哀牢山濕性常綠闊葉林地生、樹干及樹枝附生苔蘚生活型組成及其水分特性
2019-09-10 07:22:44范曉陽劉文耀宋亮陳泉袁國迪
廣西植物 2019年5期
關鍵詞:生活型
范曉陽 劉文耀 宋亮 陳泉 袁國迪



摘 要:為探討林內不同垂直高度生境下及不同生活型苔蘚植物水分特征的差異,該文對哀牢山濕性常綠闊葉林內地生苔蘚、林下樹干附生和林冠層樹枝附生苔蘚優勢或常見種的生活型組成、持水力、失水特征和水分利用效率進行了研究。結果表明:地生、樹干附生和樹枝附生苔蘚植物類群分別以交織型、扇型、懸垂型苔蘚植物占據優勢;地生、樹干附生、枝條附生苔蘚植物的飽和持水率分別為476%DW、210.98%DW、238.95%DW;地生苔蘚植物的持水率和失水速率均高于附生苔蘚,樹干附生苔蘚植物持水率低于樹枝附生苔蘚,而失水速率高于樹枝附生苔蘚。在不同生活型苔蘚的水分特性上,交織型苔蘚具有較高的持水率和失水速率,保水性能較弱,其次為懸垂型苔蘚,扇型苔蘚的持水率最小,失水速率也較快;地生和樹干附生苔蘚的水分利用效率均顯著高于樹干附生苔蘚,交織型和扇型苔蘚的水分利用效率顯著高于懸垂型苔蘚。在三種生境下,地生苔蘚持水力高,水分利用效率較高而保水能力低;枝條附生苔蘚持水力低,水分利用效率低而保水能力較高;樹干附生苔蘚水分利用效率較高而持水力和保水能力均較差。因此,不同生境下苔蘚植物生活型組成及其水分變化特性在一定程度上反映了它們對不同生境的適應策略。
關鍵詞: 苔蘚植物, 持水能力, 失水曲線, 地生, 附生, 生活型
中圖分類號:Q948.1
文獻標識碼:A
文章編號:1000-3142(2019)05-0668-13
Life-form composition and water characteristics ofterrestrial, bole and branch epiphytic bryophytes in amoist evergreen broad-leaved forest in Ailao Mountains
FAN Xiaoyang1, 2, LIU Wenyao1*, SONG Liang1, CHEN Quan1, 2, YUAN Guodi1,2
(1. CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences,Kunming 650223 , China; 2. University of Chinese Academy of Sciences, Beijing 100049, China )
Abstract:In order to explore ecological adaptation strategies of bryophytes for different water conditions, we determined the water loss characteristics during natural desiccation and water use efficiency of dominated or common bryophyte species from terrestrial, trunk and branch in a moist evergreen broad-leaved forest in Ailao Mountains. Our results demonstrated that weft, fan and pendant life-forms were dominated in terrestrial, bole epiphytic and branch epiphytic bryophytes respectively. Bryophytes from different microhabitats exhibited great variances of water-holding capacity and water loss rate: Water-holding capacity of bryophytes from three microhabitats was 476%DW, 210.98%DW and 238.95%DW respectively; Water-holding capacity and water loss rate of terrestrial bryophytes were higher than these of the trunk- and branch-dwellers. Compared with branch-dwellers, the bole epiphytic bryophytes had lower water-holding capacity and higher water loss rate. Water holding capacity and water loss rate of bryophytes among different life-forms also exhibited significant differences: Weft life-form bryophytes exhibited higher water-holding capacity and higher water loss rate, indicating their weaker water retention capacity, followed by pendant bryophytes, and fan bryophytes had lowest water holding capacity and highest water loss rate. Water use efficiency of terrestrial bryophytes and bole epiphytic bryophytes were significantly higher than branch-dwellers. Weft and fan life-form bryophytes had higher water use efficiency than that of pendant bryophytes. The results indicated that water-holding capacity and water use efficiency of terrestrial bryophytes was higher while water retention capacity was poor; Water retention capacity of branch-dwellers was higher while water-holding capacity and water use efficiency was lower; Water use efficiency of trunk-dwellers was higher while both water-holding capacity and water retention capacity was poor. Life-form composition and water characteristics of bryophytes from terrestrial bryophytes, bole and branch epiphytic bryophytes indicated their adaptation strategies to different habitats to a certain extent. The results would provide a scientific reference forelucidating hydrological features of bryophytes under different microhabitats, and understanding ecological adaptation strategies of bryophyte.
Key words: bryophytes, water-holding capacity, water-loss curve, terrestrial bryophytes, epiphytes, life-form
苔蘚植物是一類古老而獨特的高等植物, 是植物界從水生向陸生過渡的代表類群(Smith, 1982),其種類繁多(Schofield, 1985),生態適應性強(Kruys & Jonsson, 1999; Farrant & Moore, 2011),能夠在不同生境中生長和分布。由于苔蘚植物對生長基質的要求并不高, 在林冠樹枝、樹干、林下木質殘體、林地和巖石表面均能發現所對應的苔蘚植物, 但不同的生長基質上的苔蘚植物在物種組成上有所不同(Humphrey et al., 2002; 馬文章,2011)。作為山地森林生態系統中重要的結構性組分,苔蘚植物在維持山地森林生態系統生物多樣性、養分與水分循環均發揮著重要的作用,對森林生態系統水分涵養具有重要的生態學意義(郭水良等,1999;徐海清和劉文耀,2005)。經過漫長的生態適應與演變過程,苔蘚植物形成了不同的生活型,其不僅涵蓋了苔蘚植物生長位置和分枝類型等形態學特征,而且還包括了苔蘚植物對生境的拓展策略等生態信息。相同生活型的苔蘚植物具有相似的生態特征(Bates, 1998)和競爭策略(Mlson & Rydin, 2009)。
附生苔蘚植物是山地森林生態系統中一類重要而特殊的植物類群。相對于地生苔蘚植物,附生苔蘚植物不與土壤直接相連,其個體矮小,葉表面積大,其生命活動所需要的水分和養分離子都來自于雨水或空氣中的凝結水,對環境因子變化尤為敏感和脆弱(官飛榮等,2017);作為變水植物,附生苔蘚植物無表皮細胞,不能有效地保存水分,空氣干燥時很快喪失水分及光合能力(劉文耀等,2006)。地生與附生苔蘚植物由于各自的生長基質水分與營養狀況的差異,使得它們在物種組成上差異顯著(Kürschner, 2004),地生與附生苔蘚植物在持水能力及水分利用特征上的差異對其在不同生境下的生存和生長極其重要。在森林不同高度上,光強、溫度、溫度波動、風速等隨著垂直高度的增加而升高,而空氣濕度隨之降低(Petter et al.,2016),使得林冠層附生苔蘚比林下苔蘚植物面臨著更頻繁的干濕交替。目前,對苔蘚植物持水能力及其水分利用效率的研究主要集中在不同區域、不同植被類型的林下地表苔蘚層(包維楷等,2004;張洪江等,2003;車宗璽等,2006;張顯強等,2012;姬明飛和韓鴻基,2017),而鮮有研究關注不同生境下尤其林內不同垂直梯度生境下苔蘚植物的持水能力及水分利用特征的差異。
云南省哀牢山國家自然保護區保存著目前我國亞熱帶地區面積最大的原生中山濕性常綠闊葉林,森林結構保存較為完整,林地、樹干及樹枝苔蘚植物發達。
Ma et al.(2009)和馬文章等(2011)對于本區森林中不同生長基質上苔蘚植物的組成與分布特征進行了調查研究,為本研究的開展提供了較好的基礎。本研究以云南哀牢山中山濕性常綠闊葉林內地生與附生苔蘚植物為對象,通過對林地、樹干附生和冠層樹枝附生不同空間層次上苔蘚植物的物種與生活型組成、吸持水能力、失水特征及水分利用效率的試驗研究,探討不同生長基質及生境條件下苔蘚植物的水分變化特征與水分利用效率的差異性及其形成機理,為深入研究山地森林生態系統生物多樣性形成格局及其維持機制,理解不同生境下苔蘚植物的水文功能,進一步揭示苔蘚植物對不同水分條件環境的生態適應策略提供科學依據。
1 材料與方法
1.1 研究區概況
哀牢山(100°44′—101°30′ E,23°36′—24°56′ N)位于云貴高原西南部、橫斷山脈南段,屬云嶺向南分支的余脈,延綿500余km,降雨充沛(邱學忠和謝壽昌,1998)。本研究區位于哀牢山徐家壩地區(101.01° E,23.32° N),海拔為2 000~2 750 m,年均降水量為1 947mm,年蒸發量為1 192mm,該區氣候干濕季分明,降雨主要集中在每年的5—10月,占全年雨量在85%以上,11月至次年4月降雨則顯著減少,年均相對濕度為85%,年均氣溫為11.3 ℃,氣候終年溫涼潮濕(李蘇等,2007)。
該區域保存著我國亞熱帶地區最大面積(約5 100hm2)、以木果柯(Lithocarpus xylocarpus)、變色錐(Castanopsis wattii)、硬殼柯(Lithocarpus hancei)為優勢的原生中山濕性常綠闊葉林(Liu et al.,2002;徐海清和劉文耀,2005), 林冠高度為18~25 m, 其群落蓋度在95% 以上,附生植物非常豐富,其中林地上苔蘚生長茂盛,樹干及樹枝苔蘚植物密布,構成濕性常綠闊葉林的獨特景觀(游承俠,1983)。根據哀牢山生態站的觀測結果及有關研究結果(劉玉洪,1993),在中山濕性常綠闊葉林中,林地、林下樹干及林冠上層樹枝區是三種光照、溫度、濕度等因子差異明顯的生境(圖1)。
1.2 研究材料的選擇與采集
由于生境條件及生長基質的不同,在苔蘚植物的物種組成與分布上存在較大的差異。根據
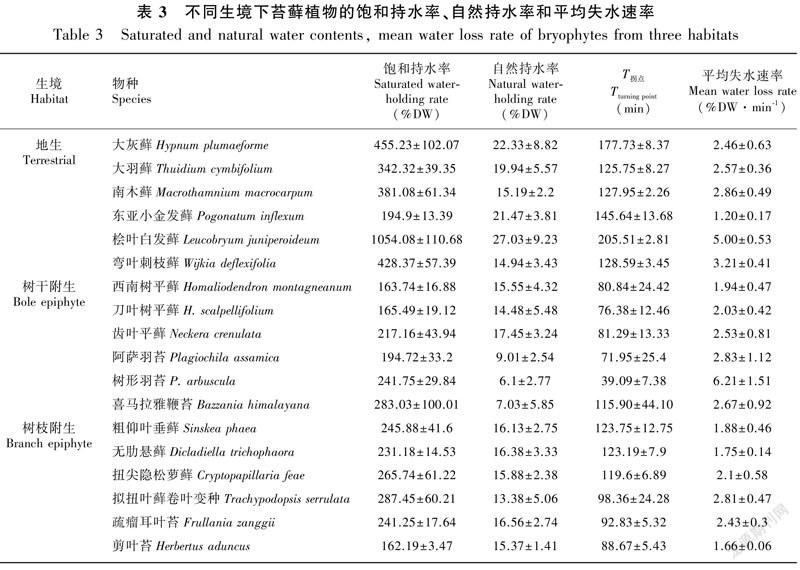
Ma et al.(2009)和馬文章等(2011)對哀牢山徐家壩地區山地濕性常綠闊葉林內苔蘚植物組成與分布特征的研究結果,結合不同生境中苔蘚植物的出現頻率、重要值等指標,我們從地生、林下樹干附生和林冠上層樹枝附生三種生境中分別選擇6種占優勢或常見的苔蘚植物,共計18種苔蘚植物,其中地生苔蘚為東亞小金發蘚(Pogonatum inflexum)、南木蘚(Macrothamnium macrocarpum)、彎葉刺枝蘚(Wijkia deflexifolia)、大灰蘚(Hypnum plumaeforme)、大羽蘚(Thuidium cymbifolium)和檜葉白發蘚(Leucobryum juniperoideum);林下樹干附生苔蘚為西南樹平蘚(Homaliodendron montagneanum)、刀葉樹平蘚(H. scalpellifolium)、樹形羽苔(Plagiochila arbuscula)、阿薩羽苔(P. assamica)、齒葉平蘚(Neckera crenulata)和喜馬拉雅鞭苔(Bazzania himalayana);林冠上層樹枝附生苔蘚為粗仰葉垂蘚(Sinskea phaea)、扭尖隱松蘿蘚(Cryptopapillaria feae)、無肋懸蘚(Dicladiella trichophaora)、擬扭葉蘚卷葉變種(Trachypodopsis serrulata)、疏瘤耳葉苔(Frullania zanggii)和剪葉苔(Herbertus aduncus)。于2017年3—4月在哀牢山徐家壩中山濕性常綠闊葉林中三種生境內采集上述苔蘚物種,每個物種5個重復,每個重復采集0.1~0.2 g樣品。
1.3 苔蘚植物的生活型劃分
依據Bates (1998) 對苔蘚植物生活型的定義, 結合參考張晉昆(1985)、馬文章(2009)對哀牢山苔蘚植物生活型的劃分, 我們將哀牢山徐家壩地區山地森林中苔蘚植物劃分為8類生活型(表2)。
1.4 失水曲線測定
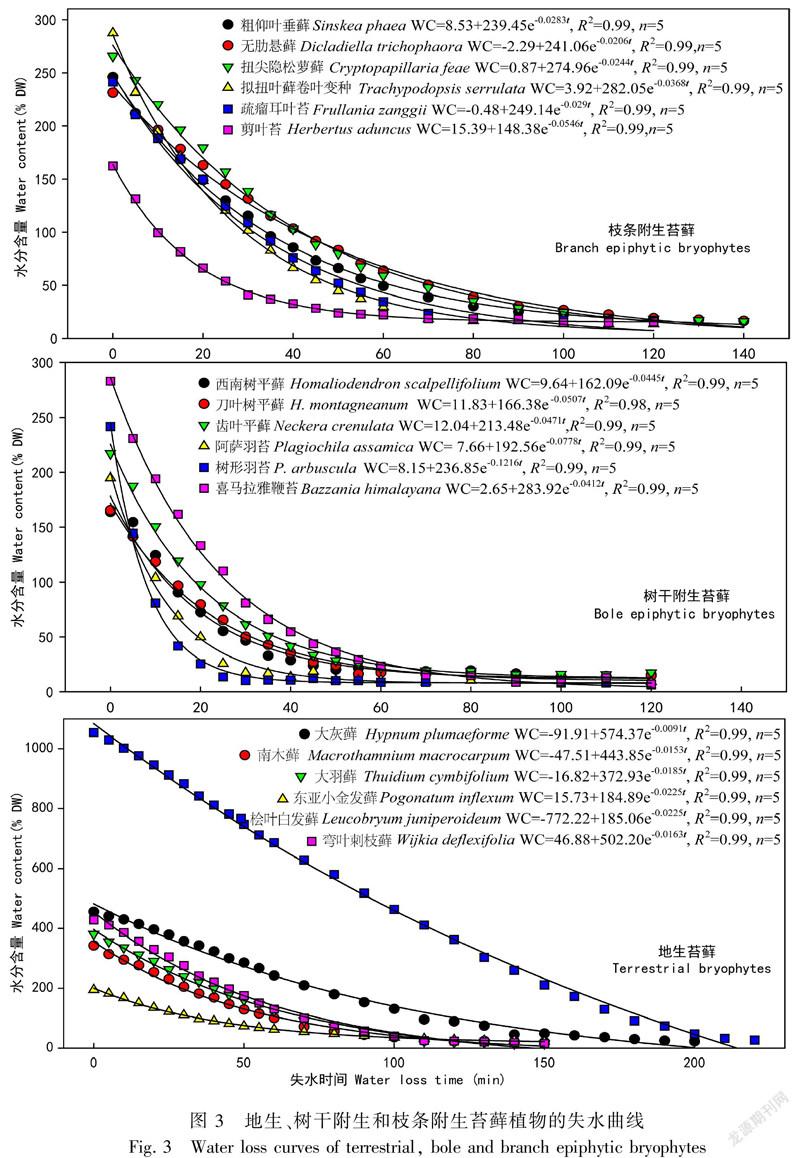
2017年3—4月,進行苔蘚水分特性的觀測試驗。每種苔蘚各選擇5個樣品,對扇型苔蘚將苔蘚莖頂端長2.0~4.5 cm剪下,每份樣品選取2~3個植物體。對其他生活型苔蘚植物小心清除苔蘚樣品上的雜物及枯黃植物體部分,每份樣品選擇3~8個植物體。將樣品在去離子水中浸沒90 min吸水達到完全飽和后,之后取出樣品,吸干樣品表面多余水分(Proctor, 2009);用分析天平(AL204-IC; Mettler Toledo Instruments Ltd., Shanghai, China)稱重(精確至0.001 g);將樣品靜置于溫度約15 ℃、光照約30 μmol·m-2·s-1的恒定環境中,讓其自然失水,前60 min內每間隔5 min對樣品進行稱重。失水60 min后,每間隔10 min對樣品進行稱重至重量不再變化。將所有的樣品在65 ℃下經烘箱烘干處理48 h后稱量得干重(DW)。采用指數消減方程擬合失水進程:
WC=y0+a·e-bt。
式中,WC是樣品含水量;t是蒸發時間;y0、a、b是回歸系數。以浸泡90 min的苔蘚樣品的初始最大絕對含水量與干重的比值為飽和持水率,失水到趨于恒重時的絕對含水量與干重的比值為自然持水率,失水至恒定時所需最短時間為T拐點,平均失水速率=(飽和持水率-自然持水率)/T拐點。
1.5 水分利用效率測定
利用便攜式光合測定系統(LI-6400XT, Li-Cor Inc., Lincoln, NE, USA),由苔蘚葉室(LI-6400-24)配合RGB光源(LI-6400-18)對不同生境苔蘚植物樣品的光合速率和蒸騰速率進行測定。光量子通量密度為250 μmol·m-2·s-1,將從不同生境采集的苔蘚樣品用去離子水浸泡90 min以使其吸水飽和,去除樣品表面過多水分(Zotz et al., 1997),測定過程中空氣流速控制在300 μmol·s-1。測定苔蘚樣品的凈光合速率(Pn)和蒸騰速率(Tr),水分利用效率(WUE)通過Pn與Tr的比值(Pn/Tr)計算求得。葉片投影面積由愛普生平板掃描儀(Perfection V700, Seiko Epson Corp., Nagano, Japan)掃描得到圖像后,利用根系分析系統(WinRHIZO, Pro,2009b, Regend Instruments, Quebec, Canada)分析確定面積從而計算光合速率。2017年3—4月對樣品進行測定。
1.6 數據處理與分析
采用SPSS 20.0 (SPSS, Inc., Chicago, USA)對所獲得數據進行差異顯著性檢驗分析, 并使用SigmaPlot 12.5(Systat Software, Inc., San Jose, USA)和Excel 2010進行相關的圖例繪制。
2 結果與分析
2.1 地生苔蘚、林下樹干和林冠層樹枝附生苔蘚植物的生活型組成
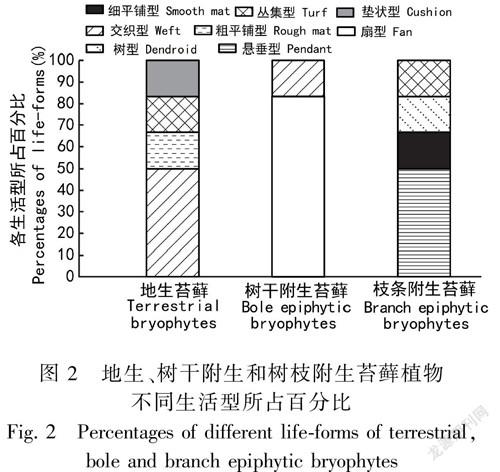
根據野外調查的結果,林地苔蘚、林下樹干及林冠上層樹枝附生苔蘚植物涵蓋了8種苔蘚植物生活型,在所調查的三類不同生境及生長基質上苔蘚植物的生活型組成存在明顯的不同(表2)。三類不同生境下18種苔蘚物種涵蓋13科11屬,其中地生苔蘚植物中有3種為交織型苔蘚,叢集型、墊狀型、粗平鋪型各1種;樹干附生苔蘚中5種為扇型,1種為交織型苔蘚;樹枝附生苔蘚中3種為懸垂型,另外三種分別為樹形、細平鋪型和叢集型。從林地向上到樹干再到林冠上層樹枝,隨著光強、溫度的增加,苔蘚植物的生活型亦隨著生境改變而改變。交織型苔蘚植物占地生苔蘚植物的50%;扇型苔蘚植物占樹干附生苔蘚植物的83%;而在較高層、光照條件較好的樹枝生境下,懸垂型苔蘚植物占據50%(表2,圖1)
2.2 不同生境苔蘚的水分變化特征與水分利用效率
2.2.1 不同生境下苔蘚植物的持水力與失水特征在失水試驗開始的1 h內,所有苔蘚植物的失水速度都很快,以后隨著時間的延長而趨緩,失水曲線均呈現出“J型”下降的趨勢,指數消減方程可以很好地擬合失水曲線變化,其決定系數(R2)均大于0.98(圖3)。除了地生苔蘚,林冠層樹枝附生苔蘚粗仰葉垂蘚、無肋懸蘚之外,其它苔蘚植物均在2 h內失去大多數水分,其后樣品基本處于恒重狀態。
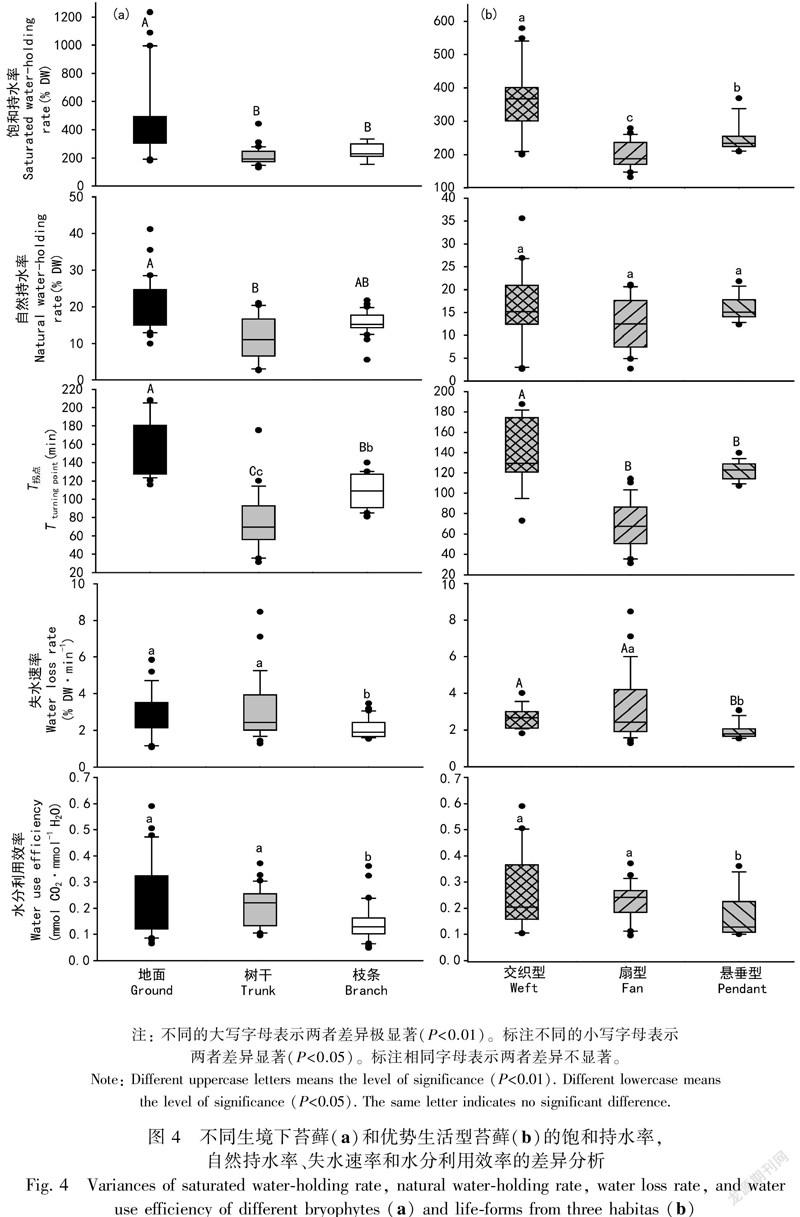
通過對不同生境下苔蘚植物的持水力與失水特征測定結果表明,地生苔蘚植物的自然持水率、飽和持水率均顯著高于附生苔蘚植物,它們之間的差異達到極顯著水平(P<0.01),但樹干附生苔蘚與樹枝附生苔蘚之間自然持水率和飽和持水率的差異不顯著。失水至恒定時所需最短時間(T拐點)表現為地生苔蘚>樹枝附生苔蘚>樹干附生苔蘚,兩兩之間存在顯著差異。地生、樹干附生苔蘚的平均失水速率均大于樹枝附生苔蘚,其差異達到了顯著水平,而地生苔蘚與樹干附生苔蘚的平均失水速率差異不顯著(圖4:a)。
2.2.2 不同生活型苔蘚植物的持水力與失水特征
苔蘚植物生活型是適應環境的表現, 是苔蘚植物的生態策略(Bates, 1998)。前面的研究結果顯示,交織型、扇型、懸垂型分別是地生苔蘚、林下樹干附生和林冠層樹枝附生苔蘚植物的優勢生活型,通過對這三種生活型的持水力和失水特征的測定結果表明,交織型苔蘚的飽和持水率大于懸垂型苔蘚,而懸垂型苔蘚的飽和持水率又大于扇型苔蘚,且兩兩之間的差異均達到顯著性水平,表明了交織型苔蘚相較于扇型和懸垂型苔蘚具有較高的持水能力,而三種生活型之間的自然持水率則無顯著差異。
苔蘚植物是變水植物,在自然干燥條件下,1 h內喪失其大部分水分。不同生活型苔蘚植物的失水特性有所不同,測定結果表明,失水至恒定時所需最短時間(T拐點)表現為交織型>懸垂型>扇型,雖然交織型苔蘚與懸垂型苔蘚之間差異不顯著,但兩者與扇型苔蘚間的差異均達到了極顯著水平(P<0.01)。交織型苔蘚與扇型苔蘚的平均失水速率均大于懸垂型苔蘚,其差異達到極顯著水平(P<0.01),而交織型苔蘚和扇型苔蘚間無顯著差異。
交織型和扇型苔蘚植物的失水速率均較快。懸垂型苔蘚植物的持水率居于中間水平,而失水進程較長,失水速率較慢,表明了懸垂型苔蘚具有較好的保水性能。而扇型苔蘚持水率最小,失水時間較短,失水速率較快,表明其持水能力和保水性能均較差(圖 4:b)。
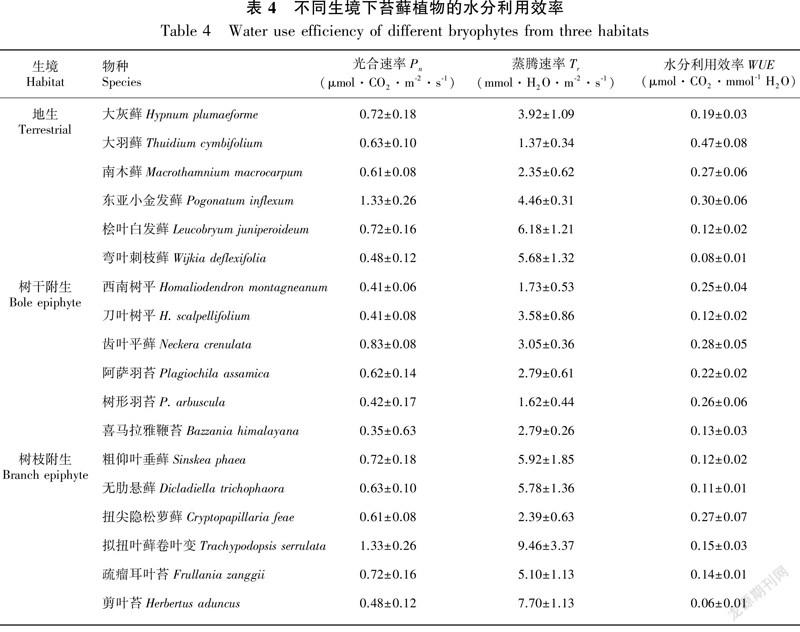
2.2.3 不同生境和不同生活型苔蘚的水分利用效率測定結果表明,地生、林下樹干附生和林冠層樹枝附生三種生境下苔蘚植物的水分利用效率分別為地生苔蘚(0.22±0.14)、樹干附生苔蘚(0.19±0.09)、樹枝附生苔蘚(0.14±0.08)。差異顯著性檢驗結果顯示地生苔蘚的水分利用效率大于樹干附生苔蘚,但它們之間的差異不顯著。而地生苔蘚和樹干附生苔蘚的水分利用效率顯著高于樹枝附生苔蘚(P<0.01)(圖4:a)。
根據對交織型、扇型和懸垂型三種優勢苔蘚植物生活型的水分利用效率的測定結果,表明三種生活型苔蘚植物的水分利用效率分別為交織型(0.26±0.14)、扇型(0.22±0.07)、懸垂型(0.17±0.09)。差異顯著性結果表明:交織型苔蘚與扇型苔蘚之間差異不顯著,而這兩種生活型苔蘚植物的水分利用效率均顯著高于在高層樹枝生境占據優勢的懸垂型苔蘚(P<0.01)(圖4:b)。
3 討論與結論
3.1 不同生境與不同生活型苔蘚植物的水分變化特征
在森林內不同垂直梯度上環境因子存在差異,隨著高度的增加,光照、溫度以及植物蒸散逐漸增高,而空氣濕度則逐漸減低(本研究圖1;Song et al.,2015;Peter et al.,2016)。不同生境條件下不同苔蘚植物在其水分變化特征上有所不同。在本研究中,地生苔蘚植物飽和持水率,自然持水率較大,具有較強的持水能力,其失水進程耗時較長,但失水速度較快; 樹干附生苔蘚的飽和持水率這與樹枝附生苔蘚長期適應其低濕、較高風速與蒸散速率的生境條件而形成水分適應策略有密切關系。苔蘚植物多為變水植物,它們缺乏角質層和維管束組織,沒有真正的根系,對自身水分控制能力不足,其含水量易隨環境變化而變化(劉文耀等,2006)。苔蘚植物往往會因環境變干而進入休眠,當外界環境條件變濕后,又能迅速復水恢復正常的新陳代謝功能(Proctor et al., 2007)。相對于地生苔蘚和林下樹干附生苔蘚植物而言,林冠層樹枝附生苔蘚植物面臨更為頻繁的干濕交替,一旦遇到降雨或者云霧天氣,其較高的保水性能可以幫助樹枝附生苔蘚在更長的時間內保持較適的含水量從而維持正常的生理活動以適應林冠層嚴酷的生境條件。
苔蘚植物的生活型是其對生境條件的一種適應表現,苔蘚植物的生活型組成隨著生境的改變而改變。通過分析不同生境下苔蘚植物的生活型組成與特征,能揭示苔蘚植物與環境之間的關系(Kürschner, 2004)。一般來說,矮小、硬挺、叢集密生或交織匍匐生長,呈叢狀、墊狀或毯狀生活型的苔蘚植物能夠有效降低葉子和大氣的接觸面積,從而降低大氣在葉子空隙之間的流動,因而能夠有效地減少水分蒸發,有利于提高毛細管系統的持水能力,因此耐旱能力較強(吳鵬程,1998)。在本研究的不同生活型苔蘚中,交織型苔蘚顯示出較高的持水能力,墊狀型的檜葉白發蘚、粗平鋪型的彎葉刺枝蘚均具有較高的持水能力,而叢集型的東亞小金發蘚的持水能力較小,但保水性較好。扇型苔蘚是哀牢山地區山地森林樹干附生苔蘚植物中占優勢的生活型(Ma et al., 2009),其具有較大的表面積和體積比,有利于它們與大氣充分接觸并從中截獲水分,但較大的表面積也使其對大氣濕度變化敏感,具有較差的耐受性(Bates, 1998; Kürschner et al., 1999)。扇型和懸垂型苔蘚的水分來源主要依靠降雨截留和空氣中的水分,這兩種生活型苔蘚在熱帶,亞熱帶山地云霧林中較為常見(León-Vargas, 2006)。在亞熱帶、熱帶的山地云霧林區域,降雨和強風事件較為頻繁,林冠層樹枝附生的懸垂型苔蘚具有較低的持水能力,以適應多雨、多風的環境條件,因為吸持過多的水分易被風吹落或者因自身重力作用脫離原來的生境(Proctor, 2004)。
3.2 不同生境與不同生活型苔蘚植物的水分利用效率
植物的水分利用效率(water use efficiency,WUE)指的是蒸發單位水所產生的干物質的量(Fischer,1979),能夠解釋植物內在的耗水機制,顯示植物有效利用水分的能力(曹生奎等,2009)。這里的水分利用效率指的是葉片水平上的瞬時水分利用效率,是生物量積累與蒸騰耗水之比(Farquhar & Richards,1984)。蔣高明和何維明(1999)對內蒙古毛烏素沙地不同生境下66種草本植物的水分利用效率的研究結果,表明其WUE范圍為0.19~4.04 μmol CO2·mmoL<sup>-1</sup>H2O;中國東部南北樣帶森林優勢木本植物的水分利用效率范圍在8.86~10.97 μmol CO2·mmoL<sup>-1</sup>H2O之間(展小云等,2012)。與維管束植物相比,苔蘚植物的水分利用效率普遍較低。本研究中不同生境條件下18種苔蘚植物的水分利用效率(WUE)的范圍為0.06~0.47 μmol CO2·mmoL<sup>-1</sup>H2O,顯然,哀牢山地區山地森林中苔蘚植物的WUE要比其他地區的維管束植物的低。在較干旱的生境下,較高的WUE和保守的水分利用策略有利用植物的生長、生存和維持。苔蘚植物較低的WUE和揮霍的水分利用模式可以使其獲得較高的生產力(曹生奎等,2009)。在哀牢山徐家壩地區,降雨充沛,常年氣候濕潤,雨季苔蘚植物基本上處于水分飽和的狀態下,這可能是苔蘚植物具有較低的水分利用效率的原因之一。
隨著生境條件由濕到干的不同水分供應條件,毛烏素沙地的草本植物的水分利用效率呈現升高的趨勢(蔣高明等,1999)。在哥斯達黎加的熱帶雨林中,附生蕨比地生蕨具有更高的WUE(Watkins et al., 2007);分布在冠層上部的附生維管束植物通過降低蒸騰速率,提高WUE,以適應相對干燥高溫的微生境(江浩等,2012)。但在本研究中,地生苔蘚和樹干附生苔蘚的水分利用效率反而高于樹枝附生苔蘚,這與苔蘚植物自身的生物學特性以及所處生境條件有關。苔蘚植物的葉片沒有氣孔的分化,缺乏對水分和CO2的控制,蒸散阻力主要取決于苔蘚植物的葉肉導度(Waite & Sack,2011)。姬明飛和韓鴻基(2017)對寶天曼自然保護區內的兩種優勢苔蘚的研究表明大羽蘚(Thuidium cymbifolium)的WUE高于無邊提燈蘚(Mnium immarginatum),他們將其歸因于兩種苔蘚植物“氣孔導度”(葉肉導度)的差異。當空氣濕度或水分可利用性降低時,苔蘚植物無法調節氣孔關閉來減弱蒸騰作用,因此遭受了較高的蒸發要求,導致植物葉片的WUE隨水分可利用性降低而降低(劉文兆,1998;曹生奎等,2009)。林冠層樹枝生境的溫度較高、風速較大,而空氣濕度相對較低,在這樣生境下植物的蒸發較大,從而導致林冠層樹枝附生苔蘚植物的WUE低于地生苔蘚和林下樹干附生苔蘚植物,尤其是林冠層樹枝懸垂型附生苔蘚植物的WUE顯著低于林下地生交織型苔蘚和樹干扇型附生苔蘚。
致謝 感謝哀牢山亞熱帶森林生態系統研究站提供氣象觀測數據,同時感謝站內工作人員羅琦,杞金華等人在野外工作中給予的幫助。
參考文獻:
BAO WK, WANG CM, WU N, 2004. Change of water holding and conservation capacity of eight species of bryophytes under subalpine coniferous forests in Eastern Tibetan Plateau [J]. J Nat Resour,19(2): 190-194.[包維楷, 王春明, 吳寧, 2004. 青藏高原東部針葉林下8種蘚類植物的持水和保水能力比較研究 [J]. 自然資源學報, 19(2): 190-194.]
BATES JW, 1998. Is ‘life-form’ a useful concept in bryophyte ecology? [J]. Oikos, 82(2): 223-237.
CAO SK, FENG Q, SI JH, et al., 2009. Summary of the plant water use efficiency at leave level [J]. Acta Ecol Sin, 29(7): 3882-3892.[曹生奎, 馮起, 司建華, 等, 2009. 植物葉片水分利用效率研究綜述 [J]. 生態學報, 29(7): 3882-3892.]
CHE ZX, LIU XD, JING WM, et al., 2006. Vertical distribution characteristics and hydrological function analysis of bryophytes in Qilian Mountain area [J]. J Soil Water Conserv, 20(6): 71-74.[車宗璽, 劉賢德, 敬文茂, 等, 2006. 祁連山林區苔蘚垂直分布特征與水文功能分析 [J]. 水土保持學報, 20(6): 71-74.]
FARQUHAR GD, RICHARDS RA, 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes [J]. Funct Plant Biol, 11(6): 539-552.
FARRANT JM, MOORE JP, 2011. Programming desiccation-tolerance: From plants to seeds to resurrection plants [J]. Curr Opin Plant Biol, 14(3): 340-345.
FISCHER RA, 1979. Growth and water limitation to dryland wheat yield in Australia: A physiological framework [J]. J Aust Inst Agric Sci, 45(2): 83-94.
GUAN FR, RU YL, HU ZJ, et al., 2017. Host specificity of epiphytic bryophytes in Hangzhou [J]. Chin J Ecol, 36(1): 61-66.[官飛榮,茹雅璐,胡忠健,等, 2017. 杭州市樹附生苔蘚植物的附生專一性 [J]. 生態學雜志, 36(1): 61-66.]
GUO SL, HAN SJ, CAO T, 1999. Indicative value of bryophytes on forest eco-boundary [J]. Chin J Appl Ecol, 10(1): 1-6.[郭水良, 韓士杰, 曹同, 1999. 苔蘚植物對森林生態界面指示作用的研究 [J]. 應用生態學報, 10(1):1-6.]
HUMPHREY JW, DAVEY S, PEACE AJ, et al., 2002. Lichens and bryophyte communities of planted and semi-natural forests in Britain: The influence of site type, stand structure and deadwood [J]. Biol Conserv, 170(2): 165-180.
JIANG GM, HE WM, 1999. Species and habitat variability of photosynthesis, transpiration and water use efficiency of different plant species in Maowusu sand area [J]. Acta Bot Sin, 41(10): 1114-1124.[蔣高明, 何維明, 1999. 毛烏素沙地若干植物光合作用、蒸騰作用和水分利用效率種間及生境間差異 [J]. 植物學報(英文版), 41(10): 1114-1124.]
JIANG H, HUANG YH, ZHOU GY, et al., 2012. Acclimation in leaf morphological and eco-physiological characteristics of different canopy-dwelling epiphytes in a lower subtropical evergreen broad-leaved forest [J]. Plant Sci J, 30(3): 250-260.[江浩, 黃鈺輝, 周國逸, 等, 2012. 亞熱帶常綠闊葉林冠層附生植物葉片形態結構及生理功能特征的適應性研究 [J]. 植物科學學報, 30(3): 250-260.]
JI MF, HAN HJ, 2017.The study of photosynthetic characteristics of two moss species under different light condition [J]. Pratac Sci, 34(9): 1787-1792.[姬明飛, 韓鴻基, 2017. 兩種蘚類植物對光強的適應性差異 [J]. 草業科學, 34(9): 1787-1792.]
KRUYS N, JONSSON BG, 1999. Fine woody debris is important for species richness on logs in managed boreal spruce forests for northern Sweden [J]. Can J For Res, 29(8):1295-1299.
KURSCHNER H, FREY W, PAROLLY G, 1999. Patterns and adaptive trends of life forms, life strategies and ecomorphological structures in tropical epiphytic bryophytes a pantropical synopsis [J]. Nova Hedwigia, 69(1-2):73-99.
KURSCHNER H, 2004. Life strategies and adaptations in bryophytes from the Near and Middle East [J]. Turk J Bot, 28(1-2):73-84.
LEO′N-VARGAS Y, ENGWALD S, PROCTOR MCF, 2006. Microclimate, light adaptation and desiccation tolerance of epiphytic bryophytes in two venezuelan cloud forests [J]. J Biogeog, 33(5): 901-913.
LI S, LIU WY, WANG LS, et al., 2007. Species diversity and distribution of epiphytic lichens in the primary and secondary forests in Ailao Mountain, Yunnan [J]. Biodivers Sci, 15(5): 445-455.[李蘇, 劉文耀, 王立松, 等, 2007. 云南哀牢山原生林及次生林群落附生地衣物種多樣性與分布 [J]. 生物多樣性, 15(5): 445-455.]
LIU WY, MA WZ, YANG LP, 2006. Advances in ecological studies on epiphytes in forest canopies [J]. J Plant Ecol, 30(3): 522-533.[劉文耀, 馬文章, 楊禮攀, 2006. 林冠附生植物生態學研究進展 [J]. 植物生態學報, 30(3):522-533.]
LIU WY, FOX JED, XU ZF, 2002. Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane subtropical moist forest on Ailao Mountains in Yunnan, South-West China [J]. J Trop Ecol, 18(4): 527-548.
LIU WZ, 1998. Dynamic interrelations of crop production, water consumption and water use efficiency [J]. J Nat Resour, 23(1): 23-27.[劉文兆, 1998. 作物生產,水分消耗與水分利用效率間的動態聯系 [J]. 自然資源學報, 23(1):23-27.]
LIU YH, 1993. An analysis of the climatic resource of ground temperature in the north part of the Ailao Mountains Yunnan Province [J]. J Nat Resour, 8(2): 158-165.[劉玉洪,1993. 哀牢山北段山地的低溫氣候資源分析 [J]. 自然資源學報, 8(2): 158-165 ]
MA WZ,LIU WY, SONG L, 2011. Species composition and distribution of bryophytes on different substrate types in middle mountain moist evergreen broad-leaved forest in Ailao Mountains, Yunnan [J]. Plant Divers Resour, 33(4): 443-450.[馬文章, 劉文耀, 宋亮, 2011. 哀牢山中山濕性常綠闊葉林不同生長基質苔蘚植物的組成與分布特征 [J]. 植物分類與資源學報, 33(4): 443-450]
MA WZ, 2009. The composition and biomass of epiphytic materials and their relationships with ecological factors in Xujiaba region from Ailao Mountain, Yunnan [D]. Beijing: Graduate school of the Chinese Academy of Sciences: 22-23.[馬文章, 2009. 云南哀牢山徐家壩地區附生(植)物的組成、生物量及其與生態因子的關系 [D]. 北京:中國科學院研究生院:22-23.]
MALSON K, RYDIN H, 2009. Competitive hierarchy, but no competitive exclusions in experiments with rich fen bryophytes [J]. J Bryol, 31(1): 41-45.
PETTER G, WAGNER K, WANEK W, et al., 2016. Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra-and interspecific trait variability, and taxonomic signals [J]. Funct Ecol, 30(2): 188-198.
PROCTOR MCF, 2004. Light and desiccation responses of Weymouthia mollis and W. cochlearifolia, two pendulous rainforest epiphytes from Australia and New Zealand [J]. J Bryol, 26(3): 167-173.
PROCTOR MCF, 2009. Physiology ecology: Bryophyte biology [M]. GOFFINET B, SHAW AJ. Cambridge: Cambridge University: 237-269.
PROCTOR MCF, OLIVER MJ, WOOD AJ, et al., 2007. De-siccation-tolerance in bryophytes: A review [J]. Bryologist, 110(4):595-621.
QIU XZ, XIE SC, 1998. Studies on the forest ecosystem in Ailao Mountains Yunnan, China [M]. Kunming: Yunnan Science & Technology Press: 7-11.[邱學忠, 謝壽昌, 1998. 云南哀牢山森林生態系統研究 [M]. 昆明: 云南科技出版社: 7-11.]
SCHOFIELD WB, 1985. Introduction to bryology [M]. New York: Macmillan Press: 316-319.
SMITH AJE, 1982. Bryophyte ecology [M]. London: Springer: 333-382.
SONG L, ZHANG YJ, CHEN X, et al., 2015. Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest [J]. J Plant Res, 128(4): 573-584.
WATKINS JE, RUNDEL PW, CARDELUS CL, 2007. The influence of life form on carbon and nitrogen relationships in tropical rainforest ferns [J]. Oecologia, 153(2): 225-232.
WAITE M, SACK L, 2011. Shifts in bryophyte carbon isotope ratio across an elevation × soil age matrix on Mauna Loa, Hawaii: Do bryophytes behave like vascular plants? [J]. Oecologia, 166(1): 11-22.
WU PC, 1998. Bryological biology,introduction and diverse branches [M]. Beijing: Science Press: 131-143.[吳鵬程,1998. 苔蘚植物生物學 [M]. 北京: 科學出版社: 131-143.]
XU HQ, LIU WY, 2005. Water interception and monitoring for environment changing of epiphytes [J]. Res Soil Water Conserv, 12(2): 116-120.[徐海清, 劉文耀, 2005. 林冠附生物對水分的截留及對環境的監測 [J]. 水土保持研究, 12(2): 116-120.]
YOU CX, 1983. Classification of vegetation in Xujiaba region in Ailao Mts. Research of forest ecosystems on Ailao Mountains Yunnan [M]. Kunming: Yunnan Science & Technology Press: 74-117.[游承俠, 1983. 哀牢山徐家壩地區的植被分類. 云南哀牢山森林生態系統研究 [M]. 昆明: 云南科技出版社: 74-117.]
ZHAN XY, YU GR, SHENG WP, et al., 2012. Foliar water use efficiency and nitrogen use efficiency of dominant plant species in main forests along the North-South Transect of East China [J]. Chin J Appl Ecol, 23(3): 587-594.[展小云, 于貴瑞, 盛文萍, 等, 2012. 中國東部南北樣帶森林優勢植物葉片的水分利用效率和氮素利用效率 [J]. 應用生態學報, 23(3): 587-594.]
ZHANG HJ, CHENG JH, SHI YH, et al., 2003. Water conserving function of moss on forest stands in the Three Gorges Area [J]. Res Environ Yangtze Basin, 12(5): 457-461.[張洪江,程金花,史玉虎,等, 2003. 三峽庫區幾種林下苔蘚的保水功能 [J]. 長江流域資源與環境, 12(5): 457-461.]
ZHANG XQ, ZENG JJ, CHEN JW, et al., 2012. The saxicolous mosss features of absorbing water and its structural adaptabilityin the heterogeneous environment with rock desertification [J]. Acta Ecol Sin,32(12): 3902-3911.[張顯強,曾建軍,諶金吾,等,2012. 石漠化干旱環境中石生蘚類水分吸收特征及其結構適應性 [J]. 生態學報, 32(12):3902-3911.]
ZHANG JK, 1985. A preliminary study of bryophytes in the moist evergreen broad-leaved forest in the Xujiaba area of Ailao Mts. Yunnan [D]. Kunming: Yunnan University.[張晉昆, 1985. 云南哀牢山徐家壩地區濕性常綠闊葉林苔蘚植物的初步研究 [D]. 昆明: 云南大學.]
ZOTZ G, BUDE B, MEYER A, et al., 1997. Water relations and CO2exchange of tropical bryophytes in a lower montane rain forest in Panama [J]. Plant Biol, 110(1): 9-17.
猜你喜歡
中國校外教育(中旬)(2017年13期)2018-03-26 08:29:38
熱帶農業科學(2017年9期)2017-10-23 23:06:56
成才之路(2017年20期)2017-08-02 21:28:40
成才之路(2017年21期)2017-07-27 20:52:40
安徽農學通報(2017年9期)2017-05-19 23:03:05
環境與發展(2017年2期)2017-05-13 20:42:53
現代農業科技(2016年22期)2017-03-24 12:21:57
湖北農業科學(2015年14期)2015-09-09 09:17:06
學周刊·下旬刊(2015年10期)2015-07-22 12:36:17
湖北農業科學(2015年2期)2015-03-20 13:47:36
