SIRT1對糖尿病胃輕癱小鼠胃排空的影響及機制
2020-07-21 12:21:54周冬梅
首都醫科大學學報 2020年4期
鄭 晗 周冬梅,2 苗 蓓,3 李 偉,2*
(1.徐州醫科大學第一臨床學院,江蘇徐州 221004;2.徐州醫科大學附屬醫院內分泌科,江蘇徐州 221002;3.徐州醫科大學附屬醫院消化內科,江蘇徐州 221002)
糖尿病胃輕癱(diabetic gastroparesis,DGP)是以腹脹、早飽、厭食、噯氣、惡心、體質量下降等為典型表現的糖尿病慢性合并癥之一。DGP嚴重降低糖尿病患者的生活質量,多達50%的DGP患者出現明顯的焦慮和/或抑郁(伴腹痛者尤甚)[1]。另外,DGP會加重糖尿病患者血糖控制難度,增加患者的住院率,加重患者的經濟負擔[2]。因此,探究DGP的發病機制并找到行之有效的防治辦法對糖尿病患者而言尤為重要。
氧化應激被認為是糖尿病各種慢性合并癥的統一發生機制[3],在DGP發病過程中發揮著重要作用[4]。Cajal間質細胞(interstitial cells of Cajal,ICC)是胃慢波活動的起搏細胞,可以生成和傳播調節胃腸平滑肌收縮的電信號。研究[4]顯示,DGP的發生與氧化應激所致ICC網絡受損有關。
沉默信息調節因子1(silent information regulator 1,SIRT1)是一種細胞煙酰胺腺嘌呤二核苷酸依賴性脫乙酰基酶,主要通過使許多類型的關鍵轉錄因子和輔因子脫乙酰化,如:P53、核因子- κB(nuclear factor κB,NF-κB)、O型叉頭轉錄因子盒蛋白(forkhead box protein,FOXO)和過氧化物酶體增生物激活受體共激活因子1(peroxisome proliferator activated receptor gamma coactivator,PGC-1),參與細胞周期停滯和衰老,這些因素可能會影響與諸如糖尿病等代謝性疾病有關的細胞途徑[5],SIRT1與糖尿病及相關性疾病的發生、發展有關。SIRT1可以通過改善線粒體的氧化代謝,并在氧化應激下正向調節線粒體功能,以抵御氧化損傷[6]。因此,上調SIRT1可能改善糖尿病小鼠的胃排空延遲。
血紅蛋白氧合酶-1(heme oxygenase-1,HO-1)被認為是可以對抗氧化應激的重要的細胞防御機制[7],HO-1的轉錄是由多種氧化還原依賴性信號通路如促分裂原活化蛋白激酶(mitogen activated protein kinase,MAPK),信號傳導及轉錄活化因子3(signal transducer and activator of transcription 3,STAT3),特別是轉錄因子核因子E2相關性因子2(nuclear factor erythroid 2 related factor 2,Nrf2)誘導的[8]。BTB-CNC異體同源體1(BTB and CNC homology 1,Bach1)作為一種重要的轉錄抑制因子,可以通過競爭性抑制Nrf2與抗氧化反應元件(antioxidant response element,ARE)的偶聯,進而下調抗氧化因子HO-1的基因轉錄[9]。SIRT1也可以調控HO-1基因轉錄[10]。因此,上調SIRT1可能通過下調組織中Bach1蛋白濃度,進一步調控HO-1的基因轉錄,從而發揮抗氧化作用。這可能成為DGP防治的新的靶點。
白藜蘆醇(resveratrol,Res)是一種非黃酮類多酚化合物,具有顯著的抗氧化活性。Res也是SIRT1的激活劑,可調節細胞周期,線粒體生物合成以及脂質和碳水化合物代謝,其抗氧化、抗炎、抗癌和心血管保護作用已在實驗[11]中得到證實,但是Res作為SIRT1激活劑是否可以改善DGP小鼠的胃排空延遲,尚未見文獻報道。
本實驗主要探究SIRT1對DGP小鼠胃排空影響以及可能的作用機制,為臨床預防和治療DGP患者提供新的思路。
1 材料與方法
1.1 實驗動物
6周雄性C57BL/6J小鼠,體質量20~23 g,購于濟南朋悅實驗動物繁育有限公司,實驗動物許可證號:SCXK(魯)2019-0003。室溫(23±1) ℃,12 h光照周期,自由飲水攝食。該研究得到徐州醫科大學動物實驗倫理委員會的批準,倫理委員會審批號:2016036。
1.2 藥物與試劑
Res(美國MCE公司),鏈脲佐菌素(streptozotocin,STZ)(美國Sigma公司);小鼠抗兔SIRT1多克隆抗體(美國Proteintech公司),小鼠抗兔Bach1多克隆抗體(美國ABclonal公司),小鼠抗兔c-kit多克隆抗體(美國Affinity Biosciences公司),小鼠抗兔HO-1多克隆抗體(美國Affinity Biosciences公司);丙二醛(malondialdehyde,MDA)試劑盒(江蘇凱基生物技術公司 ),超氧化物歧化酶(superoxide dismutase,SOD)試劑盒(江蘇凱基生物技術公司 )。
1.3 分組、造模
實驗前適應性飼養1周。6周齡C57BL/6J小鼠采用隨機數字表法分為空白組、模型組、溶劑對照組和Res組。除空白組小鼠以外,其余小鼠按160 mg/kg一次性腹腔注射STZ[以1%(質量分數)溶于枸櫞酸鈉緩沖液中,現配現用],保證充足的飼料和飲用水。空白組小鼠腹腔注射等量的枸櫞酸鈉緩沖液。造模72 h 后采集尾靜脈血檢測隨機血糖,隨機血糖≥ 16.7 mmol/L為模型制備成功。
1.4 藥物干預
模型建立成功后進行藥物干預,Res組小鼠按照30 mg·kg-1·d-1的劑量給予SIRT1激活劑Res溶解于DMSO中,注射時0.9%(質量分數)氯化鈉注射液稀釋腹腔注射,溶劑對照組給予等量的DMSO 0.9%(質量分數)氯化鈉注射液溶液腹腔注射,干預6周。
1.5 固體胃排空法檢測胃排空
干預6周后,所有小鼠禁食、不禁水20~24 h,給予小鼠已知量的食物,自由進食3 h。3 h結束時,剩余食物稱質量,以確定食物攝入量。小鼠再次禁食禁水4 h。4 h后,頸椎脫臼法處死小鼠,取出胃組織,測量整個胃的質量,沿胃大彎剪開胃組織,0.9%(質量分數)氯化鈉注射液清洗胃內容物,濾紙吸干水分后測量胃凈重。胃排空率按以下公式計算:胃排空率(4 h內)=(1-胃內殘留食物質量/食物攝入量)×100%。
1.6 取材
將取出的胃組織沿胃小彎剪開,取胃竇部位,一部分用4%(質量分數)中性甲醛固定24 h后,石蠟包埋,進行免疫組織化學染色,另一部分,液氮速凍,于-80 ℃ 冰箱保存,用于Western blotting實驗。
1.7 免疫組織化學染色
將石蠟包埋的胃竇組織切成6 μm切片,常規脫蠟再水化,按照二步法檢測試劑盒步驟進行染色,一抗為兔抗SIRT1多克隆抗體(1∶200)或兔抗Bach1多克隆抗體(1∶100),用PBS代替一抗做陰性對照。DAB顯色后,封片,在顯微鏡下進行觀察。攝片后,用Image J軟件分析結果,測得積分吸光度值(integrated optical density,IOD)代表陽性表達的強度,IOD=吸光度(A)×面積。
1.8 Western blotting檢測
將新鮮冷凍胃竇組織剪碎后加入裂解液RIPA與酶抑制劑PMSF,進行勻漿(冰上進行),約5 min。將勻漿液在4 ℃下,12 000 r/min,離心15 min,收集上清液,BCA法測蛋白濃度。等量蛋白樣品經十二烷基硫酸鈉-聚丙烯酰氨凝膠電泳(SDS-PAGE)后,將分離出的蛋白轉移的PVDF膜上,用3%(質量分數)BSA封閉2 h,加入一抗4 ℃ 孵育過夜。所用一抗如下:SIRT1抗體(1∶1 000)、Bach1抗體(1∶1 000)、HO-1抗體(1∶1 000)、c-kit抗體(1∶1 000)、β-actin抗體(1∶10 000)。次日TBST清洗后加入一抗種屬特異性二抗(1∶1 000)室溫孵育1 h,再次洗膜后用ECL顯像試劑盒顯影,置于凝膠成像儀中照相,并用Image J軟件分析條帶。
1.9 比色法檢測組織總超氧化物歧化酶(total superoxide dismutase,T-SOD)活力以及MDA含量
將新鮮組織勻漿(冰上進行)后,4 ℃下4 000 r/min離心15 min,取上清液分別按照SOD試劑盒和MDA試劑盒步驟測定胃組織中的T-SOD活力以及MDA含量。
1.10 統計學方法
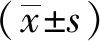
2 結果
2.1 各組小鼠的一般情況
空白組小鼠精神狀態良好,反應靈敏,毛色亮澤,進食飲水正常,對抓取抵抗明顯;模型組和溶劑對照組小鼠體型消瘦,食量減少,腹部脹滿,毛色干枯無光伴有脫毛,活動遲緩,與空白組相比,體質量明顯下降,血糖明顯升高(P<0.05);Res組小鼠飲水及食量均增加,活動性增強,一般情況較模型組和溶劑對照組有改善,體質量及血糖與模型組和溶劑對照組小鼠相比,差異無統計學意義(P>0.05,表1、2)。
2.2 各組小鼠的胃排空率
與空白組相比,模型組和溶劑對照組小鼠的胃排空率明顯下降(P<0.05);與模型組和溶劑對照組小鼠相比,Res組小鼠胃排空率明顯升高(P<0.05);溶劑對照組與模型組小鼠間的差異無統計學意義,詳見表3。
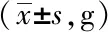
表1 各組小鼠的體質量變化
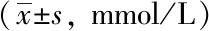
表2 各組小鼠隨機血糖濃度變化
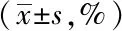
表3 各組小鼠的胃排空率
2.3 免疫組織化學檢測各組小鼠胃竇組織中SIRT1、Bach1蛋白濃度
以細胞中出現黃色或棕黃色顆粒為陽性表達。免疫組織化學結果顯示,SIRT1和Bach1主要在胞核內表達(圖1)。用Image J軟件分析免疫組織化學結果測得IOD值,與空白組小鼠相比,模型組和溶劑對照組小鼠胃組織中的SIRT1蛋白濃度明顯下降,Bach1蛋白濃度明顯升高(P<0.05);與模型組和溶劑對照組小鼠相比,Res組小鼠胃組織中的SIRT1蛋白濃度明顯升高,Bach1蛋白濃度明顯下降(P<0.05);模型組與溶劑對照組小鼠間比較,差異無統計學意義(表4)。
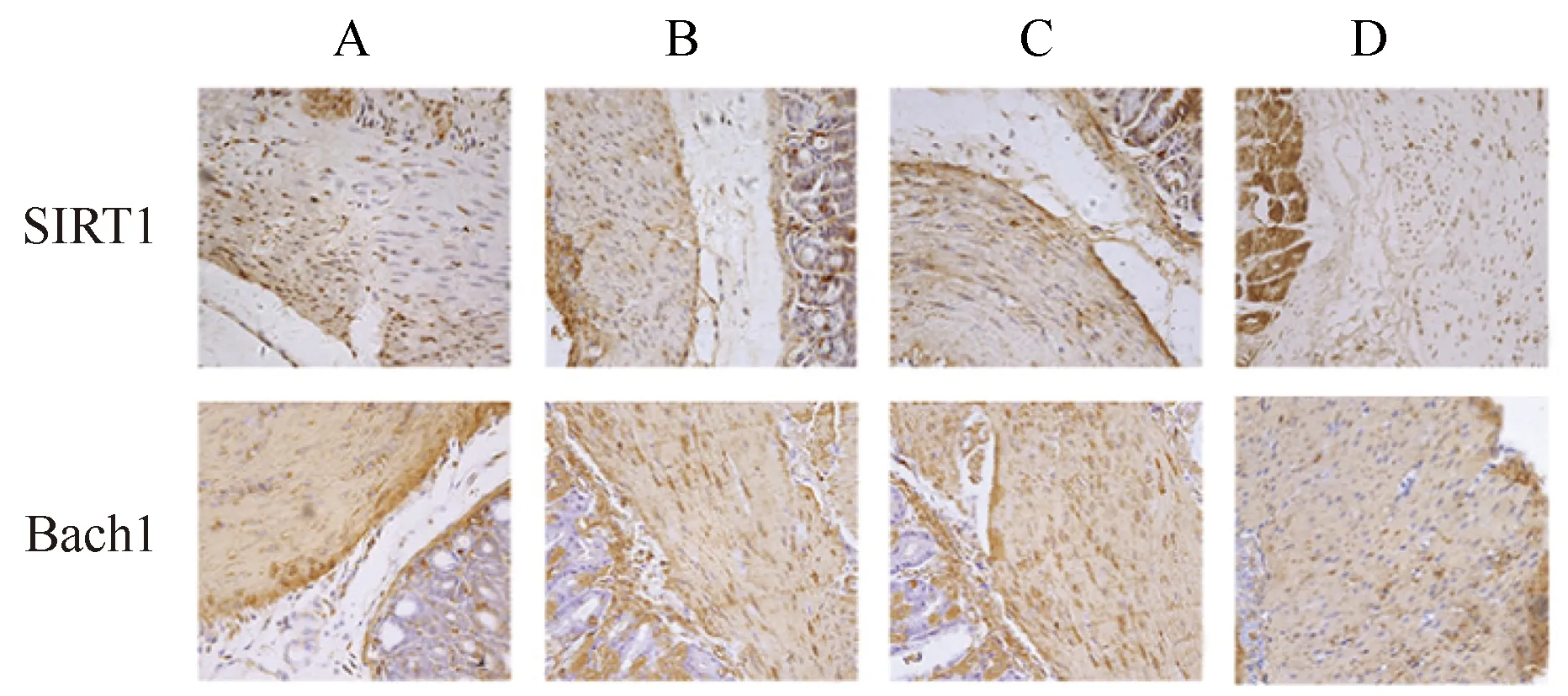
圖1 各組小鼠胃竇組織SIRT1、Bach1表達Fig.1 Expression of SIRT1 and Bach1 in gastricantrum tissue of mice in each group(SP,40×)A: Blank group; B: Model group; C: Solvent control group; D: Res group; SIRT1: silent information regulator 1; IOD: integrated optical density; Res: resveratrol.
表4 SIRT1和Bach1在各組小鼠胃竇組織中的IOD值
2.4 Western blotting法檢測各組小鼠胃竇組織中SIRT1、Bach1、c-kit以及HO-1蛋白濃度
Western blotting法檢測結果顯示,與空白組相比,模型組小鼠和溶劑對照組小鼠胃組織中的SIRT1、c-kit以及HO-1蛋白濃度明顯下降,Bach1蛋白濃度明顯升高(P<0.05),與模型組和溶劑對照組相比,Res組小鼠胃組織中SIRT1、c-kit以及HO-1蛋白濃度明顯升高,Bach1蛋白濃度明顯下降(P<0.05);模型組和溶劑對照組小鼠之間的差異無統計學意義(圖2)。
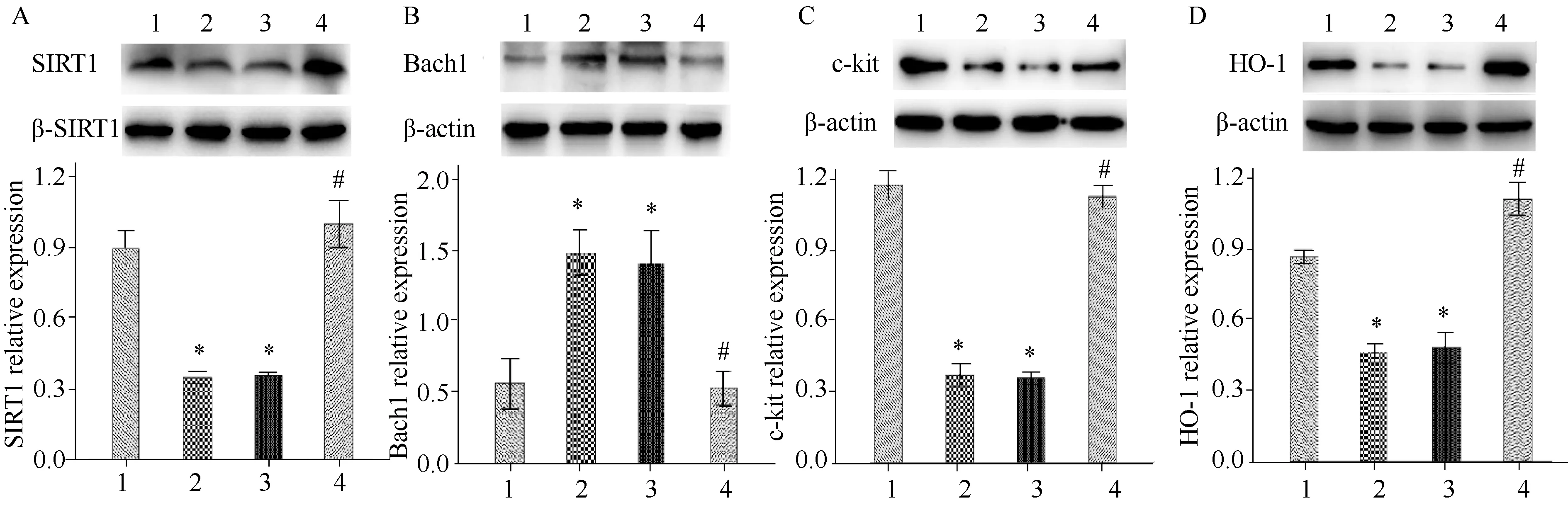
圖2 各組小鼠胃竇組織SIRT1、Bach1、c-kit和HO-1濃度Fig.2 SIRT1, Bach1, c-kit and HO-1 protein levels in gastric antrum tissue of mice in each groupA: SIRT1 protein level; B: Bach1 protein level; C: c-kit protein level; D: HO-1 protein level; 1: Blank group; 2: Model group; 3: Solvent control group; 4: Res group; *P<0.05 vs Blank group, #P<0.05 vs Model group; Res: resveratrol; SIRT1: silent information regulator 1; HO-1: heme oxygenase-1; Res: resveratrol.
2.5 各組小鼠胃竇組織氧化應激相關因子MDA含量和T-SOD活力
與空白組小鼠相比,模型組和溶劑對照組小鼠胃組織中MDA含量明顯升高,T-SOD活力明顯下降(P<0.05);與模型組和溶劑對照組小鼠比較,Res組小鼠胃組織中的MDA含量下降,T-SOD活力升高(P<0.05);模型組小鼠與溶劑對照小鼠組間的差異無統計學意義(P>0.05,表5)。
表5 各組小鼠胃竇組織MDA含量和T-SOD活力
3 討論
氧化應激是DGP發病的重要機制之一[4, 12]。持續的高血糖狀態導致細胞線粒體內產生過多的陰離子自由基,當超過細胞內抗氧化防御系統的清除能力時,就會在細胞線粒體內積累,積累的自由基可以作用于脂質發生過氧化反應,從而引起組織損傷。MDA是脂質過氧化反應的終產物之一,它會引起蛋白質、核酸等生物大分子的交聯聚合,產生細胞毒性。SOD是機體酶抗氧化系統的一員,能清除超氧陰離子自由基,保護細胞免受氧化損傷。持續的糖尿病高血糖狀態會導致體內自由基和脂質過氧化醛的產生增加,糖尿病相關氧化應激介導的器官功能損害是導致多種慢性合并癥的關鍵因素[3],血清MDA濃度的升高與胃排空延遲密切相關[13]。ICC是胃慢波運動的起搏細胞,對胃腸動力的維持至關重要[14]。研究[4, 15]顯示,DGP的發病與氧化應激所致的ICC減少有關。HO-1是機體重要的抵抗氧化應激的防御因子之一,研究[4]顯示,在胃排空延遲的非肥胖型糖尿病(non-obese diabetic mice,NOD)小鼠的胃組織中發現HO-1上調的消失以及ICC標志物酪氨酸激酶受體(tyrosine kinase receptor,c-kit)表達的減少,但是在胃排空正常的NOD小鼠中未發現這種現象,施用HO-1誘導劑可以保護ICC網絡免受氧化應激損傷,并使延遲的胃排空正常化[4]。本實驗結果證實了模型組小鼠胃排空延遲,且該組小鼠胃組織中HO-1上調的缺失同時伴隨著c-kit表達的減少,此外,本研究中胃排空延遲的小鼠胃組織中的MDA濃度明顯升高,且T-SOD活力顯著下降,氧化及抗氧化失衡,氧化應激濃度升高。以上證據表明糖尿病小鼠胃組織中較高的氧化應激濃度是ICC網絡損傷的重要原因之一,最終會導致糖尿病小鼠胃排空率的下降。
研究[16]顯示,SIRT1廣泛參與基因調節,基因組穩定性維持,細胞凋亡、自噬、衰老、增生和腫瘤發生,主要通過使組蛋白和非組蛋白靶標脫乙酰基,在組織穩態和許多疾病的表觀遺傳調控中也具有關鍵作用。對糖尿病動物模型的研究[17-19]顯示,低濃度的SIRT1會加重氧化應激損傷,而激活SIRT1可以減輕這種損傷:一項針對小鼠糖尿病心肌病的研究[20]顯示,SIRT1下調會使心肌細胞中PGC-1α和HO-1的濃度降低,心肌活性氧產生增加,導致糖尿病性心肌病;另外,上調SIRT1可以通過激活Nrf2/HO-1通路減輕氧化應激,來改善糖尿病腎病的缺血再灌注損傷[21]。由此可見,HO-1是SIRT1發揮抗氧化作用重要的下游因子之一。本研究中STZ誘導的DGP小鼠胃組織中SIRT1表達減少,進一步導致HO-1上調的缺失,因此,SIRT1蛋白濃度的下調可能參與了DGP的發生與發展,但引起SIRT1下調的具體機制有待進一步研究。相較模型組,給予SIRT1激活劑的糖尿病小鼠的胃排空率以及胃組織中的c-kit表達顯著升高,SIRT1及其下游因子HO-1的蛋白濃度也明顯升高,表明上調SIRT1可以保護糖尿病小鼠胃組織中的ICC網絡,并維持小鼠正常的胃排空,這與其上調HO-1有關;另外,與模型組小鼠相比,給予SIRT1激活劑的糖尿病小鼠胃組織中MDA含量明顯下降,T-SOD活力顯著升高,提示上調SIRT1還可以通過抑制組織中的過氧化反應,并提高T-SOD活力來發揮抗氧化能力。
Bach1主要通過與小Maf癌蛋白形成異二聚體,并與小Maf識別元件(antioxidant response element,ARE)結合以抑制靶基因(包括編碼HO-1的基因)表達[22]。在缺乏Bach1的小鼠中,HO-1在許多組織中以較高濃度組成性表達,這表明Bach1在HO-1表達的負調控中起主要作用[23]。在高氧誘導小鼠胚胎成纖維細胞衰老模型中,Bach1蛋白表達明顯升高,一定程度上阻止了抗氧化轉錄因子Nrf2的核積累,導致細胞氧化應激濃度升高,從而加速細胞衰老[24]。另外一項針對肺纖維化小鼠的研究[25]發現,Bach1敲除后,抗氧化因子(如HO-1和谷胱甘肽過氧化物酶1)表達增加,肺泡和間質細胞的炎性浸潤以及肺結構的破壞顯著減弱。Bach1缺乏對氧化應激誘導的細胞損傷具有預防作用,缺乏Bach1的小鼠可以使四氧嘧啶誘導的胰島β細胞的凋亡減少,而這種保護作用的機制歸因于HO-1的上調[22]。以上證據表明,Bach1可能是組織細胞出現氧化應激損傷的關鍵環節,其可以成為預防糖尿病及其相關性疾病的可能的治療靶標。本實驗結果顯示,胃排空延遲的糖尿病小鼠胃組織中Bach1濃度升高,提示Bach1可能是DGP發病的關鍵因子。筆者假設DGP小鼠胃組織中SIRT1蛋白濃度下降可能是組織中Bach1高濃度的重要原因之一,本實驗中給予SIRT1激活劑的小鼠胃組織中Bach1濃度較DGP小鼠相比明顯下降,HO-1表達顯著升高,因此,可以認為SIRT1維持糖尿病小鼠正常胃排空的機制可能與其下調Bach1蛋白濃度,進一步促進抗氧化因子HO-1基因轉錄有關。
此外,本研究給予Res干預可以改善DGP小鼠的胃排空延遲,但是并未使糖尿病小鼠的血糖濃度下降,這是否與實驗動物種類以及激活劑劑量和療程有關尚未可知。在本實驗條件下,上調SIRT1維持糖尿病小鼠正常的胃排空的作用不是通過影響血糖途徑實現的。
綜上所述,上調SIRT1可以通過調控Bach1和HO-1濃度,保護DGP小鼠胃組織中的ICC網絡免受氧化損傷,并改善DGP小鼠胃排空延遲。此項研究為DGP患者的防治提供了新的理論依據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2022年5期)2022-08-24 02:35:42
中老年保健(2022年1期)2022-08-17 06:14:56
中老年保健(2021年5期)2021-08-24 07:07:20
中老年保健(2021年11期)2021-08-22 03:15:16
學苑創造·A版(2020年9期)2020-10-13 09:41:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
西南軍醫(2016年6期)2016-01-23 02:21:19
西南軍醫(2015年2期)2015-01-22 09:09:37
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
