基于代謝組學比較分析5個栽培種梨葉片代謝物的差異
2024-06-30 07:52:23焦慧君冉昆董冉董肖昌關秋竹王宏偉魏樹偉
果樹學報 2024年6期
焦慧君 冉昆 董冉 董肖昌 關秋竹 王宏偉 魏樹偉
摘? ? 要:【目的】為加強對梨葉片資源的藥用價值以及農副產品的開發利用,全面分析梨葉片中次生代謝物質的種類。【方法】通過代謝組學分析庫爾勒香梨、超紅、南紅、鴨梨、翠冠5個品種與福鼎大白茶的成熟期葉片的代謝物差異。【結果】PCA分析表明,代謝物種內聚類較為集中,種間聚類較遠;5個比較組特異性差異代謝物分別有55、155、54、59和28種;Heatmap分析表明,排名前50的差異代謝物在5個主栽梨葉片中的表達量具有顯著差異;黃酮和黃酮醇生物合成、ABC轉運和類黃酮生物合成等途徑是差異代謝物富集的主要通路,其中山梨醇、果糖、甘露醇、檸檬酸、水楊酸、馬來酸等含量顯著上調。【結論】不同栽培種梨葉片的代謝物在種類和表達量上均具有顯著差異,研究結果為開發利用梨葉片資源提供了理論基礎。
關鍵詞:梨;葉片;次生代謝物質
中圖分類號:S661.2 文獻標志碼:A 文章編號:1009-9980(2024)06-1111-14
Comparative analysis of metabolites in leaves of five pear cultivars based on metabonomics
JIAO Huijun, RAN Kun#, DONG Ran, DONG Xiaochang, GUAN Qiuzhu, WANG Hongwei, WEI Shuwei*
(Shandong Institute of Pomology, Taian 271000, Shandong, China)
Abstract: 【Objective】 Plant leaves are rich in secondary metabolites, mainly including phenols, terpenoids, alkaloids and other secondary substances, which have physiological functions in regulating plant growth, biological defense, anti-abiotic stress and so on. Pear is one of the important economic fruits of Rosaceae, and widespread in the world. China is one of the origins of pears, and has a long history of cultivation and rich variety resources. Pyrus pyrifolia, P. ussuriensis, P. sinkiangensis, P. communis and P. bretschnrideri are the main cultivated species of pear in China. Pears contain a wide variety of phenolic compounds, which would be beneficial for human health. Therefore, there have been many researches on the metabolite content of pear fruit. There are also abundant secondary metabolites in pear leaves, which are very important for the growth and development of fruit trees. However, there is a lack of systematic analysis of the metabolites in pear leaves. 【Methods】 In this study, we collected mature, healthy pear leaves in September, and carried them into lab. The leaves of Fuding Dabai tea rich in secondary metabolites were selected as control, and the differences of metabolites in the mature leaves of five cultivars Kuerle, Chaohong, Nanhong, Yali and Cuiguan were compared and analyzed by metabolomics. PCA analysis, Heatmap analysis, KEGG analysis and K-means analysis were used to analysis differential metabolites. 【Results】 A total of 9011 metabolites were detected under positive and negative ion mode. 11 747 peaks and 4987 metabolites were identified in the positive ion model, and 11 575 peaks and 4024 metabolites were identified in the negative ion model. The principal component analysis (PCA) showed that the detected intra-species clustering of metabolic species was relatively concentrated, while the interspecific clustering was relatively distant. The metabolites of the leaves of Chaohong were clustered far apart from those of the other four kinds of pear, which indicated the difference of metabolites in the leaves of occidental pear and oriental pear. In the oriental pear varieties, the metabolites of Kuerle fragrant pear were far from those of Yali, Cuiguan and Nanhong, and there were some differences. The number of differential metabolites of Kuerle, Chaohong, Nanhong, Yali, Cuiguan and Fuding Dabai was 1194, 1153, 1176, 1153 and 1164, respectively. There were 456, 468, 469, 481 and 446 kinds of significantly up-regulated differential metabolites in Kuerle, Chaohong, Nanhong, Yali and Cuiguan compared with Fuding Dabai, respectively, and 728, 685, 707, 672 and 718 kinds of significantly down-regulated differential metabolites. The number of common differential metabolites was 747, and the specific differential metabolites were 55, 155, 54, 59 and 28, respectively. The heatmap analysis showed that the expression levels of the top 50 differential metabolites were significantly different in the leaves of 5 cultivated pears. Among the top 50 differential metabolites, the number of metabolites in Yali leaves was the largest, with about 24 kinds, mainly including chrysanthemin, genistin, swertiajaponin, quercetin 3-lathyroside, picein, swertiajaponin, 6alpha-Hydroxycastasterone, glycyrrhetinic acid and protobassic acid. The correlation analysis could measure the degree of correlation between different metabolites, and further understand the interrelationship between metabolites in the process of biological state change. Therefore, we used Pearson correlation coefficient to measure the correlation between the top 50 differential metabolites with metabolite expression. It was found that 642 pairs of metabolites were positively correlated, and 633 pairs of metabolites were negatively correlated. We divided all the differential metabolites into 12 subclusters using K-mean analysis, and the results showed that the variation trend of differential metabolites in these 12 subclusters was basically the same. The number of differential metabolites in subclusters 1, 2, 6, 8, 10 and 11 was the most distributed, and the number was 229, 200, 104, 304, 256 and 81, respectively. The variation trend of differential metabolites in cluster 1, 2 and 8 was basically the same. The expression of metabolites in Chaohong leaves was quite different in subgroups 3, 5 and 7, and the contents of metabolites in Nanhong leaves was quite different in subgroup 12. The KEGG analysis were used to analyze the differential enrichment of the metabolic pathways between the leaves of Kuerle, Chaohong, Nanhong, Yali and Cuiguan and the leaves of Fuding Dabai. The 5 comparison groups were enriched in 78 KEGG pathways, there were 76, 71, 73, 70 and 74 KEGG pathways in the 5 cultivars, respectively. The flavone and flavonol biosynthesis (pxb00944), ABC transport (pxb02010) and flavonoid biosynthesis (pxb00941) were the main pathways for the enrichment of differential metabolite, among them, the contents of sorbitol, fructose, mannitol, citric acid, salicylic acid, maleic acid and methylmalonic acid were significantly up-regulated. Additionally, the zeatin synthesis (pxb00908) and monobacterial biosynthesis (pxb00261) were also metabolic pathways in which metabolites were significantly enriched in the leaves of Chaohong. 【Conclusion】To sum up, there are obvious differences in kinds and expressions of metabolites in the leaves of different pear cultivars, and this study could provide theoretical basis for the development and utilization of the leaf resources of pear.
Key words: Pear; Leaves; Secondary metabolites
植物次生代謝物是由植物次生代謝產生的非必需低分子有機化合物,來源廣泛、結構復雜,主要包括酚類、類萜、生物堿類和其他次生物質。酚類是芳香族環上的氫原子被羥基或功能衍生物取代后生成的化合物,以糖苷或糖脂狀態積存于液泡中,在植物體內廣泛分布,是重要的次生代謝物質之一,主要有莽草酸和乙酸-丙二酸等兩條合成途徑;類萜又稱萜烯類化合物,是以異戊二烯為單元組成,由乙酰CoA經甲羥戊酸和異戊二烯焦磷酸轉變而來。生物堿是存在于自然界中的一大類含氮雜環化合物,通常具有堿性,廣泛存在于植物體內,是氮素代謝的中間產物,由不同氨基酸衍生而來[1]。次生代謝產物廣泛參與植物的生長、發育和防御等生理過程。武予清等[2]報道黃酮類化合物是棉花植株中重要的抗病蟲害的物質,陸地棉的黃酮類化合物以異槲皮苷和蕓香苷為主。生物堿是含氮化合物的一種,大多數生物堿是植物的防御物質,對動物具有毒性。另外,許多生物堿具有藥用價值,如嗎啡、奎寧、長春花堿、利血平等。吳雅瓊等[3]采用代謝組學系統分析了黑莓、黑樹莓成熟果實中主要代謝物的差異,為高效利用果實中黃酮、花色苷等次生代謝物提供理論依據。張琴等[4]采用UPLC-MS/MS分析不同產地的蜂糖李果實中氨基酸等初生代謝物質的差異。魯忠富等[5]采用UPLC-MS/MS分析了瓠瓜果實鮮味的代謝物質差異,發現氨基酸及其衍生物和黃酮類等代謝物可能共同影響瓠瓜風味。另外,LC-MS技術在柑橘[6]、李[7]、獼猴桃[8]等果樹,番茄[9]、青花菜[10]和平菇[11]等蔬菜,以及蕎麥[12]、玉米[13]、棉花[14]等作物研究中應用廣泛。
梨屬于薔薇科蘋果亞科梨屬多年生落葉果樹,是我國重要的經濟水果之一。我國是梨起源中心之一,種質資源豐富,在長期的栽培馴化過程中逐漸形成了五大栽培種,分別是白梨(Pyrus bretschneideri)、砂梨(P. pyrifolia)、秋子梨(P. ussuriensis)、新疆梨(P. sinkiangensis)和西洋梨(P. communis)[15]。梨果實富含多種礦質元素、維生素、酚類、糖類和有機酸等營養成分。王宏等[16]采用代謝組和轉錄組關聯分析蘇翠1號與其親本翠冠和華酥果實成熟過程中的差異代謝物以及差異基因,發現調控糖代謝、山梨醇代謝、氨基酸代謝和脂肪酸代謝的基因在蘇翠1號中高表達,解析了蘇翠1號果實品質優良的分子基礎。趙芫[17]通過代謝組學從紅早酥果皮中篩選出211種黃酮類物質,發現乙烯處理后可以抑制異黃酮的積累和提高花青苷的含量,從而誘導果皮變黃。解敏等[18]采用GC-MS技術測定碭山酥梨成熟期果實品質,分析糖、有機酸和芳香物質的組分和含量。另外,高靜怡等[19]采用非靶向代謝組學分析砂梨品系水心病發病潛在的分子機制。代謝組學多用于分析果實品質和抗病性研究,尚未有系統分析梨葉片代謝物成分及含量的相關報道。
梨葉片是梨樹重要的組成部分之一,是進行光合作用的場所,輸送營養物質支持梨樹的生長發育過程。梨葉片中含有不同種類的次生代謝物質,影響梨樹抗性以及果實品質等。梨中關于次生代謝物質的研究大多集中于果實品質方面,缺乏系統全面鑒定葉片次生代謝物質的研究。為系統分析成熟葉片代謝產物種類和組分含量差異,筆者通過非靶向代謝組學技術,以茶葉片為對照,比較分析5個主栽品種梨葉片代謝物種類和含量的差異。庫爾勒香梨、超紅、南紅、鴨梨和翠冠5個梨品種,分別屬于新疆梨、西方梨、秋子梨、白梨和砂梨系統。5個主栽品種不論果實品質特征還是栽培區域等方面均存在明顯差異,間接說明在生長發育過程中葉片合成的次生代謝物在向果實轉化時存在差異,最終導致果實品質的差異。因此,筆者在本研究中的結果不僅為合理開發利用梨葉片資源、加工梨葉農副產品提供理論基礎依據,同時也為探究果實品質、抗性調控研究提供理論參考。
1 材料和方法
1.1 梨葉片采集
5個主栽品種(庫爾勒香梨,KEL;超紅,RC;南紅,NH;鴨梨,YL;翠冠,CG)的梨葉片采自山東省果樹研究所大河種質資源圃。于9月份采集成熟期的梨葉片,每個品種設置6組生物學重復,將采集的葉片放置于液氮罐中,帶回實驗室后放入-80 ℃冰箱中保存。
1.2 代謝物提取
首先,配置代謝物提取所需的溶液,包括內標溶液(L-2-氯苯丙氨酸,0.3 mg·mL-1;Lyso PC17:0,0.01 mg·mL-1,均由甲醇配置),70%和20%的甲醇溶液。提前在-20 ℃預冷提取液;分別稱取80 mg梨和茶葉片于2 mL的無酶離心管中,然后分別加入20 μL的內標溶液和1 mL 70%的甲醇溶液;加入2個小鋼珠,放置于-20 ℃條件下預冷2 min,然后放入研磨機(60 Hz)研磨2 min;超聲提取30 min后,置于-20 ℃條件下靜置20 min;在4 ℃、13 000 r·min-1條件下離心10 min,取300 μL上清液,揮發干凈后用400 μL 20%的甲醇復溶,渦旋30 s,超聲2 min。在相同條件下,離心10 min,然后用注射器吸取150 μL的上清液,使用0.22 μm有機相針孔過濾器過濾后,轉移到LC進樣小瓶,-80 ℃保存,直到進行LC-MS分析。另外,質控樣本(QC)由所有樣本的提取液等體積混合制備而成,每個QC體積與樣本相同。分析儀器為AB ExionLC超高效液相串聯AB TripleTOF 6600高分辨質譜儀組成的液質聯用系統。
1.3 差異代謝物的多元分析
刪除QC樣本中相對標準偏差>0.4的離子峰,PCA統計分析不同組樣本之間的差異,繪制質控樣本的代謝物強度;采用UNIFI 1.8.1 軟件采集原始數據,用Progenesis QI v2.3 軟件(Nonlinear Dynamics,Newcastle,UK)處理和鑒定原始數據,使用The Human Metabolome Database(HMDB)和Lipidmaps(v2.3)以及METLIN數據庫進行定性分析;然后基于有監督的降維方法,采用偏最小二乘判別分析(Partial least squares discriminant analysis,PLS-DA)法和正交偏最小二乘判別分析(Orthogonal projection to latent structures-discriminant analysis,OPLS-DA)法來更好地區分2組樣品的代謝組學特征和類別。通過OPLS-DA分析得到比較組之間差異代謝物的變量權重值(variable important in projection,VIP)。
1.4 差異代謝物分析
依據代謝物的多元分析和單變量分析的結果,篩選出5個比較組的差異代謝物。在OPLS-DA分析中,VIP值可用來衡量各代謝物的表達模式對各組樣本分類判別的影響強度和解釋能力。篩選的標準為OPLS-DA模型第一主成分VIP>1,t檢驗p<0.05。結合邁維云平臺(https://cloud.metware.cn/#/home)在線軟件對篩選出的差異代謝物進行火山圖、韋恩圖、Heatmap分析和相關性分析。具體參數為FC篩選閾值為1、p-value篩選閾值是0.05,VIP篩選閾值是1。依據 KEGG 數據庫(https://www.kegg.jp/)對差異代謝物進行代謝通路富集分析。
1.5 K-means分析
對篩選到的1319種代謝物在6種葉片中的含量趨勢進行聚類分析。使用邁維云平臺(https://cloud.metware.cn/#/home)在線軟件對篩選出的差異代謝物含量進行K-means聚類分析。由于差異代謝物含量差異范圍較大,因此對代謝物含量的數值進行數值轉換,對其分別取對數(log2),選取手動指定聚類方法,分類數目設置為12。
2 結果與分析
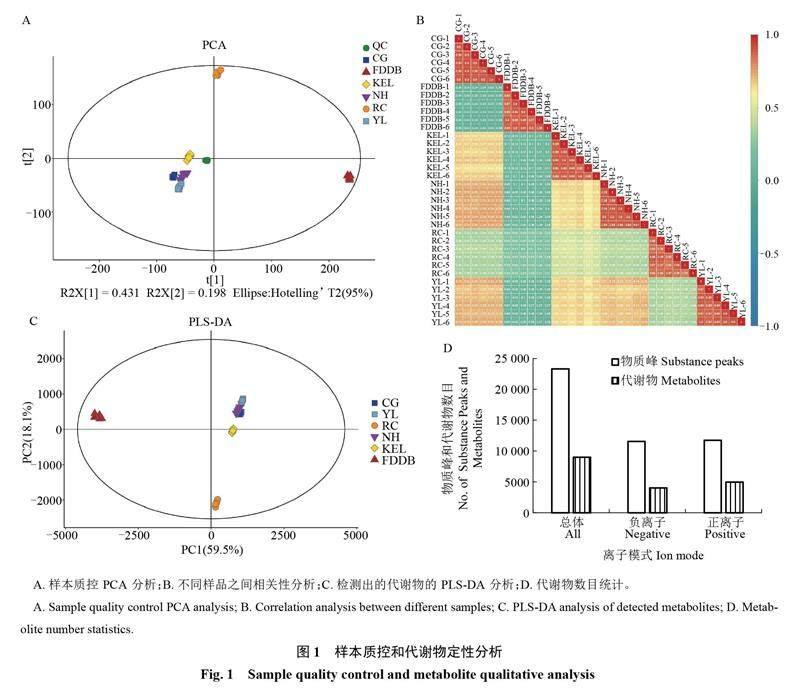
2.1 代謝組樣本質量控制和定性分析
采用PCA方法對所有質控樣本的離散程度進行分析,結果如圖1-A所示,QC樣本和6種葉片的組內樣本分別聚類緊密,穩定性和重復性較好。另外,對每組樣品的6個生物學重復進行組內相關性分析,結果顯示每組樣品的6組生物學重復均高度相關,相關系數均大于0.8;5組梨葉片的代謝物與茶葉葉片的代謝物的相關系數較小,為0.03~0.24(圖1-B)。在PLS-DA分析中,R2X(cum)均大于0.942、R2Y(cum)均是1,Q2(cum)均是1或0.999,說明該方法可以有效地區分種間和種內葉片代謝物的差異,并且可靠性較高。結果顯示,每組樣品的代謝物組內緊密聚集在一起,梨葉片與茶葉片的代謝物聚類相聚較遠,具有明顯差異。其中,超紅葉片的代謝物與其他4種梨葉片的代謝物聚類較遠,說明西洋梨和東方梨兩大種群梨葉片代謝物成分的差異。在東方梨品種中庫爾勒香梨與鴨梨、翠冠、南紅代謝物聚類分布較遠,具有一定的差異(圖1-C)。在正負離子模式下共鑒定出23 322個物質峰,相應的代謝物總共9011個。在正離子模式下鑒定出11 747個物質峰,4987種代謝物;在負離子模式下鑒定出11 575個物質峰,4024種代謝物(圖1-D)。
2.2 差異代謝物篩選及火山圖、韋恩圖分析
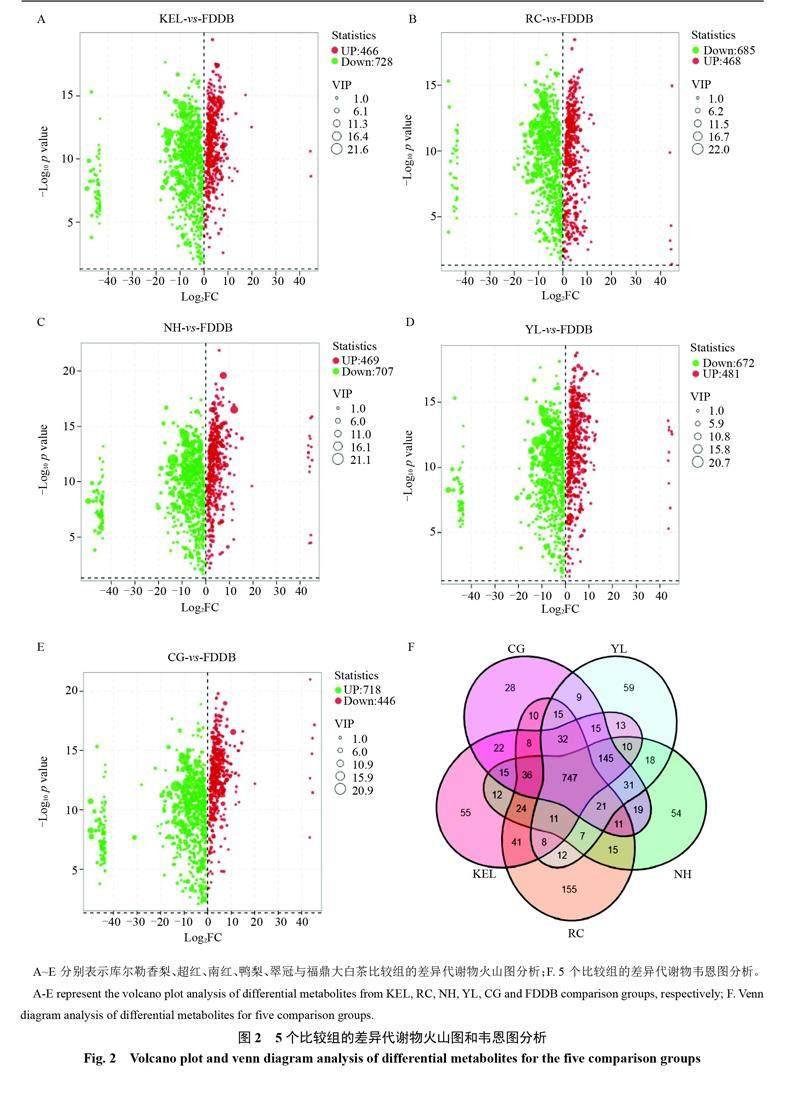
對檢測出的代謝物進行分析,庫爾勒香梨、超紅、南紅、鴨梨、翠冠分別與福鼎大白茶相比較,每個比較組分別檢測出1194、1153、1176、1153和1164種顯著差異的代謝物。依據差異代謝物的VIP、FC和p值繪制火山聚類圖。結果如圖3所示,庫爾勒香梨、超紅、南紅、鴨梨、翠冠與福鼎大白茶相比顯著上調的差異代謝物分別是466、468、469、481和446種,顯著下調的差異代謝物有728、685、707、672和718種(圖2-A~E)。韋恩圖結果表明,庫爾勒香梨、超紅、南紅、鴨梨、翠冠與福鼎大白茶相比,特異性的差異代謝物分別有55、155、54、59和28種,其中5個比較組共有的差異代謝物是747種(圖2-F)。
2.3 差異代謝物的定量Heatmap分析和相關性分析
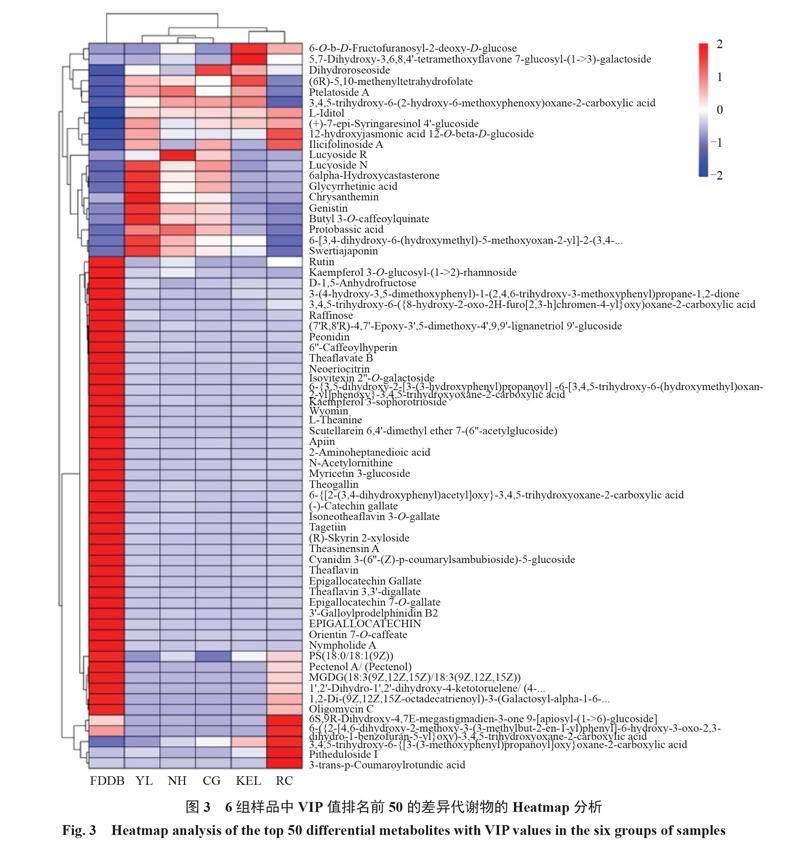
為了更直觀地展示樣本之間代謝物的表達差異,對所有顯著差異代謝物表達量進行層次聚類分析,根據VIP值對top 50差異代謝物表達量進行分析。篩選出的差異代謝物,大多數屬于脂類和類脂分子、生物堿及其衍生物、苯環型化合物、木酚素、新木酚素及其相關化合物、有機氧化合物和糖類多酮類化合物。結果如圖3所示,6種葉片的代謝物在表達豐度上具有明顯差異,福鼎大白茶葉片中含有的差異代謝物數目居多,但是在5個栽培種的梨葉片中同時也具有特異性的差異代謝物。相較于其他組代謝物質,在鴨梨葉片中,矢車菊苷(chrysanthemin)、三羥異黃酮(genistin)、獐牙菜紅素(swertiajaponin)、日當藥黃素(swertiajaponin)、6α-羥基蓖麻酮(6alpha-hydroxycastasterone )、甘草次酸(glycyrrhetinic acid)和原堿草酸(protobassic acid)等的含量相對較高;在南紅葉片中含量較高的差異代謝物主要有絲瓜皂苷R(lucyoside R)、3,4,5-三羥基-6-(2-羥基-6-甲氧基苯氧基)惡烷-2-羧酸[3,4,5-trihydroxy-6-(2-hydroxy-6-methoxyphenoxy)oxane-2-carboxylic acid]和原堿草酸(protobassic acid);在翠冠葉片中含量較高的差異代謝物主要有二氫薔薇苷(dihydroroseoside);在庫爾勒香梨葉片中含量較高的差異代謝物主要有6-O-b-D-呋喃果糖基-2-脫氧-D-葡萄糖(6-O-b-D-fructofuranosyl-2-deoxy-D-glucose)、5,7-二羥基-3,6,8,4'-四甲氧基黃酮 7-葡萄糖基-(1->3)-半乳糖苷[5,7-dihydroxy-3,6,8,4'-tetramethoxyflavone 7-glucosyl-(1->3)-galactoside]、二氫薔薇苷(dihydroroseoside)和(6R)-5,10-亞甲基四氫葉酸(6R)-5,10-methenyltetrahydrofolate等;在超紅葉片中含量較高的代謝物主要有3-反式-香豆酰圓環酸(3-trans-p-coumaroylrotundic acid)、三萜皂苷Ⅰ(pitheduloside Ⅰ)和6S,9R-二羥基-4,7E-大扁豆烯-3-酮9-[芹菜糖基-(1->6)-葡萄糖苷]{6S,9R-dihydroxy-4,7E-megastigmadien-3-one 9-[apiosyl-(1->6)-glucoside]})等(圖3)。以上結果表明,5個栽培種梨葉片中的代謝物存在顯著差異,也暗示著葉片的功能存在差異。
采用Pearson相關系數來衡量VIP值位于前50的差異代謝物含量的相關性。結果表明(圖4),1275對代謝物含量有642對(r>0)呈正相關,633對差異代謝物含量呈負相關(r<0)。與福鼎大白茶相比,5種梨葉片中排名位于前50的差異代謝物含量呈顯著正相關,如椴樹甙A(ptelatoside A)與(6R)-5,10-亞甲基四氫葉酸[(6R)-5,10-methenyltetrahydrofolate]、3-O-咖啡酰喹酸丁酯(butyl 3-O-caffeoylquinate)、原堿基酸(protobassic acid)、獐牙菜苷(swertiajaponin)、環西番蓮苷Ⅵ(cyclopassifloside Ⅵ)和葫蘆巴堿(trigonelline),其多數屬于脂類和類脂分子。相比其他組,甘草酸(glycyrrhetinic acid)是在鴨梨葉片中含量較高的代謝物,與其正相關代謝物主要有表沒食子兒茶素7-O-沒食子酸酯(epigallocatechin 7-O-gallate)、東方素7-O-咖啡酸酯(orientin 7-O-caffeate)、矢車菊苷(chrysanthemin)、三羥異黃酮(genistin)、槲皮素3-板藍根苷(quercetin 3-lathyroside)、3-O-咖啡酰喹酸丁酯(butyl 3-O-caffeoylquinate)、6α-羥基蓖麻酮(6alpha-hydroxycastasterone)、云杉苷(picein)、乙酸苯酚酯(phenyl acetate)和日當藥黃素(swertiajaponin),其正相關系數均大于0.9。相關性分析的結果表明,在葉片生長發育過程中,正相關的代謝物可能位于相同的代謝通路上,參與相同的發育過程。
2.4 差異代謝物的K-mean分析
采用K-mean對差異代謝物的表達趨勢進行聚類分析,將差異代謝物共劃分為12個亞組,在這12個組中,差異代謝物的變化趨勢基本一致,其中1、2、6、8、10和11組中差異代謝物的數目較多,分別是229、200、104、304、256和81個,其中1、2和8組中差異代謝物的含量變化趨勢基本一致。在3、6、7和9組中,福鼎大白茶葉片中代謝物的含量明顯高于梨葉片中代謝物的含量。在4組中總共含有29種代謝物,且其在梨葉片中的含量明顯高于在茶葉片中的含量,主要包括勒皮啶、吡咯黃質、艾格利諾,其分屬于苯環型化合物、糖類、多酮類化合物、脂類和類脂類分子。在5組中,超紅葉片中代謝物的含量顯著低于其他梨葉片中該代謝物的含量,主要代謝物有γ-西紅花酸(gamma-crocetin)、阿爾巴呋喃素A(albafuran A)、甲基三烯炔諾酮(gestrinone)、槲皮素3-(6''-丙二酰半乳糖苷)[quercetin 3-(6''-malonylgalactoside)]、1-十六烷基-2-[(2'-α-葡糖基)-β-葡糖基]-3-β-木糖基-sn-甘油{1-hexadecanyl-2-[(2'-alpha-glucosyl)-beta-glucosyl]-3-beta-xylosyl-sn-glycerol}、異肝素 2''-[6-(E)-阿魏酰葡糖苷] 4'-葡糖苷{isoscoparin 2''-[6-(E)-feruloylglucoside] 4'-glucoside},多數屬于脂類和類脂類分子,該結果也表明西方梨與東方梨葉片的代謝物含量具有明顯的差異(圖5)。
2.5 差異代謝物的代謝通路分析
對庫爾勒香梨、超紅、南紅、鴨梨、翠冠與福鼎大白茶葉片的差異代謝物進行KEGG富集分析,結果表明,5個比較組的差異代謝物共富集到78個KEGG通路上,每個比較組分別富集在76、71、73、70和74通路上。對5個不同的比較組分別分析發現,不同比較組的差異代謝物排名前3位的KEGG通路一致,分別是ABC轉運(pxb02010)、黃酮和黃酮醇生物合成(pxb00944)及類黃酮生物合成(pxb00941),分別有13~14、9~10和7~8種代謝物富集在這3個通路上。與福鼎大白茶相比,梨葉片中主要上調的代謝物有膽堿、D-核糖、D-甘露醇、山梨醇、木二糖和槲皮素3-(6''-丙二酰-葡萄糖苷);梨葉片中下調的差異代謝物含量主要有鄰苯二甲酸、阿洛糖、槲皮苷、異牧荊素、芹菜苷、異槲皮苷、蕓香苷、山柰酚、白藜蘆素、葉酮醇和表兒茶素。相較于茶樹葉片中的黃酮和黃酮醇生物合成途徑中代謝物的含量,梨葉片中檢測出該途徑所涉及VIP值排名靠前的差異代謝物含量呈現下調的趨勢,間接說明梨葉片中黃酮和黃酮醇次生代謝物的含量低于茶葉中該類代謝物的含量。梨中黃酮類物質主要存在于葉片和果皮中,在果樹應對非生物脅迫、調節花色、藥用、調控激素等方面具有重要作用。黃酮和黃酮醇生物合成途徑中包含的代謝物主要有槲皮苷、異牡荊素、芹菜苷、異槲皮苷、蘆丁、黃芪苷、漆葉苷、山柰酚-3-槐糖苷、槲皮素3-(6''-丙二酰基-糖苷)等,是植物體內重要的次生代謝物質,參與調節梨生長發育過程[20-21]。另外,VIP值排名前20的差異代謝物的富集通路基本一致,富集在每個通路上的差異代謝物的數目也基本一致,主要有甲硫腺苷、順-玉米素-O-葡萄糖苷、順-玉米素-7-N-葡萄糖苷、二氫玉米素-O-葡萄糖苷和二氫玉米素核苷。在單桿菌生物合成途徑中富集的代謝物主要有天門冬氨酸、精氨酸和四氫吡啶二羧酸(圖6)。
3 討 論
庫爾勒香梨、超紅、南紅、鴨梨、翠冠5個主栽品種代謝物數量和含量具有明顯差異,其中超紅梨葉片的代謝物明顯區別于其他4個梨品種,這可能是超紅梨屬于西方梨,而翠冠、鴨梨、南紅、庫爾勒香梨屬于東方梨所導致。另外,庫爾勒香梨葉片的代謝物在PCA聚類分析中與翠冠、鴨梨和南紅梨葉片代謝物聚類較遠,這也進一步說明起源不同的梨可能導致其代謝物差異。庫爾勒香梨屬于新疆梨系統,其是西方梨和東方梨雜交形成的品種[15]。因此,庫爾勒香梨葉片代謝物PCA聚類分析介于西方梨和東方梨之間。
葉片作為植物重要的營養器官,不僅支持果樹生長發育,也為果實品質發育提供營養成分。在超紅梨葉片代謝物的KEGG富集分析時發現,玉米素合成途徑(pxb00908)和單桿菌生物合成(pxb00261)途徑是超紅葉片代謝物顯著富集的兩個通路,該通路包含的代謝物主要有順-玉米素-O-葡萄糖苷、順-玉米素-7-N-葡萄糖苷和二氫玉米素-O-葡萄糖苷、天門冬氨酸、精氨酸和四氫吡啶二羧酸等,分屬于有機氧化合物和羧酸及其衍生物,主要參與植物抗性調控和果實品質前體物質的合成。梨葉片代謝物與果實品質的形成和調控具有相關性,代謝物的差異也可能是果實品質差異的原因之一。另外,果實中可溶性糖含量是評價果實品質和風味的重要因素,其主要包括葡萄糖、果糖、蔗糖、山梨醇、木糖和甘露醇等。果實中可溶性糖主要來源于葉片光合作用的產物,以蔗糖和山梨醇的形式存在于葉片中,經過韌皮部的長距離運輸最終到達果實,經過不同酶的催化反應,形成不同種類的糖,在梨中糖的運輸形式則以山梨醇為主[22-23]。葉片和果實中的糖代謝過程可按照產物的不同劃分為3種,即山梨醇代謝型、蔗糖代謝型和己糖代謝型。另外,果實獲取山梨醇的能力和對其轉化速率決定著果實的生長速度,調控果實的生長發育。山梨醇在山梨醇脫氫酶的催化下轉化成葡萄糖和果糖,在山梨醇氧化酶的作用下氧化成葡萄糖[24-25]。梨果實在生長發育的后期,葉片中山梨醇、果糖、甘露醇等代謝物的含量顯著上調是果實成熟時期合成和轉運糖分必要條件之一。此外,山梨醇還參與提高植物的抗逆性,促進養分吸收、提高愈傷誘導率、參與其他糖類物質的合成轉化等[26]。溫帶植物越冬期間,植物體內通過代謝糖醇(山梨醇、甘露醇等)、可溶性糖(蔗糖、海藻糖等)等冷凍保護劑的合成,來提高自身的耐寒性[27]。
福鼎大白茶葉片包含著豐富的營養成分與藥效成分,氨基酸、維生素、礦物質、多酚、咖啡堿和茶色素等均是茶葉中的重要組分,不僅可以滿足人們對營養物質的需求,同時具有抗氧化、抗炎、抗菌、抗癌及降血糖、降血壓、降血脂等功效[28-29]。梨葉片中也含有豐富的氨基酸類與黃酮類代謝物,具有抗菌消炎、抗氧化、防癌和抗癌的作用,如甘草次酸、三羥異黃酮、反式綠原酸等。筆者在本研究中也鑒定出具有開發利用價值的次生代謝物質,如在鴨梨葉片中的矢車菊苷、三羥異黃酮、獐牙菜紅素和日當藥黃素等;在南紅梨葉片中含量較高的絲瓜皂苷R、原堿草酸;在庫爾勒香梨葉片中含量較高的二氫薔薇苷;在超紅梨葉片中含量較高的3-反式-香豆酰圓環酸、三萜皂苷Ⅰ等。梨葉片中含有的可開發利用的次生代謝物質成分大多具有藥用價值,抗氧化性較強,在抗炎、抗菌等方面具有潛在的應用價值。這也間接增加了梨葉片的開發利用途徑,梨葉片可以作為活性物質的提取原料之一。
4 結 論
基于代謝組比較分析庫爾勒香梨、超紅、南紅、鴨梨、翠冠與對照福鼎大白茶的葉片代謝物含量和種類的差異,共檢測出9011種代謝物,代謝物種內聚類較為集中,種間聚類相對較遠。庫爾勒香梨、超紅、南紅、鴨梨、翠冠與福鼎大白茶比較組,差異代謝物的數目分別是1194、1153、1176、1153和1164種,共有的差異代謝物的數目為747種,特異性差異代謝物分別有55、155、54、59和28種。排名前50的差異代謝物在5個主栽梨葉片中的表達豐度上具有明顯差異,并且642對代謝物含量存在正相關,633對代謝物含量存在負相關。超紅葉片中代謝物在3、5和7組中含量差異較大,南紅梨在12組中代謝物含量相對較少。黃酮和黃酮醇生物合成(pxb00944)、ABC轉運(pxb02010)和類黃酮生物合成(pxb00941)等途徑是差異代謝物富集的主要通路,其中山梨醇、果糖、甘露醇、檸檬酸、水楊酸、馬來酸、甲基丙二酸等代謝物的含量顯著上調。玉米素合成(pxb00908)和單桿菌生物合成(pxb00261)途徑也是超紅葉片中代謝物顯著富集的代謝通路。因此,不同栽培種的梨葉片的代謝物在種類和含量上具有顯著差異,研究結果可為開發利用梨葉片資源提供理論基礎。
參考文獻 References:
[1] ZHAO J,DAVIS L C,VERPOORTE R. Elicitor signal transduction leading to production of plant secondary metabolites[J]. Biotechnology Advances,2005,23(4):283-333.
[2] 武予清,郭予元,楊艦. 棉株中抗蟲物質黃酮類化合物的高效液相色譜分析[J]. 植物保護,2000,26(5):1-3.
WU Yuqing,GUO Yuyuan,YANG Jian. Analysis of flavonoid substance in cotton plants for resistance to pests by HPLC[J]. Plant Protection,2000,26(5):1-3.
[3] 吳雅瓊,張春紅,楊海燕,閭連飛,李維林,吳文龍. 基于代謝組學分析黑莓、黑樹莓果實代謝物的差異[J]. 江蘇農業學報,2022,38(3):790-797.
WU Yaqiong,ZHANG Chunhong,YANG Haiyan,L? Lianfei,LI Weilin,WU Wenlong. Metabolomics analysis of differentially expressed metabolites in blackberry and black raspberry fruits[J]. Jiangsu Journal of Agricultural Sciences,2022,38(3):790-797.
[4] 張琴,董曉慶,林欣,彭俊森,羅登燦,黃世安,朱守亮. 基于UPLC-MS/MS的不同產地蜂糖李果實初生代謝差異分析[J]. 食品科學,2023,44(8):284-292.
ZHANG Qin,DONG Xiaoqing,LIN Xin,PENG Junsen,LUO Dengcan,HUANG Shian,ZHU Shouliang. Ultra-high performance liquid chromatography-tandem mass spectrometry-based metabolomic analysis of primary metabolites in ‘Fengtang plum fruits from different geographical origins[J]. Food Science,2023,44(8):284-292.
[5] 魯忠富,李艷偉,汪穎,吳曉花,吳新義,王尖,汪寶根,李國景. 基于廣泛靶向代謝組學的瓠瓜果實鮮味差異代謝物分析[J]. 中國蔬菜,2021(6):34-41.
LU Zhongfu,LI Yanwei,WANG Ying,WU Xiaohua,WU Xinyi,WANG Jian,WANG Baogen,LI Guojing. Analysis of fruit umami differential metabolites of bottle gourd based on widely targeted metabolomics[J]. China Vegetables,2021(6):34-41.
[6] 祝愛艷,梁露,侯金雪,孫雪楓,楊延峰,王遠興. 贛南臍橙代謝組學研究[J]. 中國食品學報,2020,20(3):276-281.
ZHU Aiyan,LIANG Lu,HOU Jinxue,SUN Xuefeng,YANG Yanfeng,WANG Yuanxing. Studies on metabonomics of Gannan navel orange[J]. Journal of Chinese Institute of Food Science and Technology,2020,20(3):276-281.
[7] 林欣,黃世安,張琴,彭俊森,羅登燦,朱守亮,陳紅,董曉慶. 采用UPLC-MS/MS分析低溫貯藏期間‘空心李果實初生代謝物[J]. 植物生理學報,2022,58(10):1982-1994.
LIN Xin,HUANG Shian,ZHANG Qin,PENG Junsen,LUO Dengcan,ZHU Shouliang,CHEN Hong,DONG Xiaoqing. Primary metabolites analysis of ‘Kongxin plum fruits during storage by UPLC-MS/MS[J]. Plant Physiology Journal,2022,58(10):1982-1994.
[8] 趙燕妮,張坤,許牡丹,雷靖,劉歡,劉寧,陳雪峰,張迪. 基于GC-MS代謝組學技術的不同品種獼猴桃果實化學成分差異性研究[J]. 陜西科技大學學報,2022,40(1):45-50.
ZHAO Yanni,ZHANG Kun,XU Mudan,LEI Jing,LIU Huan,LIU Ning,CHEN Xuefeng,ZHANG Di. Study on the chemical constituents of different kiwifruit cultivars based on GC-MS metabolomics[J]. Journal of Shaanxi University of Science & Technology,2022,40(1):45-50.
[9] 潘銅華. CO2富集與光強互作對番茄光合碳同化的影響及代謝組研究[D]. 楊凌:西北農林科技大學,2019.
PAN Tonghua. Interactive effects of elevated CO2 and light intensity on photosynthetic carbon assimilation and metabolomics of tomato[D]. Yangling:Northwest A & F University,2019.
[10] 唐晨晨,張文霞,陳芳珍,武志健,黃科,王軍偉. 代謝組和轉錄組聯合解析青花菜芽苗黃酮類物質對外源ABA的響應機制[J]. 中國瓜菜,2024,37(3):35-44.
TANG Chenchen,ZHANG Wenxia,CHEN Fangzhen,WU Zhijian,HUANG Ke,WANG Junwei. The metabolome and transcriptome jointly resolve the response mechanism of flavonoids in broccoli sprouts to exogenous ABA[J]. China Cucurbits and Vegetables,2024,37(3):35-44.
[11] 劉芹,黃保,胡素娟,牛森園,吳杰,周奧碩,孔維麗. 基于非靶向代謝組學的平菇子實體發育過程中菌絲體差異代謝物分析[J]. 中國瓜菜,2024,37(1):45-55.
LIU Qin,HUANG Bao,HU Sujuan,NIU Senyuan,WU Jie,ZHOU Aoshuo,KONG Weili. Analysis of differential metabolites during fruiting body development of Pleurotus ostreatus based on untargeted metabolomics[J]. China Cucurbits and Vegetables,2024,37(1):45-55.
[12] LI H Y,LV Q Y,LIU A K,WANG J R,SUN X Q,DENG J,CHEN Q F,WU Q. Comparative metabolomics study of Tartary [Fagopyrum tataricum (L.) Gaertn] and common (Fagopyrum esculentum Moench) buckwheat seeds[J]. Food Chemistry,2022,371:131125.
[13] 李川,喬江方,黃璐,張美微,張盼盼,牛軍,劉京寶. 轉錄組及代謝組聯合解析玉米響應花粒期高溫脅迫機制[J]. 華北農學報,2020,35(1):8-21.
LI Chuan,QIAO Jiangfang,HUANG Lu,ZHANG Meiwei,ZHANG Panpan,NIU Jun,LIU Jingbao. Transcriptome and metabolome analysis to reveal the mechanisms responding to high temperature stress in anthesis stage of maize[J]. Acta Agriculturae Boreali-Sinica,2020,35(1):8-21.
[14] 郭家鑫,魯曉宇,陶一凡,郭慧娟,閔偉. 棉花在鹽堿脅迫下代謝產物及通路的分析[J]. 作物學報,2022,48(8):2100-2114.
GUO Jiaxin,LU Xiaoyu,TAO Yifan,GUO Huijuan,MIN Wei. Analysis of metabolites and pathways in cotton under salt and alkali stresses[J]. Acta Agronomica Sinica,2022,48(8):2100-2114.
[15] WU J,WANG Y T,XU J B,KORBAN S S,FEI Z J,TAO S T,MING R,TAI S S,KHAN A M,POSTMAN J D,GU C,YIN H,ZHENG D M,QI K J,LI Y,WANG R Z,DENG C H,KUMAR S,CHAGN? D,LI X L,WU J Y,HUANG X S,ZHANG H P,XIE Z H,LI X,ZHANG M Y,LI Y H,YUE Z,FANG X D,LI J M,LI L T,JIN C,QIN M F,ZHANG J Y,WU X,KE Y Q,WANG J,YANG H,ZHANG S L. Diversification and independent domestication of Asian and European pears[J]. Genome Biology,2018,19:77.
[16] 王宏,楊王莉,藺經,楊青松,李曉剛,盛寶龍,常有宏. 早熟砂梨‘蘇翠1號與其親本‘翠冠‘華酥成熟果實差異代謝產物及差異基因比較分析[J]. 園藝學報,2022,49(3):493-508.
WANG Hong,YANG Wangli,LIN Jing,YANG Qingsong,LI Xiaogang,SHENG Baolong,CHANG Youhong. Comparative metabolic and transcriptomic analysis of ripening fruit in pear cultivars of ‘Sucui 1 ‘Cuiguan and ‘Huasu[J]. Acta Horticulturae Sinica,2022,49(3):493-508.
[17] 趙芫. 乙烯利協同茉莉酸甲酯調控‘紅早酥梨果皮著色的代謝組學研究[D]. 杭州:浙江大學,2019.
ZHAO Yuan. Metabonomics study on ethephon and methyl jasmonate synergistically regulating peel coloration of ‘Hongzaosu pear[D]. Hangzhou:Zhejiang University,2019.
[18] 解敏,黃瑛,王馨,王秀梅,唐娟娟,方從兵. 基于GC-MS技術測定碭山酥梨果實主要品質特征成分的代謝物譜[J]. 安徽農業大學學報,2012,39(5):692-696.
XIE Min,HUANG Ying,WANG Xin,WANG Xiumei,TANG Juanjuan,FANG Congbing. Metabolomic analysis on the major quality components in pear (Pyrus bretschneideri Rehd. cv. Dangshansuli) fruits by gas chromatography-mass spectrometry[J]. Journal of Anhui Agricultural University,2012,39(5):692-696.
[19] 高靜怡,馬浩,劉東賀,方嘯宇,劉曉. 非靶向代謝組學對砂梨水心病不同代謝物質的差異分析[J]. 分子植物育種,2021,19(24):8297-8304.
GAO Jingyi,MA Hao,LIU Donghe,FANG Xiaoyu,LIU Xiao. The difference analysis of non targeted metabonomics on different metabolites in watercore Pyrus pyrifolia[J]. Molecular Plant Breeding,2021,19(24):8297-8304.
[20] 曹運琳,邢夢云,徐昌杰,李鮮. 植物黃酮醇生物合成及其調控研究進展[J]. 園藝學報,2018,45(1):177-192.
CAO Yunlin,XING Mengyun,XU Changjie,LI Xian. Biosynthesis of flavonol and its regulation in plants[J]. Acta Horticulturae Sinica,2018,45(1):177-192.
[21] 張小雙,鄭迎春,曹玉芬,田路明,董星光,張瑩,齊丹,霍宏亮. ‘早酥和‘南果梨16個部位多酚物質組成及含量分析[J]. 中國農業科學,2017,50(3):545-555.
ZHANG Xiaoshuang,ZHENG Yingchun,CAO Yufen,TIAN Luming,DONG Xingguang,ZHANG Ying,QI Dan,HUO Hongliang. The composition and content of polyphenols in 16 parts of ‘Zaosu and ‘Nanguoli[J]. Scientia Agricultura Sinica,2017,50(3):545-555.
[22] 張虎平. 梨果實內糖的轉運及積累特性研究[D]. 南京:南京農業大學,2011.
ZHANG Huping. Study on characteristic of transport and accumulation of sugar in developing Pyrus ssp. fruit[D]. Nanjing:Nanjing Agricultural University,2011.
[23] LIU D F,NI J B,WU R Y,TENG Y W. High temperature alters sorbitol metabolism in pyrus pyrifolia leaves and fruit flesh during late stages of fruit enlargement[J]. Journal of the American Society for Horticultural Science,2013,138(6):443-451.
[24] 李甲明. 梨糖轉運蛋白基因家族分析及PbtMT4和PbSWEET基因功能研究[D]. 南京:南京農業大學,2017.
LI Jiaming. Identification of pear sugar transporter genes and PbtMT4 and PbSWEET genes function analysis[D]. Nanjing:Nanjing Agricultural University,2017.
[25] OURA Y,YAMADA K,SHIRATAKE K,YAMAKI S. Purification and characterization of a NAD+-dependent sorbitol dehydrogenase from Japanese pear fruit[J]. Phytochemistry,2000,54(6):567-572.
[26] HOEKSTRA F A,GOLOVINA E A,BUITINK J. Mechanisms of plant desiccation tolerance[J]. Trends in Plant Science,2001,6(9):431-438.
[27] JANSK? A,MARS?K P,ZELENKOV? S,OVESN? J. Cold stress and acclimation - what is important for metabolic adjustment?[J]. Plant Biology,2010,12(3):395-405.
[28] 戴偉東,解東超,林智. 白茶功能性成分及保健功效研究進展[J]. 中國茶葉,2021,43(4):1-8.
DAI Weidong,XIE Dongchao,LIN Zhi. Research progress of white teas functional ingredients and health benefits[J]. China Tea,2021,43(4):1-8.
[29] 黃彪,劉文靜,吳建鴻,吳建衍. 福鼎大白茶樹鮮葉不同茶制品活性成分比較[J]. 食品安全質量檢測學報,2021,12(8):3219-3223.
HUANG Biao,LIU Wenjing,WU Jianhong,WU Jianyan. Comparison of active components of different tea products from fresh leaves of Camellia sinensis Fuding Dabai[J]. Journal of Food Safety & Quality,2021,12(8):3219-3223.
收稿日期:2024-02-02 接受日期:2024-04-20
基金項目:山東省農科院創新工程項目(CXGC2023F21、CXGC2023A13);山東省果樹研究所青年基金(GSS2022QN11);國家梨產業技術體系(CARS-28-37);山東省自然科學基金(ZR2020MC141、ZR2021MC177、ZR2023MC061);農業農村部園藝作物種質資源利用重點實驗室開放基金(NYZS202206)
作者簡介:焦慧君,女,助理研究員,博士,研究方向為果實品質調控。Tel:0538-8806302,E-mail:jiaohj_njau@163.com。#為共同第一作者。冉昆,男,研究員,博士,研究方向為果樹遺傳育種與栽培管理。Tel:0538-8207123,E-mail:rkrl001@126.com
*通信作者 Author for correspondence. Tel:0538-8207123,E-mail:weisw2007@163.com
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
