現代舞訓練與弦樂訓練對腦灰質體積的差異影響*
2018-07-07 05:53:04李谷靜堯德中
心理科學進展 2018年7期
李谷靜 李 薪 賀 輝 羅 程 堯德中
(電子科技大學生命科學與技術學院, 成都 610054)
1 引言
大腦的可塑性受多種因素影響, 其中長期學習訓練是其結構改變的重要因素(Draganski et al.,2004)。舞蹈或音樂訓練是指依據科學原理, 計劃性地對受訓者進行機體結構機能重建和技術加工,使其通過長期練習掌握常人不易獲得的特殊技能的過程(覃嬪, 2018; 馬清, 2000)。兩類訓練的特殊性與長期性為研究腦可塑性提供了良好模型。舞蹈訓練與音樂訓練具有許多異同點:兩者都是復雜感覺運動過程, 涉及大腦對多種感知覺信息的加工整合; 都是情感表達性藝術訓練, 需要腦對內部軀體變化與外部刺激信息進行交互加工, 實現藝術情感的表現。不同在于舞蹈訓練是以舞者自身為工具的韻律性動覺訓練, 它強調本體感覺與身體控制, 強調視?聽?動的高度整合。舞蹈學習往往通過大量的動作觀察與動作模仿而實現(平心, 2004)。音樂訓練特別是器樂訓練, 是以掌握特殊演奏技能為目標的訓練, 它強調聽覺與手指精細運動的高度協調與快速反饋, 要求聽?動的高度整合。音樂學習通過聽覺信息修正動作,從而達到對演奏技能的修正(Mutschler et al., 2007;蔣存梅, 2016)。
目前, 相關研究主要從腦功能與腦結構的改變來刻畫兩種訓練的可塑性影響。關于舞蹈的研究主要聚焦于動作觀察網絡(action observation network, AON)以及感覺運動網絡。研究AON主要是為探索舞蹈專業性對動作知覺以及對目標導向性行為的影響。這些研究發現個體通過舞蹈訓練所獲得的真實動作經驗, 與動作觀察所獲得的間接經驗都會影響其動作觀察網絡, 具體來講兩種動作經驗都會增進該網絡的激活(Calvo-Merino,Glaser, Grèzes, Passingham, & Haggard, 2005; Cross,Hamilton, & Grafton, 2006; Jola et al., 2013)。與此同時, 研究者們通過掃描舞者執行下肢舞蹈動作時的腦功能活動情況, 進而研究舞蹈動作中獨特的感覺運動過程。其結果顯示舞蹈動作執行時與本體感覺、動作控制、節奏保持等功能相關的皮層、皮層下結構及小腦等廣泛區域被激活(Brown,Martinez, & Parsons, 2006; Ono et al., 2014)。關于腦結構方面的研究也表明長期舞蹈訓練將導致感覺運動區域的灰質與白質結構發生廣泛改變。H?nggi, Koeneke, Bezzola 和 J?ncke (2010)首次開展芭蕾舞蹈家的大腦結構研究。他們發現舞蹈家的左側前運動皮層、輔助運動皮層、殼核與額上回的灰質體積顯著降低; 雙側皮質脊髓束、雙側內囊與胼胝體的白質體積顯著降低; 連接前運動皮層的白質纖維微結構出現各向異性降低。舞蹈家大腦的廣泛結構改變引起了其他研究者的關注。Giacosa, Karpati, Foster, Penhune和 Hyde(2016)通過進一步對比舞蹈家與音樂家的白質纖維微結構, 發現舞蹈家感覺運動網絡內的白質纖維結構(如皮質脊髓束、上縱束和胼胝體)變化明顯。較之音樂家與對照組, 舞蹈家的白質纖結構更為發散和長程, 連接了更為廣泛的感覺運動皮層。綜上所述, 舞蹈訓練對腦結構的特異性影響主要表現在涉及感覺運動的廣泛腦區發生改變,具體表現為功能皮層灰質體積的減小, 且連接這些皮層的白質纖維微結構發生改變(各項異性降低)。關于音樂的研究主要集中于聽?動整合相關網絡以及音樂認知的神經過程。研究發現音樂家的感覺運動皮層、聽覺皮層及小腦的功能增強,說明音樂訓練可能提升了大腦對聽?動信息的處理與整合(Baumann et al., 2007; Bangert et al.,2006)。另一類研究則揭示音樂訓練中的句法加工、工作記憶和音樂情緒加工的神經過程, 結果發現音樂訓練增強了額下回、頂下小葉、腦島等相關腦區的功能(Groussard et al., 2010; Koelsch &Siebel, 2005)。關于音樂家腦結構的研究結果基本與其腦功能的特異性改變一致。相關研究發現灰質體積的增加與音樂專業性呈顯著正相關(Bermudez,Lerch, Evans, & Zatorre, 2009), 且分布在與音樂技能訓練高度相關的腦區, 如顳橫回(Schneider et al., 2002)、左側顳平面(Schlaug, Jancke, Huang, &Steinmetz, 1995)、顳上回(Bermudez et al., 2009)、中央溝內手指感覺區域(Li et al., 2010)、小腦(Hutchinson,Lee, Gaab, & Schlaug, 2003)等。也有研究發現音樂家的手指運動區(位于中央前回)的灰質結構更清晰和典型(Bangert & Schlaug, 2006)。相應的, 音樂家大腦中連接以上灰質區域的白質結構也發生特異性改變, 出現各項異性的增高, 如連接前運動皮層與感覺運動皮層的胼胝體前部和后部(?ztürk,Tas?ioglu, Aktekin, Kurtoglu, & Erden, 2002)、連接皮層?脊髓的皮質脊髓束(Rüber, Lindenberg, &Schlaug, 2015)等。總的來說, 音樂訓練對腦結構的特異性影響主要表現在與聽覺、手指運動與語義分析高度相關的皮層發生改變, 具體表現為皮層灰質體積的增大, 且連接這些區域的白質纖維微結構發生改變(各項異性增高)。
綜上所述, 舞蹈與音樂的腦可塑性研究呈現以下幾點不足:對音樂的關注遠遠多于舞蹈, 舞蹈的相關研究還有待于進一步豐富與深入。結構研究遠遠少于功能研究, 且有限的結構研究結果并不穩定。單獨研究舞蹈或音樂的可塑性改變較普遍, 而比較兩種訓練導致的差異非常有限, 僅有兩篇文章報道了舞蹈家與音樂家的白質與灰質結構的異同(Giacosa et al., 2016; Karpati, Giacosa,Foster, Penhune, & Hyde, 2017)。對兩種訓練導致的灰質體積變化結果存在爭議, 如 Giacosa等人(2016)發現舞蹈家與音樂家灰質體積在顳葉上部有共同的增加, 但最近發表的一項研究則報告舞蹈訓練對灰質結構的改變并無顯著影響(Burzynska,Finc, Taylor, Knecht, & Kramer, 2017)。除了上述不足, 我們的前期研究結果還無法完整描述舞蹈訓練的特異性影響。我們雖然發現舞蹈訓練對大腦皮層?基底節回路的連接具有增強作用(Li et al.,2015), 說明舞蹈訓練提高了受訓者的感覺運動整合功能, 但這種改變也發生在全身性運動的體育訓練中(Huang et al., 2017)。同時由于舞蹈與音樂關系最為密切, 它往往在音樂節奏中進行, 因此要刻畫舞蹈的特征就涉及對音樂效應的考慮。所以我們將研究重點放在兩種訓練的腦結構對比上,希望一定程度上分離舞蹈中的音樂影響, 進一步探索舞蹈訓練相關的腦結構特異性改變。基于前述的研究概況與深入探索舞蹈特征的需要, 我們選取接受現代舞與弦樂訓練的被試, 對兩種被試的腦灰質體積進行比較研究。選擇現代舞是因為它更接近舞蹈本質。相對于單純追求形式與技巧的芭蕾, 現代舞更強調人體動作本身。這正如表現主義現代舞創始人瑪麗.魏格曼在關于舞蹈本體論中所述, 舞蹈的核心因素是人體動作, 即人體動作構成的“力”的因素, 因此沒有音樂要素的舞蹈也能獨立存在(呂藝生, 2013)。選擇弦樂是因為它更典型地體現了音樂訓練中的聽?動整合。相對于吹奏類與鍵盤類樂器的訓練, 弦樂訓練對演奏者音準要求非常高。弦樂演奏音高的變化是演奏者根據聽覺反饋, 精準確定手指在琴弦上的位置而實現(馬清, 2000)。因此弦樂訓練涉及快速、密集和精確的聽?動整合。
本文的創新與意義在于:訓練組的類型齊整統一。我們選取相同舞蹈風格與樂器類型的被試,在腦結構可塑性分析研究中降低了舞蹈訓練風格與樂器不一致因素的干擾, 進而更好地描繪兩種訓練對腦結構的特異性影響。研究的問題具有探索性。關于兩種訓練對灰質結構影響的研究非常有限且存在爭議, 我們的研究能為這一問題提供新的證據。研究具有延續性。基于課題組前期研究發現, 我們通過比較進一步刻畫舞蹈訓練對腦結構的獨特影響。
本研究采用基于體素的形態學分析方法(voxelbased morphometry, VBM)進行組間灰質體積對比。我們預測訓練組的腦灰質體積可能存在如下改變:現代舞與弦樂訓練組的變化范圍主要位于感覺運動網絡; 現代舞訓練組可能出現灰質體積的減小, 而弦樂訓練組則可能是增加; 兩組被試還可能出現顳葉上部灰質體積的共同增高。
2 材料與方法
2.1 研究對象及納入標準
本研究招募了3組被試, 包括18名現代舞被試(現代舞訓練組)、20名弦樂被試(弦樂訓練組)及25名對照被試(對照組) (見表1)。訓練組被試均為大學藝術特長生或藝術專業學生, 其中現代舞訓練組主修現代舞, 弦樂訓練組主修西洋弦樂(小提琴、中提琴或大提琴)。在納入本實驗前, 兩組專業被試分別接受兩名舞蹈或兩名音樂專業教師的綜合評估, 其專業性被評價為達到舞蹈專長或音樂專長條件。舞蹈教師對現代舞訓練組被試進行如下評估:舞蹈模仿。向被試示范3次長度為 4個八拍的全新舞蹈動作(被試之前從未訓練過), 要求被試根據自己的記憶重復舞蹈片段, 評估其動作完整性與準確性。舞蹈基本功。要求被試完成下橫豎叉、下腰等動作, 同時要求被試根據自己的軟開度進行跳、轉、翻的技巧組合, 評估其柔韌度與技巧。舞蹈即興。要求被試根據隨機選擇的音樂進行即興舞蹈, 評估其舞蹈動作運用與舞蹈表現力。音樂教師對弦樂訓練組被試進行如下評估:自選曲目演奏。要求被試演奏一首自選曲目(3分鐘以內), 考察被試的器樂演奏與表現力。視奏能力。音樂教師譜寫一段16個小節的中等難度的演奏片段(被試之前從未演奏或練習過), 要求被試現場演奏, 考察其識譜演奏能力。現代舞訓練組被試均無音樂訓練經歷, 弦樂訓練組被試均無舞蹈訓練經歷。兩組專業被試訓練年限從7至17年不等。對照組為與之匹配的大學生,無任何舞蹈、音樂訓練經驗。現代舞訓練組、弦樂訓練組及對照組在年齡、性別和受教育年限方面均無統計顯著性差異, 兩種訓練組在訓練年限方面無統計顯著性差異, 分組達到均衡性(見表1)。3組被試完成艾丁伯格手性問卷(Oldfield, 1971)測試, 均為右利手。所有被試均無神經系統疾病、發展性疾病或物質濫用。本實驗符合赫爾辛基宣言, 并獲得作者所在研究機構倫理委員會許可,所有被試簽署實驗知情同意書。
2.2 數據獲取
本研究在電子科技大學信息醫學中心完成。MRI數據的采集設備為GE Discovery 750 3.0T磁共振系統(GE Medical Systems, Milwaukee, WI),使用標準的GE全頭線圈。高分辨率T1加權圖像通過3D梯度回波序列(T1-3DFSPGR)獲得。掃描參數為:TR = 6.008 ms, TE = 1.984 ms, 反轉角 9?,矩陣 256×256, FOV=25.6 cm × 20 cm, 層厚 1 mm,層數 152層。采集數據時, 被試在掃描儀中盡量保持頭部不動。實驗使用海綿來固定被試頭部以減少頭動, 使用耳塞來降低掃描過程中的噪音。
2.3 數據處理及統計分析
VBM是一種全腦無偏算法, 用來計算腦體積與容量(Ashburner & Friston, 2000, 2005)。本研究采用VBM8工具包進行灰質分析。(1)首先對3組被試的T1加權腦圖像進行數據預處理, 包括檢查

表1 被試人口學信息
所有被試數據的成像質量, 手動調整所有結構 MRI的原點到前連合。(2)其后在 MATLAB 2014環境下使用嵌套于SPM8 (http://www.fil.ion.ac.uk/spm)內的VBM8工具包對被試三維T1加權腦圖像進行常規的灰質體積估計流程:1)將所有被試的結構MRI圖像分割成灰質、白質和腦脊液。2)使用DARTEL算法對分割后的灰質圖像建立基于組水平的模板。3)利用構建的模板將所有灰質圖像配準到蒙特利爾神經研究所(Montreal Neurological Institute, MNI)標準人腦模板空間。4)將配準后的圖像進行調制得到灰質體積圖像。5)使用半高全寬為 8毫米高斯核將配準后的灰質圖像進行平滑。(3)最后對3組被試平滑后的灰質MRI圖像進行組間單因素方差分析(進行高斯隨機場理論校正, 校正后p< 0.05) (Nichols, 2012), 進而對差異腦區通過兩兩比較進行事后檢驗分析(雙樣本t檢驗,p< 0.05)。
3 結果
較之對照組, 現代舞訓練組與弦樂訓練組均表現出灰質結構的特異性改變(見表2)。其中現代舞訓練組的雙側丘腦、右側輔助運動皮層(SMA)與左側中央前回(M1)灰質體積顯著低于對照組和弦樂訓練組; 其右側眶部額上回、右側殼核與左側小腦則顯著高于其他兩組(p< 0.05,t檢驗) (見圖1)。弦樂訓練組的右側額中回、右側顳上回與左側中央前回灰質體積顯著大于對照組和現代舞訓練組(p< 0.05,t檢驗) (見圖 1)。

表2 現代舞訓練組、弦樂訓練組與對照組灰質體積的組間比較
4 討論
VBM分析結果表明, 較之對照組現代舞訓練組與弦樂訓練組的灰質體積存在特異性改變, 兩者灰質體積改變的區域和方式存在差異。現代舞訓練組的改變涉及左側小腦、雙側丘腦、右側殼核、右側輔助運動區等有關感覺運動控制的廣泛腦區, 不同區域灰質體積既有增加也有減小。弦樂訓練組的改變集中于聽?動?讀相關腦區, 并統一表現為增加的灰質體積。組間灰質體積的特異性改變說明現代舞訓練與弦樂訓練對腦可塑性的不同影響。
4.1 現代舞訓練對腦灰質體積的影響
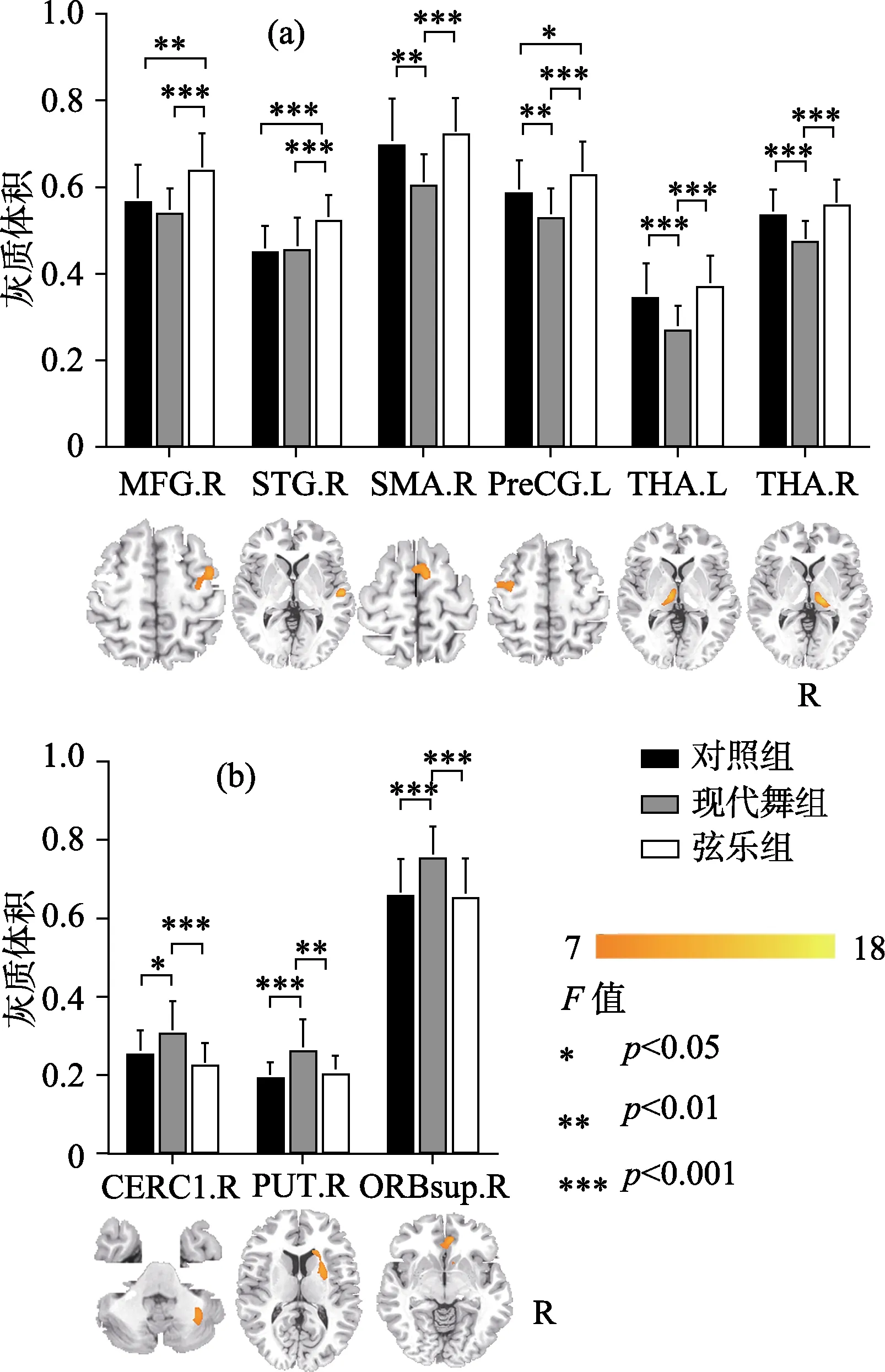
圖1 現代舞訓練組、弦樂訓練組與對照組灰質體積的組間比較
較之弦樂訓練組與對照組, 現代舞訓練組大腦灰質體積變化區域更廣泛, 在這些區域中同時存在灰質體積的增高與降低。該結果可能反映了全身性舞蹈動作訓練對腦的特異性影響。Brown等人(2006)利用正電子發射斷層成像(positron emission tomography, PET)技術觀察到舞者進行下肢舞步移動時的腦功能狀態, 結果發現小腦蚓部、右側殼核、腹側丘腦、內側上小葉、輔助運動皮層(SMA)、初級運動皮層(M1)與前運動皮層區域的激活明顯。我們的結果基本屬于以上功能區內,從結構上說明舞蹈訓練可能影響感覺運動網絡內的廣泛腦區。另外, 現代舞訓練組灰質體積的增減也可能體現某種結構與功能的優化。增高的灰質體積往往意味著該區域的神經元更多, 處理信息的效率更高, 對應的認知和行為功能更強大。大部分相關研究表明訓練水平往往與灰質體積成正比(Maguire et al., 2000)。與此一致, 現代舞訓練組在動作控制相關腦區中表現出增高的灰質體積:比如眶部額上回, 該區域與身體平衡維持相關(Taubert et al., 2010); 小腦下部, 該區域將本體感覺信息與運動前庭功能相整合, 與平衡保持及姿勢維持高度相關(Kheradmand & Zee, 2011)。現代舞訓練要求舞者不斷創造自發動作, 很多自發動作的組合可能是全新的, 因此需要大腦付出更多努力來進行各種動作和姿態的調控與保持, 進而導致以上兩個區域灰質體積的增大。其次現代舞訓練組還表現出右側殼核灰質體積的增高與雙側丘腦灰質體積的降低。這一相反變化可能與殼核與丘腦在節奏運動中的功能有關。相關研究報道舞蹈家的右側殼核在節奏運動條件下激活明顯,而丘腦在非節奏復雜運動時有明顯激活(Brown et al., 2006)。我們前期的研究也發現舞蹈家的殼核與全腦的功能連接更顯著(Li et al., 2015)。現代舞訓練強調動作性, 常常通過動作的力度變化來表現舞蹈情感, 而這種力度變化往往借助于動作節奏的改變而實現。因此, 在現代舞訓練中不僅需要舞者保持動作的節奏性, 還需要在不同節奏間進行動作切換。這必然需要殼核的高度參與而丘腦保持抑制, 因此可能導致這兩個區域相反的灰質體積變化。進一步我們還發現現代舞訓練組右側輔助運動皮層(SMA)與左側初級運動皮層(M1)的體積顯著減小。這兩個腦區在復雜運動控制和感覺運動表征方面扮演重要角色, 是參與舞蹈過程的重要腦區。關于芭蕾舞蹈家和體操運動員的相關研究均發現這兩個腦區的灰質體積顯著降低(H?nggi et al., 2010; Huang, Lu, Song, & Wang,2015), 這可能體現了全身性運動技能訓練的特殊效應。綜上所述, 現代舞訓練所影響的灰質結構變化范圍更廣泛, 灰質體積增高與降低的區域并存。同時這些變化都分布于皮層?基底節?丘腦?小腦神經回路內(該回路貫穿全腦, 與高級認知、運動調控高度相關) (Bostan, Dum, & Strick, 2013)。以上結果可能反映了現代舞獨有的訓練效應。相對于注重形式和規范的傳統舞蹈, 現代舞是一種更為自由和靈活的舞種, 強調創造性的自發動作,要求舞者容納和表現更為復雜和廣泛的內外感受。這一特點可能需要腦在更大尺度上去加工和整合信息。因此我們推測現代舞訓練影響的可能是涉及全腦感覺運動功能協同的廣泛腦區。
4.2 弦樂訓練對腦灰質體積的影響
較之對照組和現代舞訓練組, 弦樂訓練組灰質體積的改變集中于與音樂技能直接相關的腦區,且全部是增高。這一發現顯示弦樂訓練這種聽?動結合的精細運動對大腦的特異性影響。聽是音樂家的重要能力之一。我們的結果表明弦樂訓練組在右側聽皮層(顳上回)有顯著的體積增高。相關元分析研究顯示, 顳上回在音樂訓練中協調聽覺系統對錯誤的檢查, 負責對音樂旋律的解碼和音樂語義的記憶并參與音樂中的動?聽整合過程(周臨舒, 趙懷陽, 蔣存梅, 2017; Lahav, Saltzman, &Schlaug, 2007)。此外大腦聽皮層對音樂的處理具有某種程度的偏側性, 其中右側顳上回對音樂更為敏感(Hyde, Peretz, & Zatorre, 2008)。相關的結構研究也發現音樂家顳上回的灰質體積顯著增大,其體積與音樂訓練時間成正比(Groussard et al.,2014)。除了聽覺皮層的變化, 我們發現弦樂訓練組左側初級運動皮層(M1)的灰質體積顯著增加。這個區域是所有運動執行的核心區域, 類似的結果也出現在各類器樂演奏家的灰質結構研究中(Bermudez et al., 2009; Han et al., 2009; Sluming et al., 2002)。進一步我們還發現弦樂訓練組右側額中回灰質體積的增加。這個區域與工作記憶的處理與存儲相關(Jones, Adlam, Benattayallah, & Milton, 2017), 且與顳中回共同完成語義啟動(Laufer, Negishi, Lacadie,Papademetris, & Constable, 2011)。相關研究發現音樂家在這一部位的灰質體積是增加的(Bermudez et al., 2009)。弦樂訓練需要受訓者識別樂譜符號并記憶樂譜, 這個過程會頻繁調用語義表征與工作記憶的腦區, 可能促成該腦區灰質結構的特異性改變。綜上所述, 弦樂訓練對皮層的影響可能更為集中, 其灰質體積變化區域位于與訓練高度相關的聽覺、運動與語義加工皮層。這些區域表現出灰質體積的一致增高。我們推測這種改變可能與弦樂訓練要求的高度精準性相關。相對于其他樂器訓練, 弦樂訓練需要更為細微精準的手指控制和穩定的內部音高知覺。受訓者需要通過頻繁的聽?動信息整合與反饋, 才能進行正確演奏。因此弦樂訓練可能導致與訓練技能直接相關的皮層發生改變。
4.3 本研究的有趣發現
4.3.1 初級運動皮層
初級運動皮層(M1)負責發出運動控制指令及對運動進行表征, 它參與所有動作的執行, 是舞蹈與音樂訓練相關的重要腦區。有趣的是, 我們發現現代舞訓練組和弦樂訓練組灰質體積在左側M1出現了相反的改變趨勢。現代舞訓練組較之對照組與弦樂訓練組灰質體積顯著降低, 而弦樂訓練組則顯著增高。現代舞訓練組 M1灰質體積的減小可能是神經修剪和優化的結果。通過科學專業的動作訓練, 能讓相關腦區內的神經元去除冗余突觸, 增強局部回路內部以及遠距離腦區間的連接, 從而建立更為高效的神經連接模式(段旭君,2013)。這一神經可塑性改變外化于行為就是自動化舞蹈動作的獲得, 每個舞蹈動作調用較少的神經資源就可以完成。弦樂訓練組M1灰質體積的增高則可能是弦樂高度特異性的技能所致。M1在手指或手部的有序運動時會被明顯激活(Shibasaki et al., 1993)。同時該區域對運動頻率非常敏感, 它的激活程度隨著頻率的增加而增大(Turner, Grafton,Votaw, Delong, & Hoffman, 1998)。弦樂訓練是精細有序的手指運動, 要求給予高頻快速的動作反饋, 這種訓練方式可能會強烈地激活M1, 進而導致該區域灰質體積的增大。
4.3.2 顳上回
Karpati等人(2016)發現舞蹈家與音樂家灰質體積在這個區域有共同的增加, 并推測該結果與音樂舞蹈訓練所涉及的大量動?聽整合相關。我們的結果與他們相左, 僅在弦樂訓練組中發現灰質體積的顯著增高, 而現代舞訓練組沒有顯著變化。其原因可能來自被試的差異。現代舞的典型特點是關注自發動作本身, 它并不高度依賴音樂線索, 甚至可以在無音樂的背景下進行。因此現代舞訓練組涉及的動?聽整合訓練并不突出, 顳上回的灰質體積改變不顯著。而弦樂訓練則需要高度依賴聽覺信息來精確調控手指動作, 更有力地訓練了演奏者的動?聽整合功能, 因此導致顳上回灰質體積的顯著增高。
5 結論
本研究采用基于體素的形態學分析方法, 比較現代舞訓練與弦樂訓練對腦灰質體積的可塑性影響。結果表明, 兩種訓練都使腦灰質體積發生特異性改變。其中現代舞訓練的影響區域更為廣泛, 分布于皮層?基底節?丘腦?小腦回路內, 其變化既有增加也有減小。這一結果說明現代舞訓練影響的可能是涉及復雜感覺運動整合的廣泛腦區。弦樂訓練影響的則是與訓練特性(動?聽)直接相關的腦區。通過比較研究, 我們的結果首次展示了現代舞訓練與弦樂訓練對腦灰質結構影響的差異, 揭示了舞蹈訓練對腦的特異性影響, 進而為舞蹈治療和音樂治療提供神經學基礎。此外,我們的結果也提示后續有兩個問題需要進一步探索:其一, 長期舞蹈訓練對關鍵腦區的影響方式是怎樣的?我們發現現代舞訓練組存在灰質體積增高的區域, 如小腦與殼核, 而大部分相關研究報告舞蹈家或體操運動員的灰質體積都是減小的。那么腦對舞蹈訓練的適應到底是協調各腦區皮層以增減互補的方式實現?還是以減小優化的方式達成?其二, 如何理解舞蹈訓練與音樂訓練對皮層體積影響的相反趨勢? 我們發現現代舞訓練組與弦樂訓練組均在 M1出現顯著的灰質體積改變, 但這種改變趨勢卻是相反的。這一結果否說明皮層的體積增加直接與局部功能提升相關,而減小則可能提示是在一個系統內發生整體性優化的結果。以上問題均需要進一步的實證研究進行闡釋。
段旭君. (2013).基于大尺度腦網絡分析方法的腦可塑性研究(博士學位論文).電子科技大學, 成都.
蔣存梅. (2016).音樂心理學. 上海: 華東師范大學出版社.
馬清. (2000).音樂理論與管弦樂基礎. 北京: 北京大學出版社.
平心. (2004).舞蹈心理學.北京: 高等教育出版社.
呂藝生. (2003).舞蹈學導論.上海: 上海音樂出版社.
覃嬪. (2018).舞蹈藝術的訓練研究.北京: 北京理工大學出版社.
周臨舒, 趙懷陽, 蔣存梅. (2017). 音樂表演訓練對神經可塑性的影響: 元分析研究.心理科學進展, 25(11), 1877–1887.
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry--the methods.NeuroImage, 11, 805–821.
Ashburner, J., & Friston, K. J. (2005). Unified segmentation.NeuroImage, 26(3), 839–851.
Bangert, M., Peschel, T., Schlaug, G., Rotte, M., Drescher, D.,Hinrichs, H., ... Altenmüller, E. (2006). Shared networks for auditory and motor processing in professional pianists:Evidence from fMRI conjunction.NeuroImage, 30(3),917–926.
Bangert, M., & Schlaug, G. (2006). Specialization of the specialized in features of external human brain morphology.European Journal of Neuroscience, 24(6), 1832–1834.
Baumann, S., Koeneke, S., Schmidt, C. F., Meyer, M., Lutz,K., & Jancke, L. (2007). A network for audio-motor coordination in skilled pianists and non-musicians.Brain Research, 1161, 65–78.
Bermudez, P., Lerch, J. P., Evans, A. C., & Zatorre, R. J.(2009). Neuroanatomical correlates of musicianship as revealed by cortical thickness and voxel-based morphometry.Cerebral Cortex, 19(7), 1583–1596.
Bostan, A. C., Dum, R. P., & Strick, P. L. (2013). Cerebellar networks with the cerebral cortex and basal ganglia.Trends in Cognitive Sciences, 17(5), 241–254.
Brown, S., Martinez, M. J., & Parsons, L. M. (2006). The neural basis of human dance.Cerebral C ortex, 16(8),1157–1167.
Burzynska, A. Z., Finc, K., Taylor, B. K., Knecht, A. M., &Kramer, A. F. (2017). The dancing brain: Structural and functional signatures of expert dance training.Frontiers in Human Neuroscience, 11, 566.
Calvo-Merino, B., Glaser, D. E., Grèzes, J., Passingham, R.E., & Haggard, P. (2005). Action observation and acquired motor skills: An FMRI study with expert dancers.Cerebral Cortex, 15(8), 1243–1249.
Cross, E. S., Hamilton, A. F., & Grafton, S. T. (2006). Building a motor simulation de novo: Observation of dance by dancers.NeuroImage, 31(3), 1257–1267.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn,U., & May, A. (2004). Neuroplasticity: Changes in grey matter induced by training.Nature, 427(6972), 311–312.
Giacosa, C., Karpati, F. J., Foster, N. E. V., Penhune, V. B.,& Hyde, K. L. (2016). Dance and music training have different effects on white matter diffusivity in sensorimotor pathways.NeuroImage, 135, 273–286.
Groussard, M., Rauchs, G., Landeau, B., Viader, F.,Desgranges, B., Eustache, F., & Platel, H. (2010). The neural substrates of musical memory revealed by fMRI and two semantic tasks.NeuroImage, 53(4), 1301–1309.
Groussard, M., Viader, F., Landeau, B., Desgranges, B.,Eustache, F., & Platel, H. (2014). The effects of musical practice on structural plasticity: The dynamics of grey matter changes.Brain and Cognition, 90, 174–180.
Han, Y., Yang, H., Lv, Y. T., Zhu, C. Z., He, Y., Tang, H.H., ... Dong, Q. (2009). Gray matter density and white matter integrity in pianists' brain: A combined structural and diffusion tensor MRI study.Neuroscience Letters, 459(1),3–6.
H?nggi, J., Koeneke, S., Bezzola, L., & J?ncke, L. (2010).Structural neuroplasticity in the sensorimotor network of professional female ballet dancers.Human Brain Mapping,31(8), 1196–1206.
Huang, H. Y., Wang, J. J., Seger, C., Min, L., Feng, D., Wu,X. Y., ... Huang, R. W. (2017). Long-term intensive gymnastic training induced changes in intra- and inter-network functional connectivity: An independent component analysis.Brain Structure and Function, 223(1), 131–144.
Huang, R. W., Lu, M., Song, Z., & Wang, J. (2015).Long-term intensive training induced brain structural changes in world class gymnasts.Brain S tructure and Function, 220(2), 625–644.
Hutchinson, S., Lee, L. H. L., Gaab, N., & Schlaug, G.(2003). Cerebellar volume of musicians.Cerebral Cortex,13(9), 943–949.
Hyde, K. L., Peretz, I., & Zatorre, R. J. (2008). Evidence for the role of the right auditory cortex in fine pitch resolution.Neuropsychologia, 46(2), 632–639.
Jola, C., McAleer, P., Grosbras, M. H., Love, S. A., Morison,G., & Pollick, F. E. (2013). Uni- and multisensory brain areas are synchronised across spectators when watching unedited dance recordings.i-Perception, 4(4), 265–284.
Jones, J., Adlam, A., Benattayallah, A., & Milton, F. (2017,July).Working memory training increases recruitment ofthe middle fr ontal g yrus in children. Poster session presented at the Conference of Experimental Psychology Society, Reading, UK.
Karpati, F. J., Giacosa, C., Foster, N. E. V., Penhune, V. B.,& Hyde, K. L. (2017). Dance and music share gray matter structural correlates.Brain Research, 1657, 62–73.
Kheradmand, A., & Zee, D. S. (2011). Cerebellum and ocular motor control.Frontiers in Neurology, 2, 53.
Koelsch, S., & Siebel, W. A. (2005). Towards a neural basis of music perception.Trends in Cognitive Sciences, 9(12),578–584.
Lahav, A., Saltzman, E., & Schlaug, G. (2007). Action representation of sound: Audiomotor recognition network while listening to newly acquired actions.Journal o f Neuroscience, 27(2), 308–314.
Laufer, I., Negishi, M., Lacadie, C. M., Papademetris, X., &Constable, R. T. (2011). Dissociation between the activity of the right middle frontal gyrus and the middle temporal gyrus in processing semantic priming.PLoS O ne, 6(8),e22368.
Li, G. J., He, H., Huang, M. T., Zhang, X. X., Lu, J., Lai, Y.X., ... Yao, D. Z. (2015). Identifying enhanced corticobasal ganglia loops associated with prolonged dance training.Scientific Reports, 5, 10271.
Li, S. Y., Han, Y., Wang, D. Y., Yang, H., Fan, Y. B., Lv, Y.T., ... He, Y. (2010). Mapping surface variability of the central sulcus in musicians.Cerebral Cortex, 20(1), 25–33.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D.,Ashburner, J., Frackowiak, R. S. J., & Frith, C. D. (2000)Navigation-related structural change in the hippocampi of taxi drivers.Proceedings of the National Academy ofSciencesof the United States of America, 97(8), 4398–4403.
Mutschler, I., Schulze-Bonhage, A., Glauche, V., Demandt,E., Speck, O., & Ball, T. (2007). A rapid sound-action association effect in human insular cortex.PLoS One, 2(2),e259.
Nichols, T. E. (2012). Multiple testing corrections,nonparametric methods, and random field theory.NeuroImage, 62(2), 811–815.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburgh inventory.Neuropsychologia,9(1), 97–113.
Ono, Y., Nomoto, Y., Tanaka, S., Sato, K., Shimada, S.,Tachibana, A., ... Noah, J. A. (2014). Frontotemporal oxyhemoglobin dynamics predict performance accuracy of dance simulation gameplay: Temporal characteristics of top-down and bottom-up cortical activities.NeuroImage,85, 461–470.
?ztürk, A. H., Tas?ioglu, B., Aktekin, M., Kurtoglu, Z., &Erden, I. (2002). Morphometric comparison of the human corpus callosum in professional musicians and nonmusicians by using in vivo magnetic resonance imaging.Journal of Neuroradiology, 29(1), 29–34.
Rüber, T., Lindenberg, R., & Schlaug, G. (2015). Differential adaptation of descending motor tracts in musicians.Cerebral Cortex, 25(6), 1490–1498.
Schlaug, G., Jancke, L., Huang, Y., & Steinmetz, H. (1995).In vivo evidence of structural brain asymmetry in musicians.Science, 267(5198), 699–701.
Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J.,Gutschalk, A., & Rupp, A. (2002). Morphology of Heschl's gyrus reflects enhanced activation in the auditory cortex of musicians.Nature Neuroscience, 5(7), 688–694.
Shibasaki, H., Sadato, N., Lyshkow, H., Yonekura, Y., Honda,M., Nagamine, T., ... Konishi, J. (1993). Both primary motor cortex and supplementary motor area play an important role in complex finger movement.Brain, 116,1387–1398.
Sluming, V., Barrick, T., Howard, M., Cezayirli, E., Mayes,A., & Roberts, N. (2002). Voxel-based morphometry reveals increased gray matter density in Broca's area in male symphony orchestra musicians.NeuroImage, 17(3),1613–1622.
Taubert, M., Draganski, B., Anwander, A., Muller, K.,Horstmann, A., Villringer, A., & Ragert, P. (2010). Dynamic properties of human brain structure: Learning-related changes in cortical areas and associated fiber connections.Journal of Neuroscience, 30(35), 11670–11677.
Turner, R. S., Grafton, S. T., Votaw, J. R., Delong, M. R., &Hoffman, J. M. (1998). Motor subcircuits mediating the control of movement velocity: A PET study.Journal of Neurophysiology, 80(4), 2162–2176.
猜你喜歡
小天使·一年級語數英綜合(2020年3期)2020-12-16 02:56:12
河北畫報(2020年8期)2020-10-27 02:55:14
藝術啟蒙(2018年7期)2018-08-23 09:14:16
藝術家(2018年4期)2018-01-26 01:32:09
兒童繪本(2017年24期)2018-01-07 15:51:37
東方藝術·大家(2016年6期)2016-09-05 07:30:56
漢語世界(2016年5期)2016-03-15 06:46:41
黃河之聲(2016年23期)2016-02-03 04:44:53
新教育時代電子雜志(學生版)(2015年31期)2015-12-20 08:29:49
戲劇之家(2015年21期)2015-12-04 00:36:22
