子癇前期胎盤組織miRNA差異表達譜生物學分析及miR-365a-3p參與發病的機制探討
2020-04-29 12:00:40李暉焦順鄭曉丹肖黎陳智芳劉榮
山東醫藥 2020年11期
李暉,焦順,鄭曉丹,肖黎,陳智芳,劉榮
荊州市中心醫院,湖北荊州434020
子癇前期是一種在妊娠期特發的全身性疾病,其臨床表現為妊娠末3個月、分娩過程中或分娩后不久出現高血壓、蛋白尿及其他系統性疾病,全世界發病率為3%~8%[1,2]。子癇前期的確切病因和發病機制仍不明確,但已有研究證實子癇前期是由遺傳因素和非遺傳因素相互作用導致的[3]。目前研究認為,滋養細胞侵襲能力減弱所致“螺旋動脈重塑障礙”、“胎盤淺著床”是子癇前期發病的關鍵環節[4]。miRNA具有基因表達調節劑的作用,具有調控多種細胞侵襲、遷移的功能[5~7]。因此,我們猜測子癇前期患者胎盤組織中可能存在某種或某些異常表達的miRNA,這些miRNA可能在子癇前期的發生發展中發揮重要作用。2017年1月~2018年12月,本研究對子癇前期患者胎盤組織miRNA表達譜進行鑒定與功能分析,對差異表達miR-365a-3p參與子癇前期發病的機制進行探討。現報告如下。
1 資料與方法
1.1 臨床資料 選取同期荊州市中心醫院婦產科行剖宮產分娩的重度子癇前期患者以及正常孕婦各41例。重度子癇前期的診斷標準:妊娠20周后新發高血壓,且收縮壓≥160 mmHg或舒張壓≥110 mmHg;或者伴有明顯的蛋白尿;或出現其他器官受累癥狀,如肺水腫、肝功異常,尤其是肝酶異常、持續性上腹痛、進行性腎損傷、視物模糊或頭痛等癥狀。重度子癇前期患者年齡(30.25±1.12)歲,分娩時孕周(36.89±0.61)周,BMI(27.83±0.73)kg/m2;正常孕婦年齡(29.37±0.89)歲,分娩時孕周(39.32±0.22)周,BMI(27.21±0.88)kg/m2。重度子癇前期患者與正常孕婦的一般資料均具有可比性。本研究通過醫院倫理委員會審核,患者及其家屬均簽署知情同意書。
1.2 標本處理 胎盤經剖宮產分娩方式娩出1 h內完成組織采樣。以臍帶附著位點為中心,將胎盤分為四個象限;在胎盤邊緣與臍帶附著位點中間、母體面與胎兒面間,避免明顯的鈣化點、大血管及梗死灶;各象限隨機采樣,樣本大小約8 cm3,采樣后迅速置于液氮中保存。
1.3 子癇前期胎盤組織miRNA差異表達譜的建立及靶基因功能富集分析
1.3.1 miRNA差異表達譜建立 采用高通量測序技術。隨機選取子癇前期及正常胎盤組織各3例份,充分研磨后TRIzol法提取總RNA。使用Small RNA Sample Pre Kit構建文庫,利用Small RNA的5′及3′端特殊結構(5′端有完整的磷酸基團、3′端有羥基),以total RNA為起始樣品,將Small RNA兩端加上接頭,反轉錄合成cDNA。經過PCR擴增,PAGE膠電泳分離目標DNA片段,切膠回收得到cDNA文庫。文庫構建完成后,使用Qubit2.0進行初步定量,稀釋文庫至1 ng/μL;隨后使用Agilent 2100對文庫的insert size進行檢測,insert size符合預期后,采用Q-PCR法對文庫的有效濃度進行準確定量(文庫有效濃度>2 nmol/L),以保證文庫質量。庫檢合格后,把不同文庫按照有效濃度及目標下機數據量的需求混合后進行HiSeq測序。對比分析子癇前期與正常胎盤組織中的差異表達miRNA,以差異倍數≥2且P<0.05作為顯著上調和顯著下調的標準。
1.3.2 差異表達miRNA的靶基因功能富集分析 通過TargetScan、PITA、miRBase、microRA.org、miRTarBase、starBase等軟件針對差異表達miRNA預測可能作用的靶基因,取交集共獲得1 983個潛在靶基因。對差異表達miRNA潛在的靶基因進行GO和KEGG功能富集分析。GO富集分析方法:將基因向GO數據庫(http://www.geneontology.org/)的各term映射,并計算每個term的基因數,從而得到具有某個GO功能的基因列表及基因數目;應用超幾何檢驗,找出與整個基因組背景相比,在基因中顯著富集的GO條目。KEGG富集分析方法:以KEGG Pathway為單位,應用超幾何檢驗,找出與整個基因組背景相比,在基因中顯著富集的Pathway;通過Pathway顯著富集確定基因參與的最主要的生化代謝途徑和信號轉導途徑。
1.4 子癇前期與正常胎盤組織miR-365a-3p表達檢測 采用real-time PCR法。選取子癇前期及正常胎盤組織各41例份,充分研磨后采用TRIzol法提取總RNA,逆轉錄合成cDNA。PCR反應體系共20 μL:2×miRNA Universal SYBR qPCR Master Mix 10 μL,正向引物(10 μmol/L)0.4 μL,mQ Primer R(10 μmol/L)0.4 μL,cDNA模板100 ng,加入RNase-free ddH2O補充至20 μL。PCR反應條件:95 ℃預變性5 min,95 ℃變性10 s、60 ℃退火30 s,共40個循環。實驗重復3次,取平均值。以U6為內參,采用2-ΔΔCt法計算miR-365a-3p相對表達量。
1.5 miR-365a-3p與TGF-β通路關鍵因子SMAD2、MAPK1的相互作用分析 通過可視化軟件StarBase 3.0(http://starbase.sysu.edu.cn/index.php)預測miR-365a-3p的潛在靶基因。結果顯示,miR-365a-3p與SMAD2、MAPK1的3′非編碼區均存在靶向結合位點,證實miR-365a-3p的潛在靶基因為SMAD2、MAPK1。見圖1。

圖1 miR-365a-3p與SMAD2、MAPK1的結合位點預測結果
1.6 子癇前期與正常胎盤組織SMAD2、MAPK1 mRNA表達檢測 采用real-time PCR法。選取子癇前期及正常胎盤組織各41例份,充分研磨后采用TRIzol法提取組織總RNA,逆轉錄為cDNA。SMAD2上游引物5′-TGTTTTCAGTTCCGCCTCCA-3′,下游引物5′-AGCCTCTTGTATCGAACCTGC-3′;MAPK1上游引物5′-TGACCTCAAGCCTTCCAACC-3′,下游引物5′-GGACTTGGTGTAGCCCTTGG-3′;GAPDH上游引物5′- TCGGAGTCAACGGATTTGGT-3′,下游引物5′- TTCCCGTTCTCAGCCTTGAC-3′。PCR反應體系共25 μL:SYBR Premix Ex Taq 12.5 μL,上下游引物各1 μL,cDNA模板2 μL,ddH2O 8.5 μL;反應條件:95 ℃預變性30 s,95 ℃變性5 s、60 ℃退火20 s,共40個循環。實驗重復3次,取平均值。以GAPDH為內參,采用2-ΔΔCt法計算SMAD2、MAPK1 mRNA相對表達量。
2 結果
2.1 子癇前期胎盤組織中miRNA差異表達情況 與正常胎盤組織相比,子癇前期胎盤組織中有44個miRNA差異表達,其中12個miRNA表達顯著上調、32個miRNA表達顯著下調。前10位表達上調的miRNA分別為miR-365a-5p、miR-193b-5p、miR-210-5p、miR-3196、miR-629-3p、miR-210-3p、miR-193b-3p、miR-6089、miR-365a-3p、miR-365b-3p,差異倍數分別為4.13、3.81、3.78、3.42、3.15、3.04、2.81、2.64、2.57、2.36。前10位表達下調的miRNA分別為miR-652-5p、miR-3614-5p、miR-135b-5p、miR-545-5p、miR-548o-5p、miR-584-3p、miR-373-5p、miR-6513-5p、miR-296-3p、miR-877-5p,差異倍數分別為6.88、6.46、5.75、5.63、5.21、5.04、4.73、4.56、4.42、3.95。miR-365a-3p是差異表達譜中表達上調最顯著的miRNA,選擇miR-365a-3p作為后續研究對象。
2.2 差異表達miRNA的靶基因功能富集分析結果 GO富集分析結果表明,差異表達miRNA的靶基因主要參與的分子功能包括DNA結合、轉錄調節活性、鋅離子結合、轉錄阻遏物活性、過渡金屬離子結合、RNA結合、金屬離子結合、陽離子結合、蛋白質絲氨酸/蘇氨酸激酶活性、染色質結合、轉錄因子結合、序列特異性DNA結合、組蛋白甲基轉移酶活性等;參與的細胞組分包括核腔、核質部分、突觸、細胞連接、細胞器、細胞器腔、膜封閉腔、質膜部分、核仁、突觸部分、細胞內非膜細胞器、非膜細胞器、核外圍、核基質;參與的生物過程包括轉錄調控、RNA代謝過程調控、DNA依賴性RNA聚合酶Ⅱ啟動子轉錄調控、眼睛發育、感覺器官發育、轉錄負調控、依賴DNA、磷酸化、磷代謝過程、磷酸鹽代謝過程、mRNA代謝過程。KEGG富集分析結果表明,差異表達miRNA靶基因主要參與的信號通路包括子宮內膜癌相關信號通路、TGF-β信號通路、Wnt信號通路、急性粒細胞白血病相關信號通路、mTOR信號通路、醛固酮調節鈉的重吸收相關信號通路、慢性粒細胞白血病相關信號通路、胰島素信號通路、ErbB信號通路、癌癥發生相關信號通路等。見圖2。
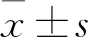
2.3 子癇前期及正常胎盤組織miR-365a-3p及SMAD2、MAPK1 mRNA表達比較 子癇前期及正常胎盤組織miR-365a-3p相對表達量分別為1.264±0.191、0.386±0.084,SMAD2 mRNA相對表達量分別為1.531±0.282、2.658±0.319,MAPK1 mRNA相對表達量分別為1.583±0.294、2.424±0.307;二者比較P均<0.01。
2.4 子癇前期胎盤組織miR-365a -3p與SMAD2、MAPK1 mRNA表達的關系 子癇前期胎盤組織中miR-365a-3p與SMAD2、MAPK1 mRNA相對表達量均呈顯著負相關關系(R2=0.536,R2=0.422,P均<0.05)。見圖3。

注:A、B、C為GO富集分析結果,D為KEGG富集分析結果。
圖2 差異表達miRNA的靶基因GO及KEGG富集分析結果

圖3 miR-365a-3p與SMAD2、MAPK1 mRNA表達的關系
3 討論
特定器官、組織中特異性表達的miRNA在人體發育過程中具有關鍵作用,而鑒定組織特異性miRNA表達譜對探討疾病發生的病理機制至關重要。研究表明,人胎盤組織中多種miRNA與妊娠過程有關,其表達在妊娠過程中可發生大幅波動[8]。Jiang等[9]研究表明,重度子癇前期的孕婦在孕早期血清miR-520g水平升高,并可通過影響MMP-2翻譯來抑制滋養細胞的遷移和侵襲,且miR-520g可能在有缺陷的螺旋動脈重塑中具有重要作用。Adel等[10]發現,與正常妊娠婦女相比,子癇前期患者胎盤組織中miR-210表達上調,且miR-210表達升高可導致其靶基因PTPN2 mRNA下調,miR-210可能參與了子癇前期的發病并與疾病嚴重程度有關。因此,子癇前期胎盤組織中存在某種或某些其他異常表達的miRNA,在子癇前期的發生發展中發揮重要作用。
本研究高通量測序及分析結果顯示,與正常胎盤組織相比,子癇前期胎盤組織中有44個miRNA差異表達,上調miRNA差異倍數前10位為miR-365a-5p、miR-193b-5p、miR-210-5p、miR-3196、miR-629-3p、miR-210-3p、miR-193b-3p、miR-6089、miR-365a-3p、miR-365b-3p,下調miRNA差異倍數前10位為miR-652-5p、miR-3614-5p、miR-135b-5p、miR-545-5p、miR-548o-5p、miR-584-3p、miR-373-5p、miR-6513-5p、miR-296-3p、miR-877-5p。這些差異表達的miRNA均具有參與子癇前期發病及預測指標的潛能。miRNA作為內源性具有調控功能的非編碼RNA,主要通過與靶基因mRNA的3′非翻譯區結合,調控靶基因表達發揮作用[11,12]。因此,本研究通過TargetScan、PITA、miRBase、microRA.org、miRTarBase、starBase等軟件針對差異表達miRNA預測可能作用的靶基因,共獲得1 983個潛在靶基因。對差異表達miRNA的潛在靶基因進行功能富集分析,結果表明其主要參與子宮內膜癌相關信號通路、TGF-β信號通路、Wnt信號通路、急性粒細胞白血病相關信號通路、mTOR信號通路、醛固酮調節鈉的重吸收相關信號通路、慢性粒細胞白血病相關信號通路、胰島素信號通路、ErbB信號通路、癌癥發生相關信號通路等。提示上述異常表達的miRNA或可通過某些通路參與子癇前期的發病。
本研究中miR-365a-3p是差異表達譜中表達上調最顯著的miRNA,因此選擇miR-365a-3p作為后續研究對象。miR-365a-3p在多種疾病中發揮重要作用。Li等[13]研究表明,miR-365a-3p在喉癌中差異表達,在喉癌細胞中抑制miR-365a-3p可以上調TET1表達,干擾TET1可以消除miR-365a-3p對細胞增殖、周期進程和侵襲的影響。Liu等[14]發現,大腦皮質神經元和原代培養神經元中LncRNA GAS5表達明顯高于對照組,其可通過與miR-365a-3p競爭性結合調節DCDC2表達,從而形成GAS5/miR-365a-3p/DCDC2調控網絡,導致大腦皮層神經元凋亡增加。本研究結果顯示,子癇前期胎盤組織中miR-365a-3p表達明顯高于正常胎盤組織,通過可視化軟件StarBase 3.0預測其潛在靶基因為TGF-β通路關鍵基因SMAD2、MAPK1。本研究結果顯示,子癇前期胎盤組織中SMAD2、MAPK1 mRNA表達降低,二者與miR-365a-3p表達呈顯著負相關關系,提示miR-365a-3p可能通過TGF-β通路參與子癇前期的發病。
綜上所述,子癇前期胎盤組織存在異常表達的miRNA,并參與諸多信號通路;子癇前期胎盤組織miR-365a-3p表達升高,其可能通過負性調控TGF-β信號通路參與子癇前期的發病。miR-365a-3p有望成為子癇前期的干預靶點,但仍有待后續細胞和動物實驗進行驗證。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
電子制作(2018年11期)2018-08-04 03:25:42
山東工業技術(2016年15期)2016-12-01 05:31:22
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
