酶在植物多糖研究中的應用進展
2024-05-06 14:10:32劉麗陽胡彥波王西崔洋洋周佳寧王鈺帆劉多
食品研究與開發 2024年8期
關鍵詞:植物
劉麗陽,胡彥波,王西,崔洋洋,周佳寧,王鈺帆,劉多
(1.長春大學食品科學與工程學院,吉林長春 130022;2.長春師范大學生命科學學院,吉林長春 130123)
多糖是由多種單糖通過糖苷鍵組成的線性或帶有支鏈的生物大分子,廣泛存在于動植物、微生物、藻類中,與蛋白質和核酸同為生命活動必不可少的物質,具有免疫調節、抗腫瘤、抗氧化、降血壓、神經保護等生物活性,在生命活動中發揮著重要作用[1]。植物多糖的提取分離、結構修飾、結構解析及生物活性是當前研究的熱點問題。生物酶因其高效、專一的特性,被廣泛應用于多糖的研究中[2]。本文主要綜述生物酶在植物多糖輔助提取、結構修飾及結構解析方面的研究進展,以期深化對酶、植物多糖的理解以及開發多糖在食品、生物醫藥和其他領域的應用。
1 植物多糖概述
植物多糖是植物重要的結構組成部分及能量來源,在自然界來源廣泛,不同植物多糖分子構成及分子量差異化顯著,因此表現出不同的功效作用。明確其結構組成,解析結構與功效聯系已經成為植物多糖研究的熱點[3]。常見植物多糖如淀粉、纖維素、果膠等,早已成為人們日常生活的重要組成部分。
淀粉是葡萄糖的高聚體,分子式為(C6H10O5)n,其基本構成單元為α-D-吡喃葡萄糖。天然淀粉平均含有20%~30% 直鏈淀粉和70%~80% 支鏈淀粉。直鏈淀粉是由吡喃式葡萄糖殘基通過α-(1,4)葡萄糖苷鍵連接的線性大分子,相對分子質量為105~106,相當于500~6 000 個葡萄糖單位,在多數情況下只有1 個非還原端和1 個還原端,呈無分支的螺旋結構。支鏈淀粉是一種高度分子化的大分子,主鏈由10~60 個葡萄糖單元通過α-(1,4)糖苷鍵連接,側鏈由15~45 個葡萄糖單元通過α-(1,6)糖苷鍵連接組成,它的相對分子質量為107~109[4]。
纖維素是植物細胞壁的主要成分,分子式為(C6H10O5)n,不溶于水及一般有機溶劑,是自然界中分布最廣、含量最多的一種多糖,占植物界碳含量的50%以上[5]。其結構是由吡喃式葡萄糖殘基通過β-(1,4)葡萄糖苷鍵連接的線性聚合物,其中吡喃式葡萄糖基的構象為椅式,相鄰葡萄糖基在形成糖苷鍵時會旋轉180°,因此,纖維二糖被認為是纖維素的基本重復單元。
果膠是在植物細胞壁內的一種多糖,由多達17 種類型的單糖通過20 種以上不同的連接方式組成[6]。在果膠中,半乳糖醛酸是含量最高的單糖,其次是阿拉伯糖、半乳糖(galactose,Gal)和鼠李糖(L-rhamnose monohydrate,Rha)等單糖。天然果膠通常由以下結構域組成:同型半乳糖醛酸聚糖(homogalacturonan,HG)、鼠李半乳糖醛酸聚糖I(rhamnogalacturonan I,RG I)、鼠李半乳糖醛酸聚糖II(rhamnogalacturonan II,RG II)和木糖半乳糖醛酸聚糖(xylogalacturonan,XG)。
半纖維素是植物細胞壁中使纖維素和木質素相互貫穿的多糖聚合物,是由重復的二糖單位構成的不均一聚糖。它是由20 多種單糖組成的一種無定形的異質多糖,包括木糖、阿拉伯糖、甘露糖、半乳糖、葡萄糖、焦糖、葡萄糖醛酸和半乳糖醛酸[7]。在大多數植物中,半纖維素約占生物干重的15%~35%,平均約占硬木的26%,軟木的22%[8]。重要的半纖維素主要有木聚糖和甘露聚糖。
2 植物多糖研究中相關的酶
2.1 淀粉酶
淀粉酶是能夠水解淀粉中糖苷鍵,獲得葡萄糖、寡糖、糊精等產物的一類酶。淀粉酶一般是從細菌、真菌和植物中分離。根據反應機理、底物特異性和蛋白質序列,淀粉酶被分為糖基水解酶(EC3.2.1.X)和糖基轉移酶(EC2.4.X.Y),前者分裂α-(1,4)和α-(1,6)糖苷鍵,后者分裂α-(1,4)糖苷鍵,從而形成新的α-(1,3)、α-(1,4)或α-(1,6)鍵[9]。研究表明,淀粉酶主要有α-淀粉酶(EC3.2.1.1)、β-淀粉酶(EC3.2.1.2)、葡萄糖淀粉酶(EC3.2.1.3)、α-葡萄糖苷酶(EC3.2.1.20)、普魯蘭酶或支鏈普魯蘭酶(EC3.2.1.41)和環糊精糖基轉移酶(EC2.4.1.19),切割α-1,4 或α-1,6-糖苷鍵,導致淀粉降解,這些酶被大量用于麥芽糖糊精、改性淀粉或葡萄糖糖漿的生產等[10]。α-淀粉酶(EC3.2.1.1)是一類水解淀粉、糖原等葡聚糖分子中α-(1,4)糖苷鍵的水解酶。α-淀粉酶在糖苷水解酶(glycoside hydrolase,GH)13 家族、GH57 家族、GH119 家族及GH126 家族中均有分布,主要集中在GH13 家族[11]。β-淀粉酶(EC3.2.1.2)是一種外切型淀粉酶,能從淀粉分子的非還原性末端水解α-(1,4)糖苷鍵并順次切下一個麥芽糖單元,生成β-麥芽糖及大分子的β-極限糊精。該酶在作用于淀粉底物時,由于發生沃爾登轉位反應,使產物異頭碳由α-構型變成了β-構型,因此得名β-淀粉酶,在碳水化合物活性酶數據庫CAZy(https://www.CAZy.org/,包括GH13、GH57、GH70 和GH77[12])分類系統中,β-淀粉酶屬于GH13 家族和GH14 家族[13]。
2.2 纖維素酶
纖維素酶是一類能夠作用于纖維素分子的酶,根據酶的功能差異,可分為三大類:內切葡聚糖酶(EC3.2.1.4)、外切纖維素酶和β-葡萄糖苷酶(EC 3.2.1.21)[14]。內切葡聚糖酶分布在不同的糖苷水解酶(GH)家族上,如GH5、GH9、GH12、GH44、GH45、GH48、GH51 和GH74[9]。內切葡聚糖酶主要作用于纖維素的無定形區,隨機水解β-(1,4)糖苷鍵,將纖維素分子長鏈截短,產生含有不溶性非還原性末端的低聚糖,為纖維二糖水解酶的外切作用提供大量的反應末端。外切纖維素酶是一種于纖維素線狀分子尾部發揮功能的酶,能夠水解β-(1,4)糖苷鍵,每次從纖維素末端水解下來一個纖維二糖分子,由此也被稱為纖維二糖水解酶(cellobiohydrolase,CBH)[15]。在國際命名EC法中,外切纖維素酶有2 個編號,分別為EC3.2.1.91 和EC3.2.1.176。其中,編號為EC3.2.1.91 的外切纖維素酶從纖維素鏈非還原末端起始催化,即CBHI;編號為EC3.2.1.176 的外切纖維素酶從纖維素鏈還原末端起始催化,即CBHII[16]。β-葡萄糖苷酶是一種非專一性酶,它可以破壞多種β-糖苷鍵,也能水解纖維六糖、纖維三糖等低聚糖[17]。3 種纖維素酶協同作用,可將纖維素徹底水解成單糖。
2.3 果膠酶
果膠酶是一組降解果膠分子的酶的總稱,果膠酶可以由許多生物體產生,如細菌、放線菌、酵母和真菌。根據裂解位點將果膠酶分為3 類:(1)由聚半乳糖醛酸酶(polygalacturonase,PG)(EC3.2.1.15)組成的水解酶;(2)裂解酶/反式消除酶,包括果膠裂解酶(pectin lyase,PNL)(EC4.2.2.10)和果膠酸裂解酶(pectinase lyase,PL)(EC4.2.2.2);(3)果膠酯酶(pectin esterase,PE)(EC3.1.1.11)[18]。PG 是一類果膠水解酶,可專一性地切斷未發生甲酯化的兩個半乳糖醛酸單元間的糖苷鍵,從而可將果膠長鏈結構高效水解為小分子片段,是果膠降解中的重要酶類。PG 有內切酶(endo-PG)和外切酶(exo-PG)之分,endo-PG 可隨機切斷果膠多糖的α-(1,4)糖苷鍵,exo-PG 則從多糖鏈的非還原性末端逐個切斷糖苷鍵[19]。果膠裂解酶和果膠酸裂解酶都通過β-消除來裂解果膠平滑區兩個D-半乳糖酸殘基之間的α-(1,4)糖苷鍵。果膠裂解酶只攻擊高度甲基化的區域,而果膠酸裂解酶通常裂解甲基化程度低的鏈[20]。果膠酯酶包括果膠甲酯酶、果膠乙酯酶和原果膠酶,它們催化果膠進行脫酯化反應,這些酶中研究最多的是果膠甲酯酶(pectin methylesterase,PME)。
2.4 半纖維素酶
半纖維素酶是一類能有效水解半纖維素多糖的酶,大多數的纖維素都來源于微生物,主要的微生物半纖維素酶有木聚糖酶、葡萄糖醛酸酶、阿拉伯呋喃苷酶、半乳糖苷酶和甘露聚糖酶[15]。半纖維素酶根據其催化活性分為糖基水解酶、酯酶和碳水化合物解酶,它們分別催化糖苷鍵的水解、乙酸或阿魏酸側基的酯鍵的水解和糖苷鍵的裂解。通過它們的協同作用才能將半纖維素完全降解[21]。木聚糖酶(EC3.2.1.8)水解木聚糖主鏈上的β-(1,4)糖苷鍵,生成短鏈木聚糖。大多數已知的木聚糖酶屬于GH 家族10 和11(已知的基因序列超過300 個),大約有20 多個木聚糖酶基因分布在GH 家族5、8 和43 之間。短鏈木聚糖再由β-木聚糖酶(EC3.2.1.37)從非還原端水解下單個木聚糖。內切甘露聚糖酶(EC3.2.1.78)從甘露聚糖內部水解β-(1,4)糖苷鍵形成低聚甘露糖鏈,外切甘露聚糖酶(EC3.2.1.25)再進一步把低聚甘露聚糖水解成甘露糖[22]。還有一些酶如α-L-阿拉伯呋喃糖苷酶、α-D-葡萄糖醛酸酶、乙酰乙酸酯酶等參與到半纖維素的降解中。
絕大多數木聚糖酶限于糖苷水解酶家族10(GH10)和家族11(GH11),后來在GH 家族5、8、30、43、51、198 和141 中也發現了木聚糖降解酶。與木聚糖酶分布在9 個家族不同,甘露聚糖酶目前被限制在4 個家族,GH 家族5、26、113 和134。GH5 和GH26是大多數甘露聚糖酶所屬的兩個家族。表1 總結了半纖維素酶的基本信息。
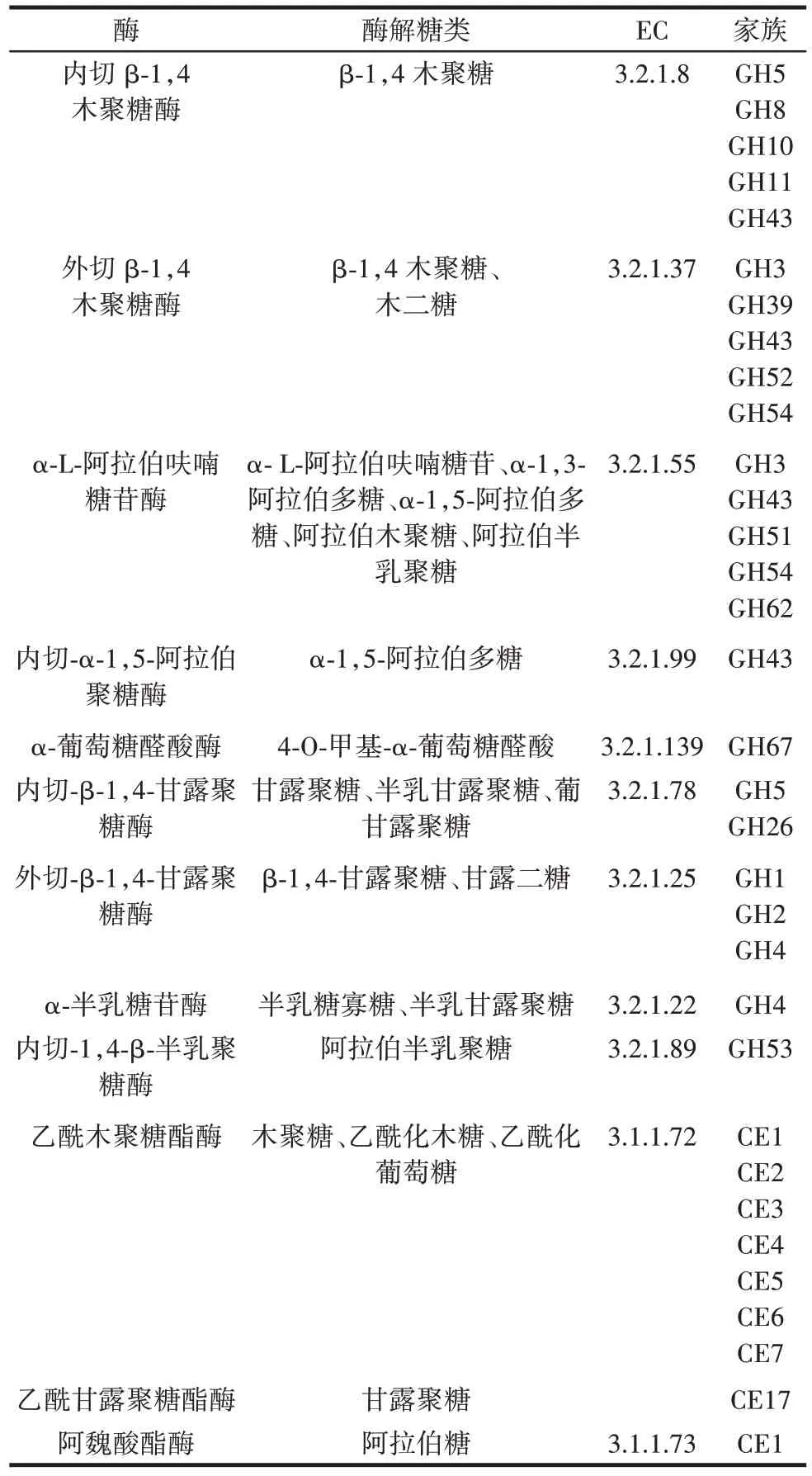
表1 半纖維素酶信息Table 1 Hemicellulase information
2.5 可用于多糖研究的其他酶
蛋白酶是一類裂解肽鏈中肽鍵的酶,廣泛存在于植物體內,蛋白酶的作用主要包括:1)消除錯誤折疊、修飾及定位的蛋白;2)為合成新的蛋白質提供氨基酸;3)通過限制性切割促使酶原成熟;4)降低關鍵酶和調節蛋白含量以控制新陳代謝平衡;5)切除已定位蛋白的定位信號[23]。根據肽鏈降解的起始位點可將蛋白酶分為肽鏈外切酶和肽鏈內切酶,肽鏈外切酶從肽鏈的一端開始切割,又可分為氨基肽酶和羧肽酶。肽鏈內切酶是從肽鏈的內部進行切割,并按照催化活性中心基團的特性進行分類。肽鏈內切酶的主要種類都已在植物中發現,包括絲氨酸蛋白酶(EC3.4.21)、半胱氨酸蛋白酶(EC3.4.22)、天冬氨酸蛋白酶(EC3.4.23)和金屬蛋白酶(EC3.4.24)[24]。
3 酶在植物多糖研究中的應用
近年來,酶在植物多糖研究中越來越受到關注,酶在植物多糖的提取、結構修飾以及結構分析中均取得了重要進展。酶法因其具有條件溫和、能耗低、提取效率高且不改變提取物分子結構的優點,被廣泛應用到植物多糖提取中;利用酶可以對植物多糖進行修飾改性以提高多糖的利用率或者改善其生物活性;酶更重要的是能在植物多糖結構分析中發揮巨大的作用,對多糖的結構與生物活性關系研究做出貢獻。
3.1 酶在植物多糖提取中的應用
生物酶法提取植物多糖是在傳統方法的基礎上,利用酶的專一性和高效性特點,根據植物細胞壁的組成,選擇相應的生物酶,將細胞壁中的纖維素、半纖維素、果膠等物質水解,從而使植物細胞中的有效成分更容易溶解擴散的一種提取方法。傳統方法苛刻的提取條件可能破壞多糖的分子結構,影響其功能性質和生物活性[25]。酶輔助提取是一種有巨大潛力的技術,具有能耗低、效率高、不改變提取成分分子結構的優勢。酶作為一種常見的生物催化劑,已被廣泛應用于不同植物組織中活性成分的提取[26]。植物中提取多糖常用的酶有α-淀粉酶、纖維素酶、果膠酶、半纖維素酶、蛋白酶等[27]。
根據試驗條件和提取方法的差異,研究者會選擇單酶或者復合酶加入其中以提高植物多糖的提取效率。Song 等[28]在紫菀多糖的提取中將紫菀混合液在α-淀粉酶、纖維素酶、果膠酶的最適合溫度下反應24 h,然后酶解液在100 ℃下加熱20 min 使酶失活,離心后的上清液在3 倍體積乙醇-20 ℃下過夜,粗多糖得率分別為果膠酶3.8%、纖維素酶3.4%、α-淀粉酶3.1%,其中,果膠酶和纖維素酶的得率高于熱水浸提法的得率(3.3%),這2 種酶可以破壞植物的細胞壁結構,使多糖等物質更好的溶解釋放。此外,果膠酶解多糖提取物具有更好的免疫刺激應激反應,因此,果膠酶輔助提取紫菀多糖在提高提取率及免疫活性方面具有更好的效果。Zhai 等[29]通過單因素試驗考察提取溫度、pH 值、酶解時間、液固比和果膠酶添加量對石榴皮多糖得率的影響,結果表明酶解時間、固液比和果膠酶添加量對石榴皮多糖的提取具有更顯著的影響,在響應面設計中得到最優條件為酶解時間19.70 min、固液比1∶20.5(g/mL)、果膠酶添加量0.68%時,石榴皮多糖最大提取效率為(27.3±0.8)%,與模型試驗預測的得率十分接近。酶解得到的石榴皮多糖在體外具有更高的自由基清除能力,試驗表明,酶解不僅能夠提高植物多糖的得率,也能一定程度提高多糖的純度。
單個酶對一些植物多糖提取有效,但多數情況下一種酶不能很好地分解植物細胞壁,通常需加入復合酶,如在纖維素酶、果膠酶、木瓜蛋白酶等共同作用下徹底破壞細胞壁的交聯組織,使有效成分更好溶解、釋放到溶液里。復合酶的使用能夠更好地破壞植物細胞中的物質(如細胞壁、纖維素、淀粉顆粒等)從而提高多糖的得率。
目前酶法更多的是與其他新型提取技術如超聲波輔助提取、微波輔助提取等相結合,如超聲-酶輔助提取法,酶水解植物細胞使細胞壁松動和分解,降低傳質阻力,促進細胞壁多糖和胞內多糖的溶解,同時去除蛋白質等部分雜質,隨后超聲波使溶劑滲透到細胞中,引起酶與底物碰撞的增加,加速有效成分的溶解[30]。與傳統的熱水提取法相比,該提取方法使多糖具有更高的得率、純度和生物活性,具有經濟、環保、高效的優點。這種通過酶處理和超聲降解的分子修飾技術可以改變多糖的空間結構,降低分子量,提高溶解度,從而影響多糖的生物活性。Olawuyi 等[31]在對秋葵多糖的提取中,用酶提取法和酶超聲聯合提取出的多糖與之前的熱水浸提法所提取多糖的性質進行了對比,發現在提取性能上,酶提取法的提取率比熱水浸提法低但是連續性更好,且酶解提取的秋葵多糖分子量低、純度高、單糖組成不同,但化學性質不變。酶-超聲聯合提取可促進生物活性物質的釋放,增強秋葵多糖的抗氧化活性和抑菌能力,擴大秋葵多糖在食品中的應用。Yang 等[32]用木瓜蛋白酶和纖維素酶1∶1 組成的復合酶提取紅芪多糖,在pH5、50 ℃條件下水浴2 h 后,105 W、60 ℃條件下提取1 h,處理3 次。酶解和超聲處理后提取的多糖得率以及碳水化合物含量分別為14.01% 和 92.45%,與之前的水提法(5.82% 和79.40%)對比有了很大的提升,二者都是以葡萄糖為主的雜多糖,對酶提取的紅芪多糖做細胞免疫活性測定,發現當紅芪多糖在200 ng/mL 以上濃度時,與熱水浸提法提取的紅芪多糖相比,能極顯著增強脂多糖誘導的淋巴細胞增殖,紅芪多糖的巨噬細胞免疫調節活性與多糖平均分子量呈正相關。酶解和超聲作用引起多糖的降解、接枝和交聯反應,改變其大小、空間構象和溶解度,影響其物理化學性質和生物活性。表2 總結了部分酶在多糖提取中的應用情況。
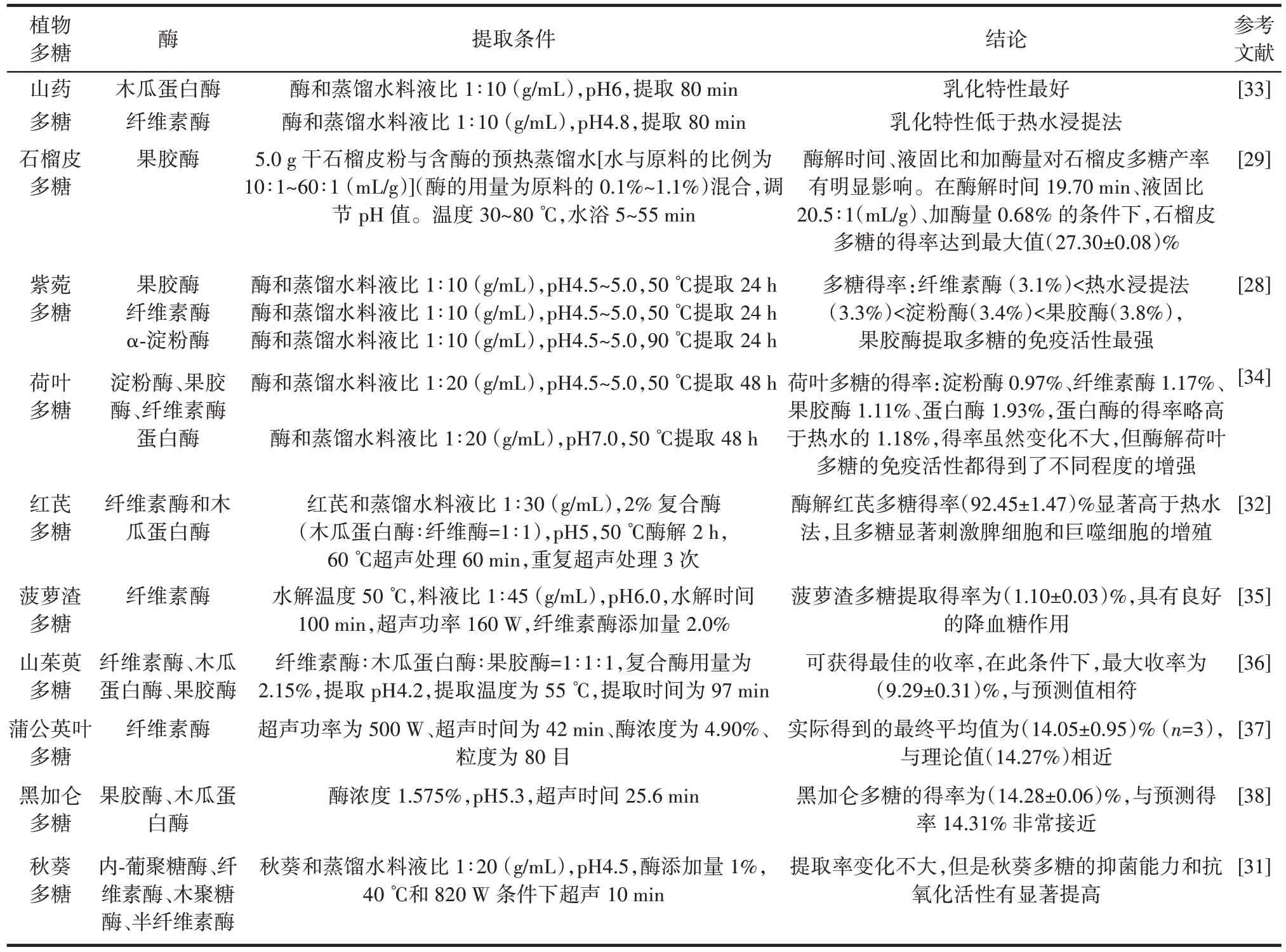
表2 酶在植物多糖提取中的應用Table 2 Application of enzymes in extraction of plant polysaccharides
3.2 酶法在植物多糖修飾中的應用
多糖的生物活性與其結構有著密切聯系,由于多糖結構的復雜性,多糖并不利于人體的消化吸收,低聚糖作為多糖的水解產物,本身和多糖有許多相似生物活性和結構,低聚糖作為多糖的替代品在食品領域受到越來越多的重視。目前,酸水解和酶解是多糖制備低聚糖的主要方法[39-40]。各種制備低聚糖的方法都有其獨特的特點,但也存在時間長、效率低、污染大的缺點。酶解法是一種較為成熟的多糖降解方法,具有降解條件溫和、酶切位點特異、低聚糖得率高、工藝簡單、重復性高、開發潛力大等優點[41]。
目前已有很多研究將生物酶法應用到低聚糖制備中,并取得了一定的成果。酶法修飾多糖會根據多糖的組成選擇合適的酶,通常使用果膠酶、纖維素酶、半纖維素酶等中的一種或多種酶來修飾多糖,得到更好生物活性的低聚糖或寡糖。Song 等[42]在對人參多糖進行提取時加入纖維素酶50 ℃下處理24 h、α-淀粉酶90 ℃處理24 h 以及纖維素酶和α-淀粉酶的聯合試劑處理24 h,最后得到了3 種多糖,與熱水浸提法相比,3 種酶輔助提取方法多糖的得率較低,但是酶解改變了多糖中各種單糖的比例,兩種酶聯合提取的多糖具有更顯著的體外免疫刺激特性,這表明兩種酶聯合可有效提取新鮮人參中新型功能性多糖。Hu 等[43]以桑葉多糖為原料用半纖維素酶進行降解得到了桑葉寡糖,其中與多糖相比,桑葉寡糖羰基、羥基和羧基含量增加,但分子量、表面張力、表觀黏度和熱穩定性降低。酶解還改善了桑葉多糖的觸變性能和結構恢復性能。酶解后桑葉寡糖結構和流變學行為的變化導致其抗氧化能力增強,促進兩歧雙歧桿菌、青春雙歧桿菌、鼠李糖乳桿菌和嗜酸乳酸桿菌的生長,表明酶修飾是一種能提高多糖生物活性的方法。Fang 等[44]用果膠酶、葡萄糖淀粉酶、纖維素酶、木聚糖酶和β-葡聚糖酶將江蘺多糖分別降解為低分子量多糖,酶解后的江蘺多糖的抗氧化活性均高于原多糖,其中β-葡聚糖酶降解的江蘺多糖的抗氧化活性最好,檢測其對H2O2誘導的人胎肺成纖維細胞(human fetal lung fibroblasts 1,HFL1)氧化損傷的細胞保護作用,結果表明江蘺多糖經酶預處理后,可顯著提高HFL1 細胞活力,降低活性氧和丙二醛水平,提高抗氧化酶活性和線粒體膜電位,減輕HFL1 細胞的氧化損傷,β-葡聚糖酶降解江蘺多糖可顯著提高其抗氧化活性,證明酶解法是使多糖獲得更好生物活性、具有開發潛力的新型方法,這也是酶在植物多糖降解的典型應用。酶解改性可以降低多糖的黏度和稠度指數,提高多糖的結晶度和流動行為指數[45]。在很多的研究中發現,經酶解處理后多糖的生物利用度以及抗凋亡、抗氧化和免疫活性增加,這是由于多糖在水解后暴露了許多活性基團所致。
單一的酶制劑使用較為廣泛,為了提升多糖的生物活性會利用復合酶對多糖進行降解,有研究用細菌或真菌中的胞內酶來降解糖以達到提升其生物活性的目的。復合酶相較于單酶酶切位點更加豐富,可以把多糖切割為更小的結構單元,使多糖暴露更多的活性位點。Liu 等[46]利用可食用菌胞內酶來降解米糠多糖,通過正交試驗優化得到了酶解米糠多糖的最佳條件,降解后米糠多糖的單糖組成比例發生改變,分子量降低,對自然殺傷細胞的毒性提升(12.01±0.08)%,且抗氧化試驗研究表明,降解后米糠多糖的自由基清除能力更強。表3 列出了酶在植物多糖修飾中的應用。多糖活性的提升與其分子量大小的改變、單糖組成比例、活性基團的暴露有密切聯系,利用復合酶或食用菌內酶來降解多糖相較于其他方法更加節省時間和成本。
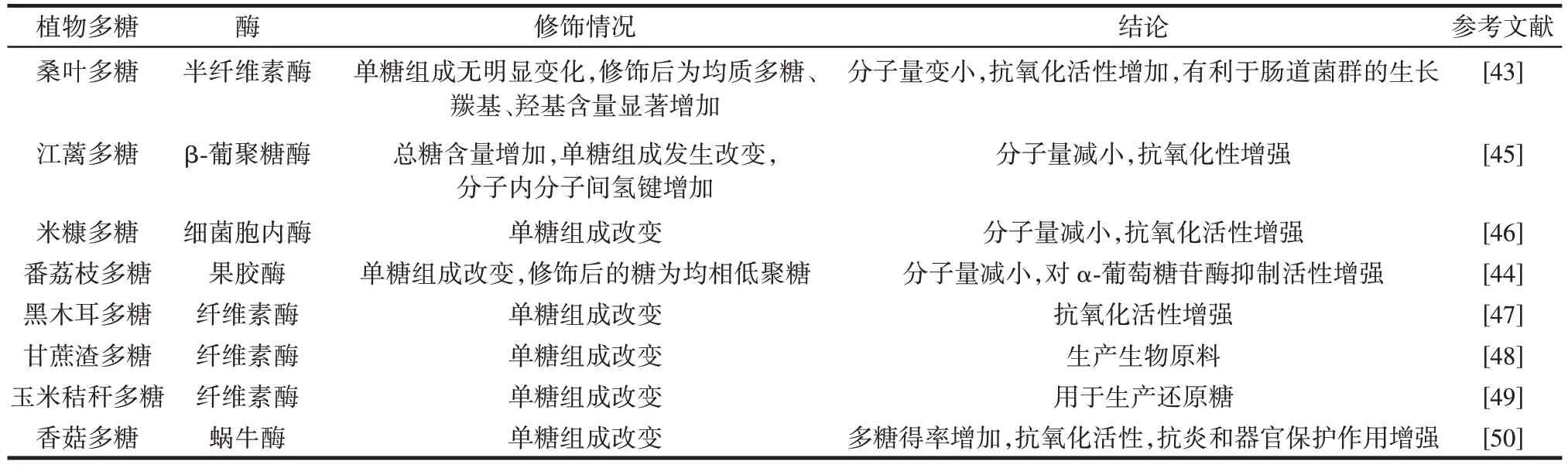
表3 酶在植物多糖修飾中的應用Table 3 Application of enzymes in modification of plant polysaccharides
3.3 酶法在植物多糖結構解析中的應用
多糖的結構表征和構效分析是研究多糖生物活性的關鍵,目前多糖的結構表征和構效分析仍然是一個挑戰。酶法分析多糖結構具有選擇性和高效性,對于結構未知的多糖,酶解法可以保持多糖分子重復單元的完整性,與酸水解相比酶解的特異性強,可控性比較高,污染少,條件溫和且利于實現,酸水解有時也會除去多糖的分支,改變多糖結構的完整性[21]。但在分析多糖分子中存在的各種單糖的差向異構化時存在局限性。要區分具有不同異構體的單糖,并確定該多糖的精細結構,需要采用其他儀器分析方法。
隨著科學技術的發展,酶法逐漸成為分析多糖結構的主要技術,利用內切糖苷酶可以將多糖降解成寡糖片段,并從混合物中通過選擇性純化特定的多糖。這些來自聚合物酶解的組分可以通過進一步的酶降解、甲基化分析、核磁共振光譜和質譜檢測。利用外切糖苷酶可以對多糖結構進行解析,確定末端糖基的連接方式,也可以對多糖結構進行修飾,研究多糖結構與活性之間的聯系。Ji 等[51]在人參堿性多糖的研究中,通過淀粉酶和木聚糖酶對純化后的人參多糖進行酶解,然后對得到的高聚組分和低聚組分進行分析。結果顯示,低聚組分中主要含有葡萄糖,同時還包含少量的阿拉伯糖、木糖和甘露糖,表明人參多糖中存在淀粉樣多糖。而高聚組分則主要由葡萄糖和木糖構成,同時也含有少量的半纖維素成分,如半乳糖、阿拉伯糖和甘露糖。通過紅外光譜中的β-連接糖殘基波段進一步證實了人參多糖中的淀粉樣多糖、果膠多糖和半纖維素多糖成分,通過特定的酶產物可更加快捷地分辨多糖的性質。酶法不僅能正面分析多糖的結構特征,也能利用酶解逆向推理出多糖的結構。Liu 等[52]對枸杞多糖進行了結構分析。他們使用熱水提取、脫色、色譜和透析等方法,從枸杞中提取并純化了酸性多糖。經過整體結構表征后,發現枸杞酸性多糖的主鏈由→4-α-GalpA-(1)→重復單元組成,同時存在一些支鏈,其中包括少量的→4-α-GalpA-(1)→和→2-α-Rhap-(1)→。枸杞多糖屬于典型的果膠分子。通過對其酶降解產物的分析,研究結果證實了枸杞多糖中含有高甲基果膠、寡聚半乳糖醛酸、寡聚阿拉伯糖醛酸和寡聚阿拉伯糖醛酸區域。這些結構特征與典型的果膠分子相符,因此,對枸杞多糖的構效研究可以與果膠的結構研究有進一步的相關性。
目前酶法分析多糖結構最常見的應用是根據酶切位點確定多糖的鏈接方式,從煙草葉片中脈提取的半纖維素為阿拉伯木聚糖,經纖維素酶處理后,得到復雜的低聚糖混合物,這些混合物的分離和表征為α-L-阿拉伯吡喃糖與D-木糖殘基的結合提供了第一個依據。為獲得結構更簡單的低聚糖,對阿拉伯木聚糖進行弱酸預處理(去除L-阿拉伯糖),再進行纖維素酶處理。對得到的三糖、五糖進行了化學性質和質譜分析,分別得到它們的結構信息,最終判斷其精細結構[53]。
4 展望
展望未來,酶法在多糖研究領域將持續發揮重要作用。隨著生物技術和工程學的進步,新的酶工程方法可以創造更高效、更專一性的酶,用于多糖的提取和純化,將大幅提高多糖的產量和純度。繼續為多糖的結構修飾提供有力工具。通過精確控制反應條件,包括溫度、pH 值、酶濃度等,研究人員可以通過調整多糖的結構實現特定的生物活性和功能。這將為制定多糖藥物和醫療材料打開新的可能性,有助于更深入地解析多糖的生物活性和功效。同時可以期待更多的多糖與生物分子的相互作用研究,以揭示多糖在藥物傳遞、免疫調節和抗氧化等方面的應用潛力。通過不斷創新和跨學科合作,采用酶法可以更深入地了解多糖的結構和功能,以及它們在醫藥、食品、材料等領域的廣泛應用,為人類健康和可持續發展做出更大貢獻。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
