社區獲得性肺炎免疫相關基因的鑒定與分析
2024-12-15 00:00:00董海麗王國勝谷雙龍宋南萍
上海醫藥 2024年21期
摘 要 目的:社區獲得性肺炎(CAP)是兒童住院和死亡的主要原因,本研究旨在從CAP中篩選免疫相關的基因。方法:從GEO數據庫獲取CAP數據,利用生物信息學方法篩選免疫相關基因,并進一步探究這些基因的功能及其與免疫細胞的關系。結果:CAP疾病組與正常組之間的差異基因主要與抗原加工和遞呈、核糖體、細胞凋亡和炎癥反應等功能相關。WGCNA和PPI分析共確定了10個免疫相關的關鍵基因:IL4R、FGR、CCR3、CCR1、ARG1、CEBPB、IL17RA、NFKBIA、PPARG和NCF4。這些基因的表達水平與免疫細胞的浸潤水平呈顯著的相關性。結論:本研究篩選出了可能在CAP免疫反應中起重要作用的基因。
關鍵詞 社區獲得性肺炎 免疫 兒童 呼吸道感染
中圖分類號:R563 文獻標志碼:A 文章編號:1006-1533(2024)21-0053-07
引用本文 董海麗, 王國勝, 谷雙龍, 等. 社區獲得性肺炎免疫相關基因的鑒定與分析[J]. 上海醫藥, 2024, 45(21): 53-59; 75.
Identification and analysis for immune-related genes in community-acquired pneumonia
DONG Haili, WANG Guosheng, GU Shuanglong, SONG Nanping
(Department of Pediatrics, Zhengzhou 460 Hospital, Zhengzhou 450000, China)
ABSTRACT Objectives: Community-acquired pneumonia (CAP) is a leading cause of hospitalization and death in children. The present study mainly aimed to screen out immune-related genes in CAP. Methods: CAP data were retrieved from GEO online database. The immune-related genes were identified using bioinformatics method, and their function and relationship with immune cells were further investigated. Results: The differential expressed genes between CAP group and normal group were mainly related to antigen processing and presentation, ribosome, apoptosis and inflammatory response. Moreover, a total of 10 key immune-related genes were selected using WGCNA and PPI analysis, including IL4R, FGR, CCR3, CCR1, ARG1, CEBPB, IL17RA, NFKBIA, PPARG, and NCF4. The expression levels of these genes were significantly correlated with the infiltration level of immune cells. Conclusion: The key genes identified from this study might play important roles in the immune response of CAP.
KEY WORDS community-acquired pneumonia; immune; children; respiratory tract infection
肺炎是一種常見的急性呼吸道感染,按照患者感染場所,大致分為社區獲得性肺炎(community-acquired pneumonia,CAP)和醫院獲得性肺炎。多種微生物可引起肺炎,包括細菌、呼吸道病毒和真菌[1]。其中,CAP是兒童住院和死亡的主要原因[2]。但是,該疾病的診斷和治療均存在一定的障礙。首先,CAP沒有特定的癥狀,其診斷一般依賴微生物學診斷或者胸部X射線檢查。但前者取樣困難,后者可靠性不夠[3-4]。其次,其治療通常建議使用抗生素,但CAP可能是細菌感染,也可能是細菌-病毒共感染,使得其治療效果非常有限[4]。故而,探尋CAP特異性標志物是當前工作的重中之重。CAP的發展在很大程度上取決于宿主的免疫反應,多種免疫細胞在其中發揮重要作用[5]。在本研究中,我們利用生物信息學分析,通過對來自公共數據庫中的數據進行分析,在CAP中篩選與免疫反應相關的基因,以期為CAP的診斷與治療提供新的思路。
1 資料與方法
1.1 數據來源
從NCBI GEO數據庫(Gene Expression Omnibus,GEO)中下載GES103119數據集的數據,該數據集包含152例CAP樣本和20例對照樣本。樣本類型為血液,測序平臺為GPL10558 Illumina Human HT-12 V4.0 expression bead chip。研究思路如圖1所示。
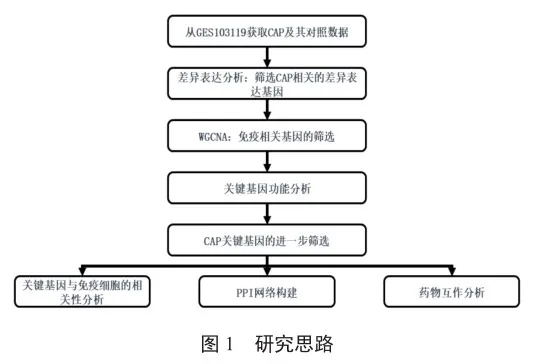
1.2 CAP相關差異基因篩選
利用R包limma,基于CAP基因表達數據以及樣本分組數據,對所有基因進行差異表達分析,篩選患病樣本與正常樣本之間顯著差異表達的基因,閾值設置為P<0.05和|log2FC|>0.585。
1.3 CAP疾病組和對照組之間功能通路及免疫細胞浸潤差異分析
利用基因集富集分析(gene set enrichment analysis,GSEA)算法識別患病組與正常組之間的差異調節通路。基于基因表達數據和樣本分組,進行生物加工(biology process,BP)、細胞組分(cellular component,CC)、分子功能(molecular function,MF)和京都基因與基因組百科全書(Kyoto encyclopedia of genes and genomes,KEGGK)通路富集分析,顯著富集閾值設定為|NES|>1和FDR<0.25。
基于CAP表達數據,采用免疫浸潤分析工具CIBERSORT進行22種免疫細胞構成比例的計算,比較患病組和正常組之間免疫細胞的浸潤程度差異。
1.4 CAP免疫相關關鍵基因篩選
加權基因共表達網絡分析(weighted gene co- expression network analysis,WGCNA)是一種用于描述不同樣品之間基因關聯模式的系統生物學方法。基于CAP的數據集和免疫微環境分析得到的免疫浸潤數據,首先篩選變異較大(MAD排名TOP 20%)的基因,然后利用R包WGCNA對篩選后的基因進行分析,鑒定出與免疫細胞具有高度協同變化的基因集模塊。然后將篩選出的模塊基因與差異表達基因取交集,得到CAP免疫關鍵基因。
1.5 關鍵基因富集分析
針對CAP免疫關鍵基因進行富集分析,使用R包clusterProfiler對這些關鍵基因進行BP、CC、MF和KEGG通路富集分析,并將P<0.05視為顯著性閾值,篩選出顯著性富集結果。
1.6 CAP 關鍵基因的進一步篩選
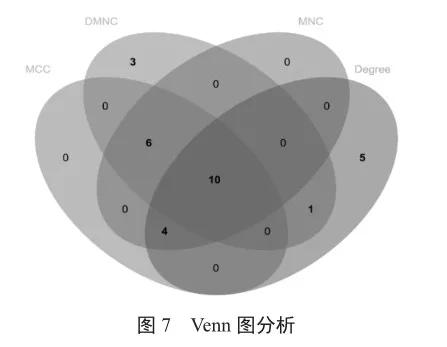
基于CAP關鍵基因,利用STRING數據庫中的人類蛋白-蛋白相互作用關系數據,構建CAP免疫關鍵基因的蛋白相互作用關系(PPI)。然后利用Cytoscape構建蛋白互作網絡。使用Cytoscape中的CytoHubba插件通過4種拓撲網絡算法最大集團中心度(maximal clique centrality,MCC)、最大鄰域分量密度(density of maximum neighborhood component,DMNC)、最大鄰域分量(maximum neighborhood component,MNC)和Degree分別給這個PPI結構中包含的基因進行打分并排序,并篩選TOP 20的基因,選擇共有基因作為最終的hub基因。
1.7 hub基因與免疫細胞的相關性分析
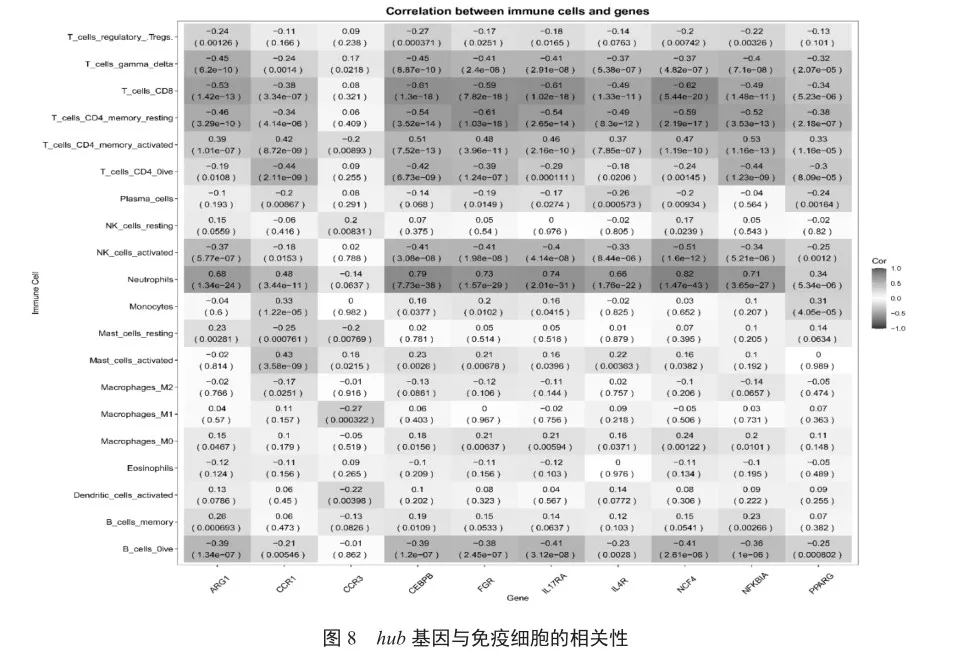
結合hub基因表達矩陣和CAP免疫浸潤數據進行相關性分析,將hub基因與所有免疫細胞進行相關性分析,最后挑選出免疫細胞相關性較高的基因。
1.8 hub基因的PPI構建
利用GeneMANIA在線數據庫對hub基因及其相互作用基因進行PPI分析,來預測基因共定位、共享蛋白質結構域、共表達、預測以及通路之間的相關性。
1.9 hub基因藥物相互作用分析
利用DGIdb基因藥物關系數據庫對hub基因進行基因藥物靶點預測,利用Cytoscape對其進行可視化,構建藥物與基因之間的相互作用網絡。
2 結果
2.1 CAP疾病相關差異基因篩選
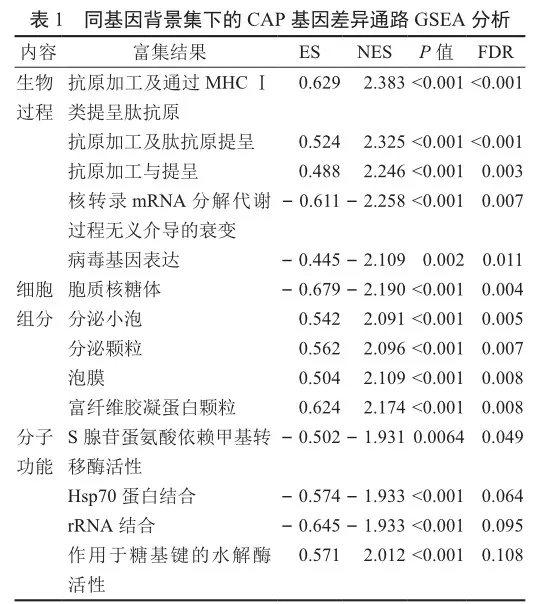
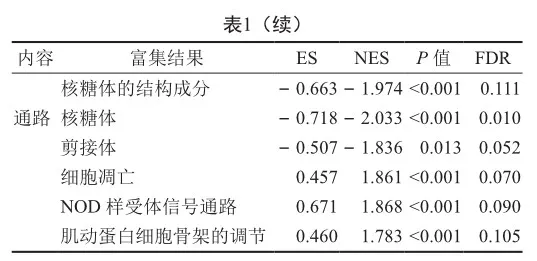
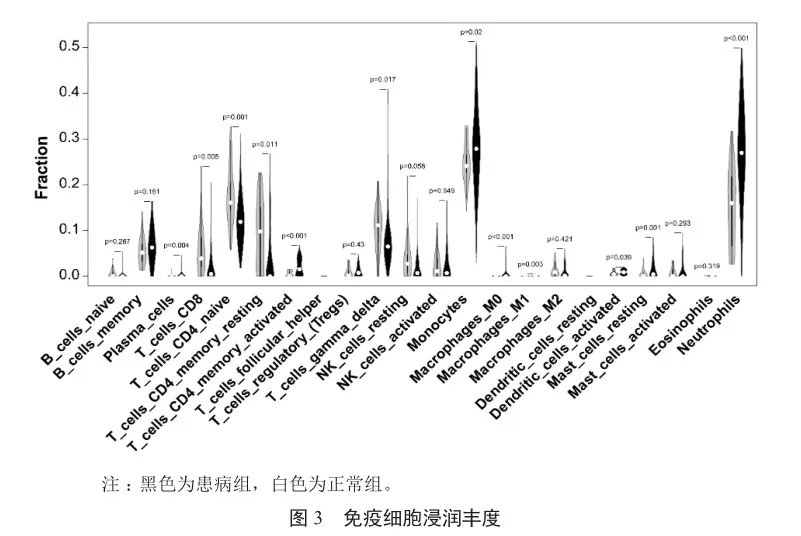
與對照組相比,CAP疾病組共篩選得到284個上調基因,239個下調基因(圖2)。GSEA分析表明CAP相關基因主要參與抗原加工和遞呈、核糖體、細胞凋亡和炎癥等相關功能或通路(表1)。此外,CAP疾病組和正常組免疫細胞浸潤程度有顯著差異,如CD4記憶激活T細胞、單核細胞和中性粒細胞在疾病組浸潤水平顯著高于正常組,而CD8 T細胞、幼稚CD4 T細胞、靜息CD4 T細胞在疾病組浸潤水平顯著低于正常組(圖3)。

2.2 WGCNA篩選免疫相關基因
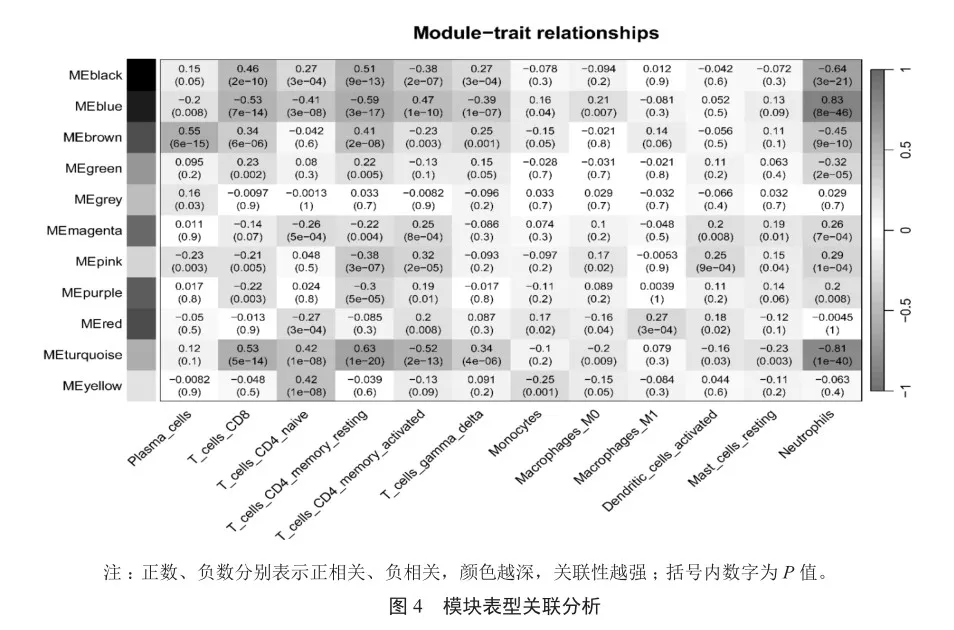
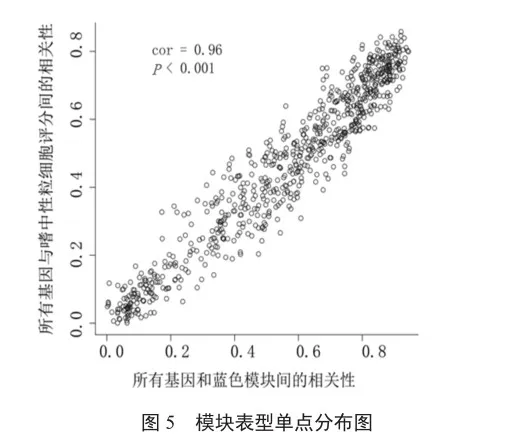
12種免疫細胞的豐度在不同分組中呈顯著性差異(P<0.05),以這12種免疫細胞作為表型,將CAP表達數據中的基因用于構建WGCNA網絡,篩選出與差異免疫細胞相關的關鍵模塊及基因。軟閾值為4時WGCNA網絡接近無尺度分布;結合混合動態剪切樹算法共得到11個模塊。結合差異免疫細胞表型數據,將模塊與表型進行關聯分析,結果表明,這11個模塊與免疫細胞之間具有顯著的正或負相關關系(圖4)。在這11個模塊中,MEblue模塊(764個基因)與中性粒細胞呈最強顯著正相關關系(圖4和圖5,r=0.83,P<0.05)。因此選用藍色模塊中的基因作為關鍵模塊基因,將這些關鍵模塊基因與差異表達分析所得的差異基因進行比對Venn圖分析,得到235個共有的免疫相關基因。
2.3 免疫相關基因功能富集分析
對上一步得到的235個免疫相關基因進行功能和通路富集分析,得到顯著的499個生物過程、40個細胞組分、 67個分子功能和17個通路,各選取最顯著的10個功能或通路進行展示(圖6)。結果顯示,這些免疫相關基因與免疫反應相關。
2.4 hub基因鑒定
PPI網絡構建得到一個共包含99個基因的網絡結構。然后,使用Cytoscape中的CytoHubba插件通過4種拓撲網絡算法MCC、DMNC、MNC和Degree分別給這個PPI結構中包含的基因進行打分并排序,并篩選TOP 20的基因,Venn分析中得到的10個基因IL4R、FGR、CCR3、CCR1、ARG1、CEBPB、IL17RA、NFKBIA、PPARG和NCF4作為hub基因(圖7)。這10個基因與免疫細胞之間也具有顯著的相關性(圖8)
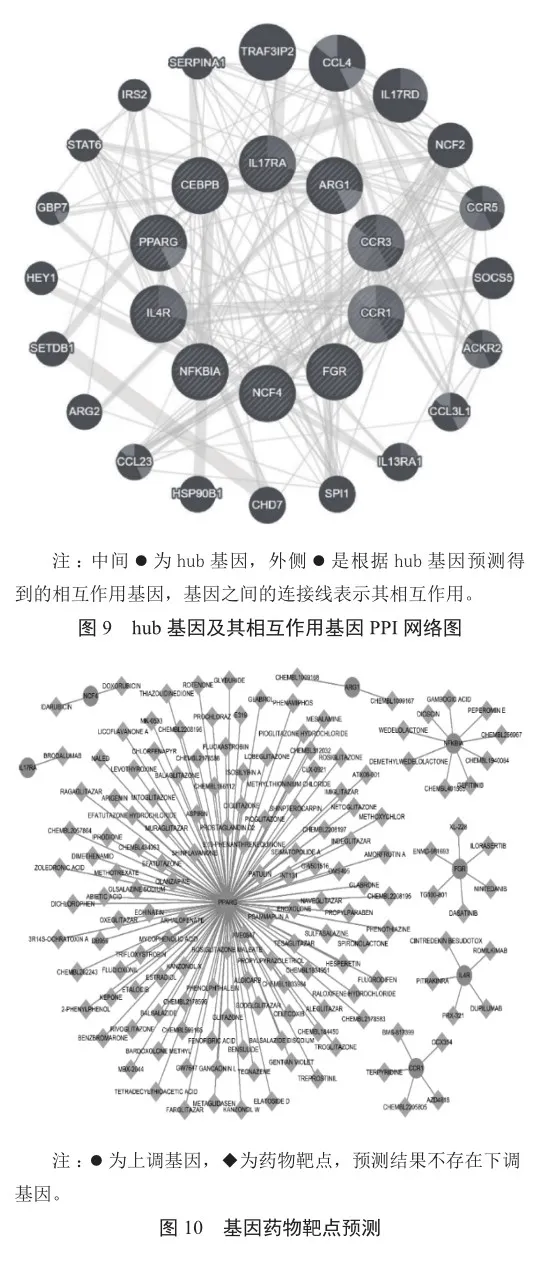
2.5 hub基因的PPI構建以及基因藥物相互分析

此外,hub基因本身之間也具有非常緊密的聯系(圖9)。對上述hub基因進行藥物靶點預測,并探究這些基因可能與哪些藥物互相作用。預測結果顯示這些基因在疾病中都是上調的。其中,IL4R存在5個藥物靶點,NCF4存在2個藥物靶點,FGR存在6個藥物靶點,CCR1存在5個藥物靶點,ARG1存在2個藥物靶點,IL17RA存在1個藥物靶點,NFKBIA存在9個藥物靶點,PPARG存在121個藥物靶點(圖10)。
3 討論
本研究通過一系列生物信息學分析手段如WGCNA、 PPI等,共篩選得到10個免疫相關的hub基因:IL4R、FGR、CCR3、CCR1、ARG1、CEBPB、IL17RA、NFKBIA、PPARG和NCF4。這些基因的表達水平與免疫細胞的浸潤水平具有顯著的相關性。
細菌、病毒等感染均可引發CAP,其發展在很大程度上取決于宿主的免疫反應。本研究鑒定得到的10個hub基因在以往研究中均有報道,且均與免疫反應相關。其中,IL4R是IL4和IL13的常見受體,在IL4和IL13介導的炎癥反應中起關鍵作用,與IL4結合可調節過敏性炎癥中的免疫反應信號[6]。CCR3屬于G蛋白偶聯受體,在嗜酸性粒細胞、嗜堿性粒細胞、2型T輔助細胞等相關炎性細胞中均有表達,通常被認為是人類免疫缺陷病毒感染和過敏性疾病的治療靶點[7]。CCR1是巨噬細胞炎癥蛋白1α(MIP-1α)等趨化因子的膜受體[8],在保護宿主免受炎癥反應以及對病毒和寄生蟲的易感性方面發揮作用[9]。ARG1(精氨酸酶-1)在肺炎中具有炎癥調節作用,小鼠骨髓細胞中Arg1基因的缺失會導致嚴重的中性粒細胞主導的炎性肺損傷[10]。Arg1敲除的小鼠呼吸道和淋巴結中中性粒細胞、巨噬細胞和淋巴細胞浸潤水平均較高,淋巴結、肺間質和氣道中T細胞活化、IFNγ和IL17的產生增加[10]。CEBPB與多種疾病的免疫反應相關[11]。IL17RA是由IL17A編碼的跨膜蛋白,與IL17A親和力較低,但在許多炎癥及自身免疫性疾病中起致病作用[12]。PPARG是一種配體依賴性轉錄因子,參與代謝、免疫應答和發育的多方面,兩個Pparg等位基因的全身性缺失會引起小鼠全身脂肪萎縮以及嚴重的2型糖尿病[13]。NCF4是煙酰胺二核苷酸磷酸氧化酶復合物的組成部分,是先天免疫反應的關鍵因子[14],NCF4的缺失會導致活性氧減少,小鼠表現為中性粒細胞凋亡,機體免疫反應加重[15]。以上研究表明,本研究中篩選出來的免疫相關基因或多或少均與CAP進程或免疫反應相關,但這些基因的具體作用及相關機制需要在進一步的研究中進行完善。此外,本研究還表明,這些關鍵的基因可能與一些藥物作用相關,故而在進一步的研究中有必要探討這些基因與藥物的關系。
參考文獻
[1] Torres A, Cilloniz C, Niederman MS, et al. Pneumonia[J]. Nat Rev Dis Primers, 2021, 7(1): 25.
[2] Kronman MP, Hersh AL, Feng R, et al. Ambulatory visit rates and antibiotic prescribing for children with pneumonia, 1994-2007[J]. Pediatrics, 2011, 127(3): 411-418.
[3] Eklundh A, Rhedin S, Ryd-Rinder M, et al. Etiology of clinical community-acquired pneumonia in Swedish children aged 1-59 months with high pneumococcal vaccine coveragethe TREND study[J]. Vaccines (Basel), 2021, 9(4): 384.
[4] Pernica JM, Kam AJ, Eltorki M, et al. Novel care pathway to optimise antimicrobial prescribing for uncomplicated community-acquired pneumonia: study protocol for a prospective before-after cohort study in the emergency department of a tertiary care Canadian children’s hospital[J]. BMJ Open, 2022, 12(11): e062360.
[5] Gao M, Wang K, Yang M, et al. Transcriptome analysis of bronchoalveolar lavage fluid from children with mycoplasma pneumoniae pneumonia reveals natural killer and T cellproliferation responses[J]. Front Immunol, 2018, 9: 1403.
[6] Junttila IS. Tuning the cytokine responses: an update on interleukin (IL)-4 and IL-13 receptor complexes[J]. Front Immunol, 2018, 9: 888.
[7] 唐思藝, 朱新華. CCR3基因在呼吸道過敏性疾病中對相關炎性細胞的作用研究[J]. 臨床耳鼻咽喉頭頸外科雜志, 2021, 35(1): 80-84.
[8] Nakano H, Kirino Y, Takeno M, et al. GWAS-identified CCR1 and IL10 loci contribute to M1 macrophage-predominant inflammation in Beh?et’s disease[J]. Arthritis Res Ther, 2018, 20(1): 124.
[9] Kholodnyuk I, Rudevica Z, Leonciks A, et al. Expression of the chemokine receptors CCR1 and CCR2B is up-regulated in peripheral blood B cells upon EBV infection and in established lymphoblastoid cell lines[J]. Virology, 2017, 512: 1-7.
[10] Haydar D, Gonzalez R, Garvy BA, et al. Myeloid arginase-1 controls excessive inflammation and modulates T cell responses in Pseudomonas aeruginosa pneumonia[J]. Immunobiology, 2021, 226(1): 152034.
[11] Selvaraj G, Kaliamurthi S, Peslherbe GH, et al. Identifying potential drug targets and candidate drugs for COVID-19: biological networks and structural modeling approaches[J]. F1000Res, 2021, 10: 127.
[12] Yanagisawa H, Hashimoto M, Minagawa S, et al. Role of IL-17A in murine models of COPD airway disease[J]. Am J Physiol Lung Cell Mol Physiol, 2017, 312(1): L122-L130.
[13] Toffoli B, Gilardi F, Winkler C, et al. Nephropathy in Pparg1 mice highlights PPARγ systemic activities in metabolism and in the immune system[J]. PLoS One, 2017, 12(2): e0171474.
[14] Ju Z, Wang C, Wang X, et al. Role of an SNP in alternative splicing of bovine NCF4 and mastitis susceptibility[J]. PLoS One, 2015, 10(11): e0143705.
[15] Winter S, Hultqvist Hopkins M, Laulund F, et al. A reduction in intracellular reactive oxygen species due to a mutation in NCF4 promotes autoimmune arthritis in mice[J]. Antioxid Redox Signal, 2016, 25(18): 983-996.
猜你喜歡
少兒美術·書法版(2021年11期)2021-10-20 06:23:28
少兒美術·書法版(2021年8期)2021-10-20 06:08:10
中國實用醫藥(2016年28期)2016-12-07 07:29:40
中國實用醫藥(2016年19期)2016-08-05 21:48:50
雜文選刊(2016年7期)2016-08-02 08:39:56
中國實用醫藥(2016年16期)2016-07-26 21:29:20
中國實用醫藥(2016年14期)2016-05-20 04:03:00
小天使·一年級語數英綜合(2016年6期)2016-05-14 12:21:05
中國實用醫藥(2016年12期)2016-05-04 09:22:06
中國實用醫藥(2016年6期)2016-03-17 11:12:31
