牛的產奶量下降和乳品質降低[2-3],使得牛奶的經濟效益下降,并且會增加疾病處理成本,以及奶牛過早淘汰而增加總體生產成本,給牧場造成巨大的經濟損失。【前人研究進展】研究發現,造成奶牛乳房炎的病原體大致分為傳染性病原體和環境性病原體,其中大腸桿菌(Escherichiacoli)、金黃色葡萄球菌(Staphylococcusaureus)、無乳鏈球菌(S.agalactiae)和發育不良鏈球菌(S.pyogenes)被認為是傳染性病原體,可在乳腺中定植,感染乳腺組織,產生炎癥[4-8]。在眾多病原體中,大腸桿菌被認為是導致乳房炎的主要病原體之一[9]。脂多糖(LPS)是革蘭氏陰性菌細胞壁的主要結構成分,也是大腸桿菌的內毒素,乳腺組織接觸LPS會引發炎癥的發作[10]。炎癥基因表達的一個重要轉錄調節因子是轉錄的Janus激酶/信號轉導器和轉錄激活劑(JAK-STAT)[11],在特異性皮炎免疫應答中,胸腺基質淋巴細胞生成素(Thymic stromal lymphopoietin,TSLP)是一種具有致炎作用的細胞因子,其介導的反應通過JAK-STAT信號通路[12]。據報道,JAK-STAT在LPS、干擾素-γ(Interferon γ,IFNγ)和其他細胞因子激活的炎癥信號通路中起作用[13]。JAK-STAT調節編碼促炎細胞因子和趨化因子靶基因的轉錄[14]。大量哺乳動物細胞因子,包括白細胞介素和干擾素,通過JAK-STAT信號通路來調節細胞內信號傳導[15]。干擾素(Interferon,IFN)是參與細胞生長調節和免疫激活的多功能分泌蛋白[16]。根據感染病原體的不同,IFN信號傳導可能對細菌感染產生保護作用或有害作用[17]。干擾素ε(IFNE)是一種I型IFN[18],Choo等[19]在對哺乳動物如穿山甲的研究中發現,IFNE僅在上皮細胞中表達,主要在粘膜免疫、皮膚感染以及炎癥方面也起著一定的作用[20-21]。【本研究切入點】目前,奶牛IFNE的序列信息以及分子功能尚不清楚。【擬解決的關鍵問題】本試驗克隆奶牛IFNE基因的編碼區(CDS),并通過qPCR及RNAi技術對IFNE基因在奶牛乳腺上皮細胞(bMECs)炎癥反應中的表達模式及功能進行分析,以期為奶牛乳房炎的分子治療提供參考。
1 材料與方法
1.1 試驗材料
1.1.1 主要儀器 采用高速冷凍離心機(Eppendorf)、普通PCR儀(Bio-Rad公司)、多功能酶標儀(Bio-Rad公司)、電泳凝膠成像儀(Bio-Rad公司)、核酸凝膠電泳儀(Bio-Rad公司)、CFX 96 Touch 熒光定量PCR儀(Bio-Rad公司)和倒置熒光顯微鏡(Olympus)。
1.1.2 主要試劑 采用DMEM/F12培養基(Hyclone)、PBS(Hyclone)、0.25%胰酶消化液(Hyclone)、胎牛血清(BI)、Trizol(Takara)、PCR 2×TaqMaster Mix(諾唯贊生物科技有限公司)、膠回收試劑盒購(Omega公司)、2×M5 HiPer SYBR Premix EsTaq(北京聚合美生物科技有限公司)、X-treme GENE HP DNA轉染試劑(Roche)、OPTI-MEM(Gibco)、2×TaqMaster Mix(諾唯贊生物科技有限公司)和RNase-free ddH2O(北京天根生化科技有限公司)。
1.1.3 奶牛乳腺上皮細胞 選用寧夏回族自治區反芻動物分子細胞育種重點實驗室凍存的經鑒定的bMECs[22]。
1.2 研究方法
1.2.1 乳腺上皮細胞培養及炎癥誘導 在液氮中取出bMECs后接種于含10% FBS的DMEM/F12培養基,在37 ℃含5% CO2和100%飽和濕度條件下培養,當單層細胞融合達70%時,采用LPS(工作濃度為50 ng/μL)誘導bMECs,使其產生炎癥反應。在LPS誘導的第0、3、6、12和24小時收集細胞,用于RNA提取。
1.2.2 引物設計、RNA提取與cDNA合成 利用Primer Permier 5.0軟件設計IFNE基因的PCR擴增引物(表1)。采用Trizol法提取LPS誘導的bMECs總RNA。用瓊脂糖凝膠電泳儀和多功能酶標儀檢測RNA的完整性和濃度。選擇OD260/280值在1.8~2.0且較完整的RNA進行反轉錄得到cDNA。反轉錄體系及條件:①去除基因組DNA的反應:向無RNA酶離心管加入5×g DNA Eraser Buffer 2.0 μL,gDNA Eraser 1.0 μL,總RNA為1000 ng,最后加RNase free ddH2O 補至10.0 μL,混勻,瞬時離心,反應條件為42 ℃ 15 min,85 ℃ 5 s,4 ℃保存。②反轉錄反應:向步驟①中10 μL的反應液中加入PrimerScript RT Enzyme mix I 1.0 μL,RT Primer Mix 1.0 μL,5×primerscript Buffer 2(for Real Time)4.0 μL,RNase free ddH2O 補至20.0 μL,混勻,瞬時離心,反應條件為37 ℃ 15 min,85 ℃ 5 s,4 ℃保存。

表1 引物序列Table 1 Primer sequences
1.2.3IFNE基因的PCR擴增與回收、測序 以單鏈cDNA為模板,利用基因特異性引物擴增IFNE基因片段。反應體系:2×TaqPCR Master 10.0 μL,上、下游引物各0.8 μL,cDNA 1.0 μL,RNAse free ddH2O補至20.0 μL。PCR反應條件:95 ℃預變性3 min; 95 ℃變性15 s,54 ℃退火15 s,72 ℃延伸1.5 min,39個循環; 72 ℃終延伸5 min,4 ℃保存。PCR產物經1.5%瓊脂糖凝膠電泳進行檢測后,用DNA凝膠回收試劑盒進行擴增目標片段的回收,與pMDTM19-T載體進行連接。反應體系:Solution buffer 5.0 μL,純化產物4.0 μL,pMDTM19-T 1.0 μL,4 ℃連接過夜,將連接產物進行細菌轉化,并通過菌液PCR,選取擴增出條帶單一且大小符合預期的菌液,送往楊凌天潤奧科生物科技有限公司進行菌液測序。
1.2.4IFNE基因的生物信息學分析 根據DNAMAN軟件將IFNE基因菌液測序所得結果與GenBank數據庫進行比對,并利用ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)在線軟件分析基因的開放閱讀框,并推導其所編碼的氨基酸的序列;利用在線工具ExPASy-ProtParam(https://web.expasy.org/protparam/)分析IFNE基因所編碼蛋白質的理化性質;利用在線工具Conserved Domains(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)對IFNE蛋白進行保守結構域分析。
1.2.5IFNE基因干擾片段的合成與轉染 為探究IFNE基因對bMECs炎癥反應中的影響,委托生工生物工程(上海)股份有限公司合成3個奶牛IFNE基因的特異性干擾片段(si-152、si-653和si-730)以及對照組si-NC片段(表2)。細胞轉染方法:將生長狀態良好而且無污染的bMECs接種至6孔細胞培養板中,當單層細胞融合達70%時,配制甲液(si-干擾組:5 μL 干擾片段+100 μL OPTI;NC組:5 μL si-NC+100 μL OPTI)和乙液(3 μL X-tremeGENE HP轉染試劑+100 μL OPTI);甲乙液均靜止5 min,將甲和乙液等體積混合,靜止20 min,每組進行3個重復;轉染24 h后,采用熒光倒置顯微鏡觀察轉染情況;轉染48 h后,在6孔板中加入1 ug/mL的LPS刺激bMECs炎癥的發生,6 h后收集細胞進行RNA提取。
表2 奶牛IFNE的干擾片段序列Table 2 Interference fragment sequences of IFNE in dairy cows
1.2.6 qPCR實驗 采用Primer Permier 5.0軟件設計qPCR引物(表1),利用qPCR技術檢測IL-6、IL-8、IL-1β、IFNE、IFNB、IFNK、CASP3、CASP9、BAD和IL-10RB基因mRNA的表達水平。qPCR反應體系:2×M5 HiPer SYBR Premix EsTaq10.0 μL,上、下游引物各0.8 μL,cDNA 1.5 μL,RNase free ddH2O補至20 μL;反應條件:95 ℃ 30 s,95 ℃ 5 s,55 ℃ 30 s,40個循環。
1.2.7 數據統計分析 以GADPH為內參,所有試驗均進行3個重復,采用2-△△Ct法計算各檢測基因的表達量,通過GraphPad Prism 8.3.0軟件的獨立樣本T檢驗進行兩組間差異顯著性分析。P<0.05表示具有統計學意義,*表示差異顯著(P<0.05);**、***和****表示差異極顯著(P<0.01,P<0.001和P<0.0001)。
2 結果與分析
2.1 奶牛IFNE基因編碼區的克隆
利用IFNE基因PCR引物進行擴增,擴增產物經瓊脂糖凝膠電泳檢測,結果顯示,有清晰且單一的目的條帶,其擴增片段大小為838 bp(圖1),該序列已遞交GenBank(ID:OQ909434)。測序結果顯示,測序序列各堿基均為單峰,且582 bp的CDS區序列與牛基因組序列比對結果的同源性為100%,表明本試驗成功克隆出奶牛IFNE基因的CDS區。利用ORF Finder對IFNE基因的測序序列進行分析發現,IFNE基因的CDS全長為582 bp(546~1127 bp),可編碼193個氨基酸(圖2)。
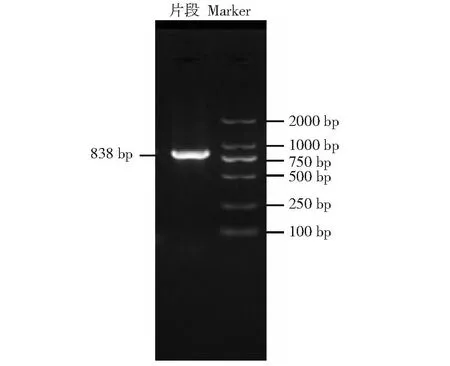
圖1 奶牛IFNE基因的擴增產物電泳Fig.1 Electrophoresis of amplification products of IFNE gene in dairy cows

下劃線表示擴增引物結合位置。The underline indicates the binding sites for primer sequences.圖2 奶牛IFNE基因所編碼的氨基酸序列Fig.2 Amino acid sequence encoded by IFNE in dairy cows
2.2 奶牛IFNE蛋白的生物信息學分析
利用ExPASy-ProtParam軟件對IFNE基因所編碼蛋白質的特性分析結果表明,IFNE蛋白的分子式為C1017H1631N279O288S10,原子總數為3225個,理論等電點為8.59。在IFNE蛋白的氨基酸組成中,亮氨酸(Leu)和谷氨酰胺(Gln)所占比例較高,分別為16.6%和11.4%;帶負電荷的殘基總數天冬氨酸(Asp)+谷氨酸(Glu)為19個,帶正電荷的殘基總數精氨酸(Arg)+賴氨酸(Lys)為22個;不穩定性指數(II)為48.56。跨膜結構域分析結果表明,奶牛IFNE蛋白存在于細胞內,無跨膜結構域。根據對其保守結構域的分析,IFNE蛋白質包含一個IFab超家族結構域,屬于I型干擾素成員之一,初步推測IFNE在炎癥反應的發生和發展過程中可能起到作用。使用DNAMAN軟件對本研究獲得的奶牛IFNE蛋白序列與野牛(XP_005887920.1)、水牛(XP_006054631.1)、綿羊(XP_011982517.1)、馬鹿(XP_043746624.1)、山羊(XP_013821322.2)、非洲狒狒(XP_047623512.1)、藍鯨(XP_036711910.1)、黑猩猩(XP_054513956.1)和馬(XP_014686768.1)9個物種進行同源性比對,結果顯示各物種之間IFNE蛋白的同源性較高(圖3),表明各物種的IFNE蛋白可能具有相似的生物學功能。
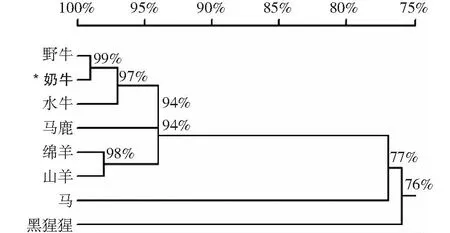
圖3 不同物種 IFNE 蛋白質的同源聚類樹Fig.3 Homologous cluster tree of IFNE proteins in different species
2.3 奶牛乳腺上皮細胞炎癥細胞模型的構建及IFNE基因表達量的檢測
通過體外培養bMECs(圖4-A),采用LPS誘導bMECs進行炎癥細胞模型構建。為驗證炎癥細胞模型是否構建成功,采用qPCR檢測炎癥因子IL-6、IL-8和IL-1β基因mRNA水平的表達量。結果表明,與0 h相比,IL-6和IL-1β在LPS誘導3、6、12和24 h的bMECs中表達量顯著上調(P<0.05)(圖4-B、4-D),IL-8在LPS誘導的3、6、12和24 h的表達量極顯著上調(P<0.01)(圖4-C),表明成功構建了細胞炎癥模型。同時,IFNE基因在LPS誘導bMECs 6 h的表達量與對照組0 h的表達量相比顯著上調(P<0.0001)(圖5),用于后續試驗。

A.培養的bMECs;B. IL-6在LPS誘導bMECs不同時間的表達量;C. IL-8在LPS誘導bMECs不同時間的表達量;D. IL-1β在LPS誘導bMECs不同時間的表達量。*表示差異顯著(P<0.05);**、***和****表示差異極顯著(P<0.01、P<0.001、P<0.0001)。下同。A. Culture of bMECs; B. Relative expression of IL-6 in LPS induced bMECs at different time; C. Relative expression of IL-8 in LPS induced bMECs at different time; D. Relative expression of IL-1β in LPS induced bMECs at different time. * indicates significant difference(P<0.05);**, ***and****indicate extremely significant difference(P<0.01, P< 0.001,P<0.0001).The same as below.圖4 bMECs炎癥模型構建的驗證Fig.4 Validation of the construction of inflammatory model for bMECs
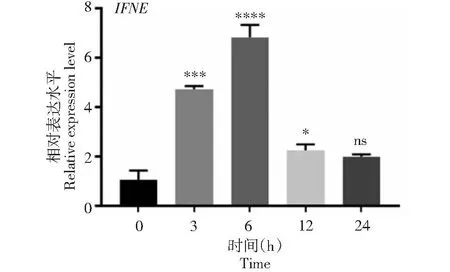
圖5 IFNE在bMECs炎癥誘導不同時間的表達Fig.5 Relative expression of IFNE of inflammation induction in bMECs at different times
2.4 IFNE干擾片段的篩選與檢測
為研究干擾IFNE后在LPS誘導的bMECs炎癥反應中基因的表達量,將IFNE的3條特異性的IFNE干擾片段轉染至bMECs后設置轉染Cy3標記的對照組(NC)。轉染24 h后,倒置熒光顯微鏡的成像結果顯示Cy3所標記的對照組(NC)的細胞中均有清晰的細胞輪廓(圖6-A、6-B),結果表明轉染成功。此外,qPCR結果顯示,與對照組(NC)相比,si-152、si-653和si-730均能夠極顯著抑制IFNE的表達(P<0.01),其中si-730片段的干擾效果最好(圖6-C)。因此,選擇si-730片段進行后續的IFNE基因干擾試驗研究。

A.Cy3標記的對照組(NC)轉染bMECs的明場圖像;B.Cy3標記的對照組(NC)轉染bMECs的熒光圖像;C.干擾片段轉染組細胞的IFNE表達量檢測結果。A.Cy3-labeled control(NC)transfected into bMECs; B.Fluorescent image of Cy3-labeled control(NC)transfected into bMECs; C.qPCR results of three interfering fragments. 圖6 IFNE干擾效率的qPCR檢測Fig.6 Detection of IFNE interference efficiency by qPCR
2.5 IFNE基因干擾對炎性bMECs中炎癥因子基因表達的影響
為研究干擾IFNE對bMECs炎癥反應的影響,本試驗轉染si-730至LPS誘導6 h的bMECs中,在此基礎上,采用qPCR法檢測bMECs內IFNE基因、炎癥因子IL-1β、IL-6和IL-8在IFNE基因干擾后的表達水平。結果表明,與對照組(NC)相比,干擾組的IFNE基因和IL-8的表達量顯著下調(P<0.05)(圖7-A、7-C),IL-6和IL-1β的表達量極顯著下調(P< 0.01)(圖7-B、7-D)。上述結果表明,干擾IFNE基因可通過下調促炎癥因子(IL-1β、IL-6和IL-8)的表達,從而緩解bMECs炎癥反應。

A. 干擾IFNE后IFNE基因在炎性乳腺上皮細胞中的表達量; B. IL-6在干擾IFNE后炎性乳腺上皮細胞的表達量;C. IL-8在干擾IFNE后炎性乳腺上皮細胞的表達量; D. IL-1β在干擾IFNE后炎性乳腺上皮細胞的表達量。A. Relative expression of IFNE in inflammatory bMECs after interference with IFNE; B. Relative expression of IL-6 in inflammatory bMECs after interference with IFNE; C. Relative expression of IL-8 in inflammatory bMECs after interference with IFNE; D. Relative expression of IL-1β in inflammatory bMECs after interference with IFNE. 圖7 干擾IFNE 對LPS誘導炎性bMECs主要炎癥因子基因表達的影響Fig.7 Relative expression of inflammatory factors in inflammatory bMECs after interference with IFNE
2.6 IFNE干擾對IFNE相關基因及細胞凋亡相關基因表達的影響
為研究干擾IFNE對IFNE相關基因及細胞凋亡相關基因表達的影響,利用qPCR技術檢測干擾IFNE后bMECs內JAK-STAT信號通路中的IFNE相關基因以及細胞凋亡相關基因的表達量(圖8),結果表明,與對照組(LPS+NC)相比,IFNB、IFNK和IL-10BR基因的表達量均顯著下調(P<0.05)。此外,CASP3、CASP9和BAD基因的表達量均極顯著下調(P<0.01)。上述結果表明,IFNE相關基因參與促進調節bMECs的凋亡,IFNE可能促進細胞炎癥反應。
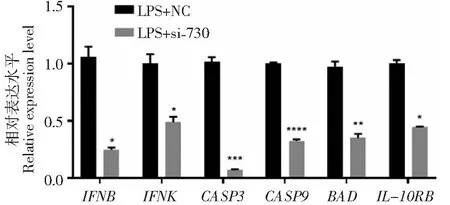
圖8 干擾IFNE對炎性乳腺上皮細胞內IFNE相關基因表達的影響Fig.8 Relative expression of IFNE related genes in inflammatory bMECs after interference with IFNE
3 討 論
3.1 IFNE基因克隆及蛋白生物信息學分析
奶牛乳房炎嚴重影響乳品質,對牧場的經濟收益造成損失,其中病原微生物E.coli是誘發臨床型乳房炎的主要病原體之一[24]。JAK-STAT參與調節細胞因子的產生,以及調控炎癥反應和免疫反應,其與乳房炎的發生、發展和調控密切相關[15],而IFNE基因在JAK-STAT信號通路中起著至關重要的作用[25]。本研究通過克隆測序獲得了奶牛IFNE基因CDS區序列,發現其可以編碼一條由193個氨基酸組成的不穩定親水性蛋白。在IFNE蛋白的保守結構域分析中,IFNE屬于IFab超家族中,IFN-α和IFN-β與IFNE屬于同一超家族基因,免疫系統長期暴露于IFN-α和IFN-β經常導致自身抗體和自身免疫性疾病的發生,如系統性紅斑狼瘡的發生[26]。基于IFNE超家族基因的分析,結果顯示IFNE與IFN-α和IFN-β在免疫反應中起到相似的作用,推測IFNE在炎癥反應的發生和發展過程中可能起到作用[26]。在山羊乳腺炎的研究中發現,干擾素(IFN)參與山羊乳房炎的調節[27],本研究奶牛與山羊的IFNE蛋白高度同源,推測奶牛IFNE蛋白可能參與調節bMECs炎癥反應,與奶牛乳房炎的發病率相關。
3.2 乳腺上皮細胞炎癥模型構建及IFNE的表達模式分析
bMECs不僅參與乳汁的合成與分泌,還與乳腺的應答有關[28]。外源性病原微生物感染奶牛乳腺時會被bMECs表面的特異性受體識別并會激活細胞免疫應答機制,產生炎癥細胞因子等介質,進而導致炎癥反應的發生[28]。有報道顯示,LPS可誘導bMECs發生炎癥反應[29],并且JAK-STAT信號通路可參與促炎細胞因子對LPS誘導的細胞炎癥的調節[20]。在奶牛乳腺炎的研究中,通常以LPS誘導的bMECs為研究材料,構建炎癥細胞模型[30]。此外,炎癥因子IL-6、IL-1β和IL-8都會引發對免疫穩態、炎癥、發育和代謝的生理調控[31]。本研究用LPS誘導bMECs產生炎癥反應,發現炎癥因子IL-6、IL-1β和IL-8的表達量都顯著上調,這與楊箭等[32]成功建立炎癥模型的結果相似,表明本研究建立的bMECs炎癥模型可靠。
3.3 IFNE基因在bMECs炎癥中的功能
IFNE基因是哺乳動物免疫反應中的重要調節因子[33]。迄今為止,有關于IFNE調控乳房炎的分子機制研究在奶牛和小鼠中還未報道。JAK-STAT信號通路內,IFNB、IFNK和IFNE同屬于干擾素家族基因[34],在人體免疫中,IFNB和IFNK在抗病毒與抗炎癥免疫和系統性自身免疫性疾病反應中發揮著重要作用[35-36]。IL-10RB基因是一種多效性因子,在早發性腸炎[37]和系統性紅斑狼瘡(SLE)[38]的發病機制中起重要作用。本研究發現,奶牛IFNE基因所編碼的蛋白質與山羊的同源性較高,推測奶牛IFNE蛋白在炎癥反應中也起著相似的功能。此外,本研究在炎性bMECs中進行IFNE基因干擾,可使IFNB、IFNK和IL-10RB基因表達量顯著下調,表明干擾奶牛IFNE基因可緩解bMECs的炎癥反應。CASP3與CASP9是細胞凋亡早期和晚期公認的標志基因,與乳腺炎風險有關[39-40];BAD是BCL2家族的促凋亡成員,其蛋白質是細胞凋亡的關鍵調節因子[41]。本研究發現在炎性bMECs中進行IFNE基因干擾可顯著下調CASP3、CASP9和BAD基因的表達量,表明干擾IFNE基因的表達,可抑制炎性bMECs的凋亡。
綜上所述,IFNE不僅參與調節bMEC炎癥反應,還參與調節bMECs增殖與凋亡,本研究結果可為奶牛乳房炎抗性育種提供基礎資料。
4 結 論
本研究成功克隆了奶牛IFNE基因的CDS序列,可編碼由193個氨基酸組成的不穩定性親水性蛋白質。IFNE基因在LPS誘導的bMECs炎癥反應中表達量顯著上調;干擾IFNE可下調炎癥因子IL-6、IL-1β和IL-8基因以及IFNB、IFNK、CASP3和CASP9基因的表達水平,進而緩解bMECs的炎癥反應。該結果可為奶牛乳房炎的分子調控機制研究提供參考。
