
小麥抗白粉病及分子標記研究進展
2020-07-09 03:47:58周軍徐如宏謝鑫
湖北農業科學 2020年6期
周軍 徐如宏 謝鑫

摘要:小麥白粉病是威脅小麥的主要病害。實踐證明,選育和應用抗病品種是解決小麥白粉病最經濟、安全、有效的措施。近年來,分子標記技術的快速發展為小麥抗白粉病基因的研究奠定了基礎。對小麥白粉病菌的形態特征、毒性頻率、抗性基因來源、染色體定位、生理抗性、抗性基因應用等方面研究進展進行了綜述,旨為小麥抗病育種提供參考依據。由于病原菌生理小種極容易變異,導致克服寄主原有抗性,可通過基因聚合使不同抗性基因實現有效結合,拓寬其抗譜,提高其抗性持久性,對小麥白粉病抗性研究具有重要理論和實踐意義。
關鍵詞:小麥;白粉病;分子標記
中圖分類號:S512.1 ? ? ? ? 文獻標識碼:A
文章編號:0439-8114(2020)06-0010-06
DOI:10.14088/j.cnki.issn0439-8114.2020.06.002 ? ? ? ? ? 開放科學(資源服務)標識碼(OSID):
Research advances on wheat powdery mildew resistance and molecular markers
ZHOU Jun,XU Ru-hong,XIE Xin,REN Ming-jian
(College of Agriculture,Guizhou University/Guizhou Sub-center of National Wheat Center,Guiyang 550025,China)
Abstract: Wheat powdery mildew is the main disease that harms the field of wheat. It has been proved that breeding and application of resistant varieties are the most economical, safe and effective measures to solve wheat powdery mildew. In recent years, the rapid development of molecular marker technology has laid the foundation for the research of resistant genes of wheat powdery mildew. The morphological characteristics, virulence frequency, source of resistance gene and chromosome location, physiological resistance and application of resistance gene of wheat powdery mildew were reviewed, in order to provide reference for wheat disease resistance breeding. Because the physiological races of pathogenic bacteria are extremely susceptible to mutation, which leads to overcoming the host's original resistance, different resistance genes can be effectively combined through gene aggregation, the resistance spectrum can be broadened, and the resistance can be lasted. And it is of great theoretical and practical significance to study the resistance of wheat powdery mildew.
Key words:wheat; powdery mildew; molecular markers
小麥白粉病是由禾本科布氏白粉病菌(Blumeria graminis)引起的氣傳性真菌病害,具有發生強度高、流行范圍廣等特點,嚴重威脅中國小麥生產。世界各主要麥產區均有白粉病發生,產量損失2%~5%;白粉病發生嚴重的歐美地區,小麥產量減產達20%[1]。化學防治及其他防治措施雖已取得階段性的成效,但實踐證明,選育和推廣抗病品種是解決白粉病危害最經濟、安全、有效的措施。本文綜述了小麥白粉病的危害、抗性基因發掘和利用等方面的研究進展,展望了小麥抗病育種的方向,旨在為小麥抗白粉病研究提供參考。
1 ?小麥白粉病發生概況
20世紀60年代,小麥白粉病(真菌氣傳性病害)僅發生在西南地區。隨著小麥矮稈品種的推廣和栽培技術的提高,使株間環境更有益于白粉病孢子的傳播,危害日漸加重[2,3]。
1.1 ?小麥白粉病菌的形態特征
小麥白粉病菌為專化性活體寄生菌。電鏡掃描觀察小麥白粉病菌的微觀形態,發現其菌絲體主要附著于葉片正面,依靠吸器汲取營養供其生長[4]。與菌絲體垂直的分生孢子梗端,串生單胞無色的分生孢子。白粉病菌的繁殖世代分為兩類:即無性與有性。無性世代中其形態為串珠狀粉孢菌,依靠分生孢子的增長[5];有性世代中,病菌形態呈球狀、無孔的閉囊殼[6,7]。
1.2 ?小麥白粉病的發生規律
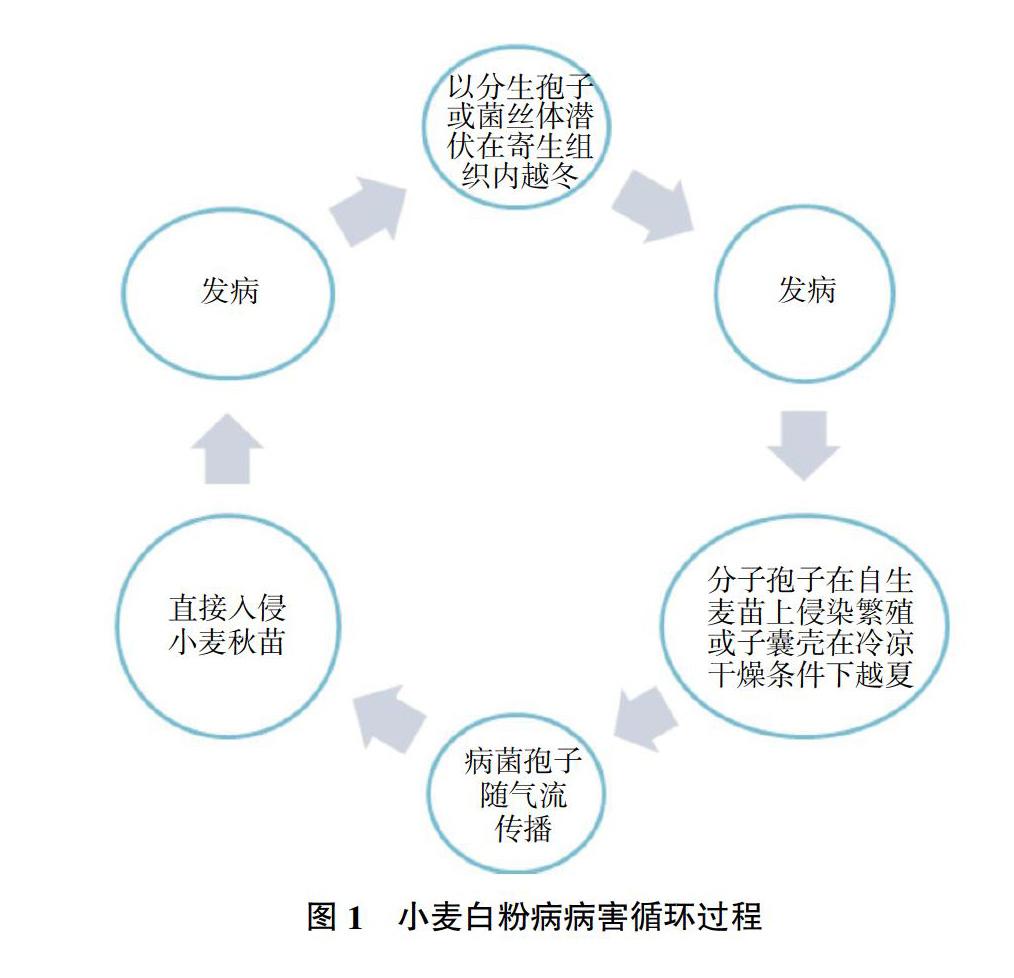
小麥白粉病菌的子囊孢子或分生孢子可借助氣流傳播。因分生孢子壽命短暫,不耐高溫,只能附著于閉囊殼或春生麥苗上順利越夏[8-10]。小麥白粉病菌越冬形式主要有以下兩種:一是分生孢子(無性孢子)的形態;二是潛伏在宿主體內的菌絲體,并且兩種越冬形式均能成功越冬。小麥白粉病病害循環過程如圖1所示[9]。
2 ?小麥白粉病菌的毒性研究
在20世紀30年代,國外學者Stakman創建病原菌生理小種的鑒別標準(通過寄主的表現型鑒定)。隨后,諸多學者根據鑒定標準對白粉病菌做了大量研究。如1956年Flor提出“基因對基因”假說,即寄主與病菌存在相互對應的關系。Wolfe等[11]根據“基因對基因”假說,提出毒性基因分析法的理念。Persaud等[12]、Parks等[13]研究發現,美國的小麥白粉病菌對Pm3a、Pm3c、Pm5a等基因的毒性頻率高。
由于病原菌生理小種變異速度快、幅度大,難以掌控毒性基因的動態變化,因此國內學者著手研究白粉病菌的毒性頻率[14]。通過采集不同白粉病菌的生理小種,鑒定發現毒性基因V1和V5的毒性頻率達90%以上;V2、V4、V2+6毒性基因頻率低[15]。毒性頻率因地域不同,存在差異[16]。在東北春麥區中,其毒性基因(V3c、V3f、V5、V8等)頻率達50%以上,V2、V4a、V5+6等毒性頻率低,可作為有效的抗性來源[17,18]; 湖北省麥區的小麥白粉病菌的毒性基因(V2、V4b、V13等)頻率低于25%;甘肅省中西部春小麥白粉病菌毒性頻率分布在16.7%[19];新疆小麥白粉病菌群體中,抗性基因Pm16和Pm21的毒性頻率為0,可作為優秀的抗源[20]。小麥白粉病菌群體時刻處于變化的狀態,因此掌握遺傳群體結構和毒性頻率的變化,對合理布局抗源具有重要意義。
3 ?小麥白粉病的抗性研究
3.1 ?小麥白粉病的生理抗性
植物具有主動防御機制,響應病原菌侵染時,體內的生長調節劑、類黃酮等生物合成途徑發生改變,激活其防御機制及增加防御反應的代謝物[21]。幾丁質酶參與植物體內重要免疫反應,能抑制病原菌的吸器數量及菌絲體的生長,對增加病程相關蛋白有促進作用,降低寄主應答幾丁質誘導的免疫反應[22,23];李新燕等[24]通過Western雜交體系檢測小麥近等基因系(含Pm21),結果發現供試材料中均含有幾丁質酶,表明幾丁質酶與白粉病菌侵染密切相關。
根據“基因對基因”假說,植物的抗病性由抗病基因與病原菌的無毒基因共同作用決定。這種主動抗病機制分為3個階段:①病原菌與寄主的初識別;②無毒基因與抗性基因特異性識別;③寄主植物產生系統獲得抗性或誘導系統抗性(ISR)[25]。
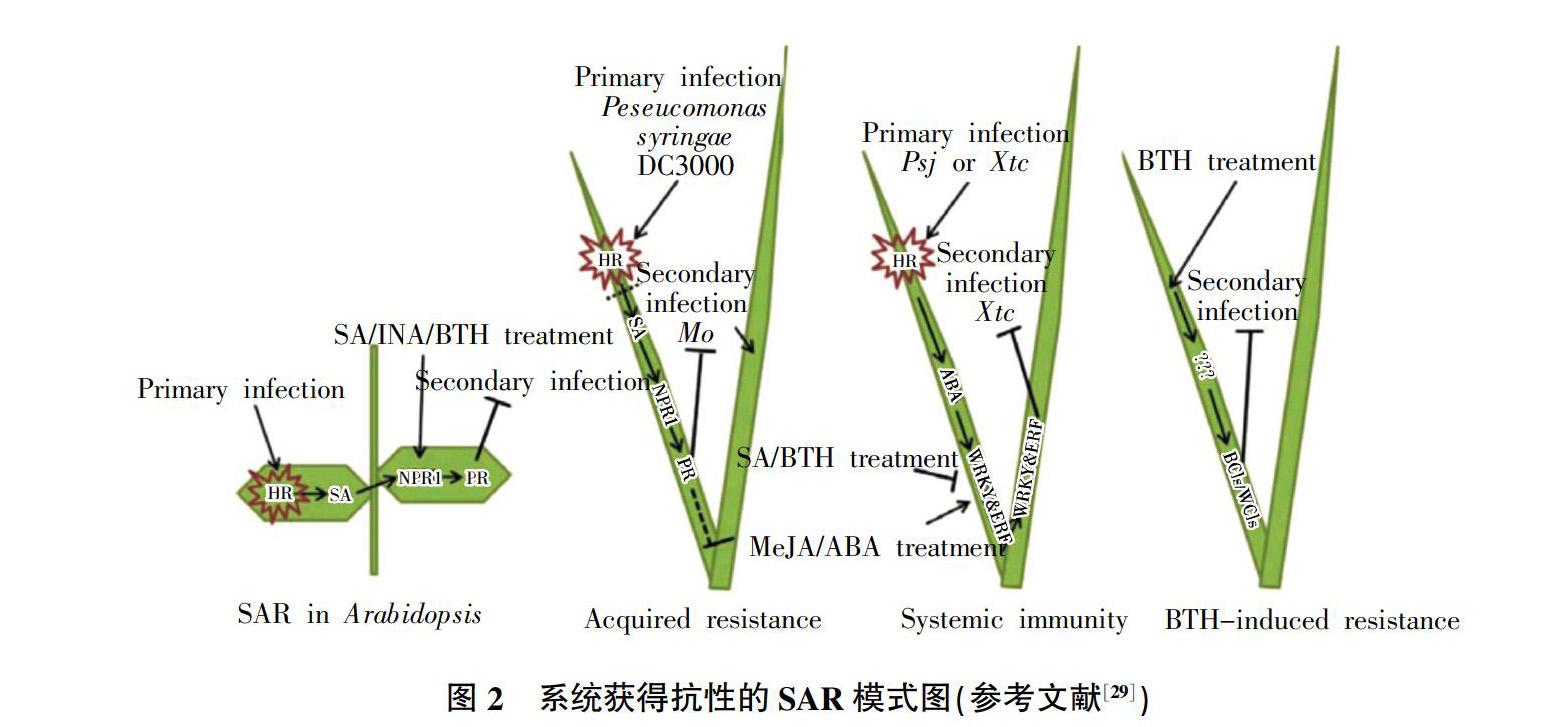
麥類作物響應白粉病菌侵染時,其免疫體系產生系統獲得抗性(SAR)。SAR過程的關鍵調控因子NPR1蛋白,是調節植物整體抗病性的重要作用因子,參與植物多種抗性代謝通路,是多個抗病信號傳導通路的交叉點。若作物(小麥)缺失NPR1的功能,將導致相關蛋白PR基因表達不完全和響應病害侵染時,系統獲得抗性(SAR)的作用全部喪失[26]。在麥類作物中,可觀察到3種類似SAR(圖2)現象,即獲得抗性(AR)、系統免疫(SI)及BTH誘導抗性(BIR)。3種現象雖與植物SAR相似,卻存在明顯的差異,推測是由不同基因通路調控。小麥應對白粉病菌侵染時,不僅產生系統獲得抗性,同時激活免疫系統中的核苷酸結合位點類受體(NLR),病原物的效應子被LRR區域識別結合,觸發植物的防御機制[27]。病原菌侵染或水楊酸類似物BTH處理均能誘導植株產生對白粉病、葉銹病、赤霉病等多種植物病害的廣譜抗性[28,29]。
在小麥與白粉病菌的相互作用中,抗性基因(如Pm3)所編碼的核苷酸與富含亮氨酸的重復型受體相結合,能賦予植株對白粉病的抗性,并且抗性基因的每個位點都能賦予對不同株系分離單株的抗性[30]。通過植物體內泛素化機制,調節HvARM1基因的表達活性,提高對白粉病的抗性[31]。相關學者通過分離編碼WDR蛋白的TaHOS15,瞬時誘導基因沉默敲低TaHOS15表達,減弱小麥對白粉病菌的易感性;TaHOS15可能還作為銜接蛋白寡聚TaHDA16到小麥防御相關基因中,抑制組蛋白乙酰化,增強防御相關轉錄及對白粉病的抗性[32]。不少學者通過病毒誘導基因沉默,沉默主要負責CHH甲基化的重組酶DRM2,結果發現DRM2下降增強了植物對白粉病菌的抗性,揭示了DNA甲基化參與調控活體寄生類真菌抗病應答的新機制[33]。
3.2 ?小麥抗性基因來源及其染色體定位
小麥抗白粉病基因分布在小麥基因組中。除3D和4D染色體外,其余染色體均存在抗性基因位點[2]。小麥抗白粉病基因主要來源于普通小麥族及其近緣種屬,分為3類,即小麥近緣種、普通小麥和小麥近緣屬,詳見表1[34]。
小麥抗白粉病基因雜亂分布在染色體上。由表2可知,第2、7染色體組的抗性基因位點最多;第1、3、4染色體組的抗性位點少。從抗性基因分布情況看,A、B染色體組的抗性基因位點較多。3D和4D染色體上尚未發現抗性基因位點。其中,Pm10、Pm11等抗性基因只對冰草白粉病菌產生抗性。來自黑麥的Pm17和Pm8抗白粉病基因均被Heun等[35]和Hsam[36]等定位于1RS。由于抗性基因來源不同、表達程度不同,因此對小麥白粉病菌的抗性存在差異。
3.3 ?小麥白粉病抗性基因研究
3.3.1 ?質量抗性基因 ?質量抗性基因抗性水平好、遺傳力較好、選育方便,是當前生產上選育的主要形式。根據“基因對基因假說”,質量抗性由小種專化性基因控制,將其命名為“Pm(Powdery mildew)”。迄今為止,共發現110個主效抗白粉病基因,其中62個基因被命名(Pm1~Pm62)、7個基因含有多個抗性位點及8個新被挖掘的抗性基因(Pm55、Pm56、Pm57、Pm58、Pm59、Pm60、Pm61、Pm62),即Pm1(Pm1a~Pm1e)、Pm2(Pm2a~Pm2c)、Pm3(Pm3a~Pm3j)、Pm4(Pm4a~Pm4d)、Pm5(Pm5a~Pm5e)、Pm8(Pm8~Pm17)和Pm24(Pm24a~Pm24b),分別位于7A、5D、1A、4A、7B、1R和1D染色體上;新抗性基因Pm59[37]、Pm58[38]、Pm57[39]、Pm56[40]、Pm55[41]、Pm62[42]、Pm61[43]、Pm60[44]被分別成功定位于7AL、2DS、2S、6RS、T5VS·5AL、2VL、4AL染色體上。根據緊密連鎖分子標記的物理定位信息,許多研究學者對抗病基因或其等位基因進行物理定位分析,確定62個抗性基因在小麥染色體上的物理位置。
3.3.2 ?數量抗性基因 ?數量抗性又稱水平抗性、成株抗性[4,7,45]。小麥白粉病的數量抗性沒有質量抗性明顯,主要表現為阻礙病菌在植株上的侵染、生長及繁殖。成株期抗性由微效多基因控制,以加性效應為主[46,47],為非專化性。成株抗性具有廣譜抗性基礎,抗性持久。相較質量抗性,數量抗性易受環境影響。隨著QTL分子標記技術的發展,相關研究者已發現140個抗白粉病QTL,分布于小麥21條染色體,其中4個QTL已被證實并在部分地區或單位廣泛應用,為兼抗小麥種質資源的選育提供理論支撐[34]。
3.3.3 ?數量抗性基因的作用機理 ?有關學者認為數量抗性即成株抗性,可能是系統獲得抗性或者誘導系統抗性,亦或是兩種抗性機制與周圍環境共同作用決定。其中“SAR”誘導的抗性反應與植物防御體系有關;ISR與土壤微生物、植物次生代謝有關,并伴隨抗病相關基因(如NPR1)的表達[48]。研究表明,在煙草[49]、大豆[50]等作物體內存在類似成株抗性的體制。在小麥體內尚未發現,有待進一步探索發掘。
4 ?小麥抗白粉病分子標記應用
隨著DNA分子標記技術的不斷成熟與發展,被廣泛應用于小麥傳統育種。研究者利用相關分子標記檢測99份育種材料,結果發現,只有4份材料攜帶抗病基因,其余材料未發現任何抗病基因[51,52];隋建樞等[53]運用特異性標記,對F2群體194個單株進行檢測,發現標記SCAR1265與抗白粉病基因Pm21呈共分離狀態;曹廷杰等[54]利用相關分子標記檢測908份供試材料,發現只有少量材料攜帶廣譜抗源基因Pm21;劉易科等[55]利用相關分子標記檢測湖北省小麥主要品種,發現所用標記均可在參試材料中檢測出相應基因。相關研究者先后利用特異性分子標記,篩選出若干抗源材料,為小麥抗病育種奠定了基礎[56-59]。
國內不少學者對分子標記篩選進行了大量的研究。如利用SSR標記對小麥F2代群體進分析,篩選到與抗病基因緊密連鎖的分子標記[60-63];利用分子標記對小麥F2代群體進行PCR分析,獲得與抗白粉病基因Pm6、Pm4a、Pm16緊密連鎖的特異性標記[64-67];殷貴鴻[68]、陳松柏等[69]采用不同的抗病基因標記對后代群體進行連鎖遺傳分析,估算抗病基因與分子標記的相對遺傳距離。因此,篩選與抗病基因連鎖的分子標記的相關研究奠定了分子標記輔助選擇的基礎。
5 ?小麥抗白粉基因的克隆
隨著現代分子生物技術(如克隆)的廣泛應用,小麥白粉病抗性基因被相繼克隆,如Pm21[70]、Pm38/Lr34/Yr18/Sr57/Ltn1、Lr67/Yr4/Pm46/Sr55、Pm60[44]。特別是兼抗葉銹病、白粉病、稈銹病的成株抗性基因的克隆,為小麥兼抗種質資源的篩選奠定基礎。
6 ?展望
6.1 ?小麥白粉病菌生理小種的動態監測
由于中國獨特的氣候與環境,導致白粉病菌生理小種基因突變或遺傳重組,與寄主協同進化,使抗性基因失去作用[71]。若產生新生理小種或白粉病菌群結構改變,將給農業生產帶來毀滅性的后果。因此,必須監測小麥白粉病菌生理小種毒性基因頻率,掌握該菌群體遺傳結構的動態變化,才能正確使用抗病材料,避免盲目利用抗源。
6.2 ?小麥白粉病抗源多樣化
小麥白粉病菌生理小種時常變化易使生產上推廣的抗病品種失去抗性,對小麥產量造成威脅。因此,國內外學者針對小麥的垂直抗性,產生了幾點設想:①抗源的合理分布;②選育、推廣多系品種及利用混合品種;③國內外收集、積累抗源。抗病育種實踐證明,由多個抗病基因控制的復雜抗性比單個基因的抗性更穩定、持久。國內學者針對中國小麥白粉病菌新生理小種產生等問題,提出應加強收集、利用“二線抗源”[72]的方案,注重多基因水平抗性的研究和選育,借以解決持久抗性的問題。
6.3 ?抗性基因差異
不同類型的抗病基因存在抗性質量或抗譜能力差異,導致其利用時間、地域也不盡相同。據普查,貴州大學農學院麥作研究中心于2007年選育的貴農19號小麥品種(黔審麥2007002)在田間對白粉病表現出高抗甚至免疫,作為穩定持久的抗源,被很多育種單位利用。若抗性基因的抗譜基礎狹窄,則難以應對復雜多變的生理小種。特此建議盡量從小麥近緣屬中尋求具有廣譜抗性基礎的抗性基因,作為小麥抗白粉病育種的有效抗源。
6.4 ?基因聚合育種
病原菌的高度變異性和抗性資源的不恰當使用,致使小麥抗病育種面臨抗源匱乏、抗譜遺傳基礎狹窄的威脅。育種家們應遵循“滾動回交育種”[73]的策略,廣泛收集不同類型的高質量抗性基因,運用MAS的技術優勢,使不同抗性基因通過基因聚合手段實現有效結合,不僅可彌補抗性基因間的差異,還能提高抗病基因強度,延長抗性基因使用壽命,使抗病表現更加持久,保證小麥抵御病害的能力。因此,應以基因聚合育種為主,常規育種為輔將作為今后抗病育種工作的重要方向。基因聚合育種雖已取得階段性的成效,但大部分仍停留在理論研究中,這需要育種工作者們在今后的工作中不斷努力探索,將理論成效轉化為實踐。
參考文獻:
[1] 王宏梅.貴農6號小麥抗白粉病基因的分子標記[D].貴陽:貴州大學,2009.
[2] 申曉柯.小麥白粉病新抗源的遺傳分析及抗性基因的分子標記定位[D].成都:四川農業大學,2013.
[3] 劉萬才,劉振東,黃 ?沖,等.近10年農作物主要病蟲害發生危害情況的統計和分析[J].植物保護,2016,42(5):1-9.
[4] 王保勤.蘭考906抗白粉病新基因的分子標記篩選[D].鄭州:河南農業大學,2008.
[5] 巢凱翔.三個小麥品種(系)抗條銹病和白粉病基因的遺傳分析和分子作圖[D].陜西楊凌:西北農林科技大學,2018.
[6] 郝元峰.小麥抗白粉病基因的分子標記定位及標記輔助選擇[D].濟南:山東農業大學,2008.
[7] 馬宏棋.普通小麥抗白粉病新基因的發掘和分子標記定位[D].南京:南京農業大學,2010.
[8] 黃苗苗.甘肅省小麥白粉病菌的溫度敏感性及其抗病性遺傳分析[D].蘭州:甘肅農業大學,2015.
[9] 才旦卓瑪.2012年小麥白粉病菌溫度敏感性和遺傳多樣性的相關性研究[D].西寧:青海大學,2014.
[10] 楊美娟,黃坤艷,韓慶典.小麥白粉病及其抗性研究進展[J].分子植物育種,2016,14(5):1244-1254.
[11] WOLFE M S,SCHWARZBACH E.Patterns of race changes in powdery mildews[J].Annu Rev Phytopathol,1978,16:159-180.
[12] PERSAUD R R.Virulence genes and virulence gene frequencecies of Blumeria graminis f.sp.tritici in Ohio[J].Plant disease,1995,79(5):494-499.
[13] PARKS R,CARBONE I,MURPHY J P,et al.Virulence structure of the eastern of U.S. wheat powdery mildew population[J].Plant disease,2008,92(7):1074-1082.
[14] 段雙科,許育彬,吳興元.小麥白粉病菌致病毒性和抗病基因及抗病育種研究進展[J].麥類作物學報,2002,22(2):83-86.
[15] 郭建國.甘肅中部小麥白粉病流行影響因素初步探討及我國部分麥區小麥白粉菌抗藥性監測[D].蘭州:甘肅農業大學,2005.
[16] 肖仲久.貴州省小麥白粉菌群體多樣性分析及品種抗性研究[D].貴陽:貴州大學,2006.
[17] 魏松紅,曹遠銀,牟連曉.東北春麥區小麥白粉病菌生理小種鑒定及毒性基因分析[J].植物保護學報,2006,33(1):27-31.
[18] 季宏平,孟慶林,王 ?芊,等.黑龍江省小麥白粉病菌毒性結構和毒力頻率研究[J].黑龍江農業科學,2007(3):49-52.
[19] 王 ?龍,王生榮,甘麗萍.甘肅中西部春小麥白粉菌群體毒性分析[J].西北農業學報,2005,14(1):106-110.
[20] 王振花,劉 偉,高海峰,等.新疆小麥白粉病菌群體的毒性監測和分析[J].新疆農業科學,2017,54(10):1903-1910.
[21] YUAN H J,ZENG X Q,YANG Q F,et al.Gene coexpression network analysis combined with metabonomics reveals the resistance responses to powdery mildew in Tibetan hulless barely[J].Scientific reports,2018,8(1):14928.
[22] TOYDA H,MATSUDA Y,RAMAGE T,et al.Suppression of powdery mildew pathogen by chitinase microinjection into barely coleoptileepidermal cells[J].Plant cell reports,1991(10):217-220.
[23] SCHMMBAUM A,MAUCH F,VOGELI U,et al.Plant chitinases are potential inhibititors of fungal growth[J].Nature,1986(24):365-367.
[24] 李新燕,陳文品,馬正強.小麥白粉菌誘導的幾丁質酶同工酶分析[J].南京農業大學學報,2002,25(1):1-4.
[25] 李愛麗.小麥抗白粉病基因同源序列的分離鑒定及分子標記的研究[D].河北保定:河北農業大學,2002.
[26] 韓永光,馬利剛,趙 樂,等.植物抗性基因NPR1研究進展[J].安徽農業科學,2018,46(26):18-20.
[27] ZHANG X,DODDS P N,BERNOUX M.What do we know about NOD-like receptors in plant immunity?[J].Annual of review of phytopathology,2017,55:205-229.
[28] GAO J,BI W S,LI H P,et al.WRKY transcription factors associated with NPR1-mediated acquired resistance in barley are potential resources to improve wheat resistance to Puccinia triticina[J].Frontiers in plant science,2018,9:1486.
[29] WANG X D,BI W S,GAO J,et al.Systemic acquired resistance,NPR1,and pathogenesis-related genes in wheat and barley[J].Journal of integrative agriculture,2018,17(11):2468-2477.
[30] KOLLER TERESA,BRUNNER SUSANNE,HEREN GERHARD,et al.Field grown transgenic Pm3e wheat line show powdery mildew resistance and no fitness costs associated with high transgene expression[J].Transgenic research,2019,28(1):9-20.
[31] RAJARAMAN J,DOUCHKOV D,LUECK S,et al.The partial duplication of an E3-ligase gene in Triticeae species mediates resistance to powdery mildew fungi[EB/OL].BioRxiv,2017,https://doi.org/10.1101/190728.
[32] LIU J,ZHI P F,WANG X Y,et al.Wheat WD40-repeat protein TaHOS15 functions in a histone deacetylase complex to fine-tune defense response to Blumeria graminis f.sp.tritici[J].Journal of experimental botany,2019,70(1):255-268.
[33] GENG S,KONG X,SONG G,et al.DNA methylation dynamics during the interaction of wheat progenitor Aegilops tauschii with the obligate biotrophic fungus Blumeria graminis f. sp. tritici[J].New phytologist,2019,221(2):1023-1035.
[34] GUO J,LIU C,ZHAI S N,et al.Molecular and physical mapping of powdery mildew resistance genes and QTLs in wheat:A review[J].Agricultural science & technology,2017, 18(6):965-970.
[35] HEUN M,FRIEBE B,BUSHUK W.Chronosomal location of the powdery mildew resistance gene of Amigo wheat[J].Phytopathology,1990,80(10):1129-1133.
[36] HSAM S L K,ZELLER F J. Evidence of allelism between genes Pm8 and Pm17 and chromosomal location of powdery mildew and leaf rust resistance genes in the common wheat cultivar ‘Amigo[J].plant breeding,1997,116(2):119-122.
[37] TAN C,LI G,COWGER C,et al.Characterization of Pm59,a novel powdery mildew resistance gene in Afghanistan wheat landrace PI 181356[J].Theor Appl Genet,2018,131(5):1145-1152.
[38] WIERSMA A T,PULMAN J A,BROWN L K,et al.Identification of Pm58 from Aegilops tauschii[J].Theor Appl Genet,2017,130(6):1123-1133.
[39] LIU W,KOO D H,XIA Q,et al.Homoelologous recombination-based transfer and molecular cytogenetic mapping of powdery mildew-resistant gene Pm57 from Aegilops searsii into wheat[J].Theor Appl Genet,2017,130(4):841-848.
[40] HAO M,LIU M,LIU J T,et al. Introgression of powdery mildew resistance gene Pm56 on rye chromosome Arm 6RS into wheat[J].Frontiers in plant science,2018,9:1040.
[41] ZHANG R Q,SUN B X,CHEN P D,et al.Pm55,a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat[J].Theoretical and applied genetics,2016,129(10):1975-1984.
[42] ZHANG R Q,FAN Y L,KONG L N,et al. Pm62,an adult-plant powdery mildew resistance gene introgressed from Dasypyrum villosum chromosome arm 2VL into wheat[J].Theoretical and applied genetics,2018,131(12):2613-2620.
[43] SUN H G,HU J H,SONG W,et al. Pm61:A recessive gene for resistance to powdery mildew in wheat landrace Xuxusanyuehuang identifed by comparative genomics analysis[J].Theoretical and applied genetics,2018,131:2085-2097.
[44] ZOU S H,WANG H,LI Y W,et al. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat[J].New phytologist,2018,218(1):298-309.
[45] 張志良.小麥地方品種紅蚰麥抗白粉病基因的發掘和定位[D].南京:南京農業大學,2015.
[46] DAS M K,GRIFFEY C A.Diallel analysis of adult-plant resistance to powdery mildew in wheat[J].Crop Sci,1994,34(4):948-952.
[47] DAS M K,GRIFFEY C A.Gene action for adult-plant resistance to powdery mildew in wheat[J].Genome,1995,38(2):277-282.
[48] PIETERSE C M,WEES S C,PELT J A,et al.A novel signaling pathway controlling induced ?systemic resistance in Arabidopsis[J].Plant cell,1998,10(9):1571-1580.
[49] WYATT S,PAN S,KUC J.β-1,3-Glucanase,chitinase and peroxidase activities in tobacco tissues resistant and susceptible to blue mould as related to flowering,age and ucker development[J].Physiology molecular plant pathology,1991,39(6):433-440.
[50] WARD E W B,STOSSEL P,LAZAROVITS G. Similarities between age-related and race-specific resistance of soybean hypocotyls to Phytophthora megasperma var. sojae[J].Phytopathology,1981,71:504-508.
[51] 張小輝,暢志堅,喬麟軼,等.99份小麥地方品種抗白粉病種質發掘及其分子鑒定[J].山西農業科學,2017,45(5):692-698.
[52] 張小輝.小麥抗白粉病種質資源發掘及其分子標記鑒定[D].太原:山西大學,2017.
[53] 隋建樞,任明見,徐如宏.貴農775抗白粉病基因的分子標記定位[J].貴州農業科學,2013,41(1):4-7.
[54] 曹廷杰,陳永興,李 ?丹,等.河南小麥新育成品種(系)白粉病抗性鑒定與分子標記檢測[J].作物學報,2015,41(8):1172-1182.
[55] 劉易科,朱展望,佟漢文,等.湖北省主要小麥品種抗病基因分析[J].分子植物育種,2018,16(4):1040-1049.
[56] 張增艷,陳 ?孝,張 ?超,等.分子標記選擇小麥抗白粉病基因Pm4b、Pm13和Pm21聚合體[J].中國農業科學,2002,35(7):789-793.
[57] 董建力,張增艷,王敬東,等.3種小麥抗白粉病基因聚合體的STS和SCAR標記[J].西北農業學報,2007,16(3):64-67.
[58] 高安禮,何華綱,陳全戰,等.分子標記輔助選擇小麥抗白粉病基Pm2、Pm4a和Pm21的聚合體[J].作物學報,2005,31(11):16-21.
[59] 董 ?娜,張亞娟,張軍剛,等.分子標記輔助小麥抗白粉病基因Pm21和Pm13聚合育種[J].麥類作物學報,2014,34(12):1639-1644.
[60] 朱玉麗,王黎明,王洪剛.小麥抗白粉病基因Pm5e的SSR標記研究[J].分子植物育種,2008,6(6):1080-1084.
[61] 朱玉麗.小麥抗白粉病基因Pm2和Pm5e的SSR分子標記研究[D].山東泰安:山東農業大學,2007.
[62] 王黎明,朱玉麗,李興鋒,等.小麥抗白粉病基因Pm2的SSR標記篩選[J].植物保護學報,2011,38(3):216-220.
[63] 張軍剛,董 ?娜,閆文利,等.小麥抗白粉病基因Pm13的SSR標記篩選[J].河南農業科學,2014,43(10):62-66.
[64] 王俊美,劉紅彥,王 ?飛,等.小麥抗白粉病基因Pm6的微衛星標記鑒定[J].植物病理學報,2007,37(3):329-332.
[65] 王俊美.小麥抗白粉病基因Pm4、Pm6的PCR鑒定[D].陜西楊凌:西北農林科技大學,2005.
[66] 王 ?瑞,劉紅彥,王俊美,等.小麥抗白粉病基因Pm6的PCR標記鑒定[J].麥類作物學報,2007,27(3):421-424.
[67] 羅瑛皓.小麥抗白粉病基因Pm16的SSR標記定位以及多基因累加體的分子標記輔助選擇[D].成都:四川農業大學,2003.
[68] 殷貴鴻.小麥抗條銹病和白粉病基因的分子標記[D].陜西楊凌:西北農林科技大學,2009.
[69] 陳松柏,蔡一林,周榮華,等.小麥抗白粉病基因Pm4的STS標記[J].西南農業大學學報,2002,24(3):231-234.
[70] H G HE,S Y ZHU,Y Y JI,et al.Map-based cloning of the gene Pm21 that confers broad spectrum resistance to wheat powdery mildew[J].BioRxiv,2017,177857.
[71] 蘭彩霞.普通小麥條銹病和白粉病成株抗性QTL定位[D].北京:中國農業科學院,2010.
[72] 蔡士賓,程順和,吳紀中,等.引進小麥白粉病二線抗源的鑒定與改良利用[J].麥類作物學報,2005,25(6):124-128.
[73] 張菲菲. 轉抗赤霉病基因小麥的聚合育種和回交轉育研究[D].武漢:華中農業大學,2015.
