伊犁馬對補喂α-酮異己酸的吸收及其某些相關代謝的研究
2021-01-25 06:45:50賴曉鶴張文杰艾畢肯楊開倫
中國飼料 2021年1期
賴曉鶴,張文杰,韓 明,艾畢肯,劉 律,楊開倫
(新疆農業大學動物科學學院,新疆肉乳用草食動物營養重點實驗室,新疆烏魯木齊830052)
α-酮異己酸(KIC)是亮氨酸合成與分解的中間代謝產物,能夠與亮氨酸通過支鏈氨基酸轉氨酶(BCAT)進行可逆的轉化,KIC也是肝臟生成β-羥基-β-甲基丁酸(HMB)的直接前體物質。研究表明,KIC具有促進肌肉蛋白質合成(Van和Edwarda,2005)、抑制蛋白質降解(Tischler 等,1982)、調節能量代謝(Moghei等,2016;Zhou 等,2010)及氮平衡(Sapir等,1983)等多種生理作用。在機體內KIC能夠與亮氨酸相互轉化,但KIC在降低血漿游離氨基酸濃度、促進骨骼肌蛋白質合成和降低血漿中尿素氮濃度等方面更加有效(Duan等,2019)。
Duan等(2019)給仔豬通過飼糧補喂1.25%KIC-Ca發現,其可促進肌肉蛋白質合成能力和抑制蛋白質降解。給禁食仔雞補喂2.25 mg/kg的KIC能夠起到抑制肌原纖維蛋白降解的作用(Nakashima等,2007)。研究發現,通過飼糧補喂1.25%的KIC-Ca能夠促進仔豬肌肉組織中脂肪酸氧化,減少脂肪沉積(Duan等,2016)。桑丹等(2009)每天給綿羊分別補喂 0.5、1.0、1.5 g 的瘤胃保護性KIC-Ca發現,其能夠調節氮平衡。每天補喂15 g的瘤胃保護性KIC-Ca能夠增加羔羊體重(Kuhlman等,1988)。 目前,KIC作為營養添加劑的研究較少,將KIC作為一種添加劑補喂伊犁馬,伊犁馬對KIC的吸收及其對體內其他氨基酸會產生怎樣的影響鮮見報道。本試驗選用1歲伊犁馬為研究對象,探究伊犁馬對飼糧中補喂不同水平α-酮異己酸的吸收及其某些相關代謝情況,為KIC在馬匹飼糧中的應用提供參考依據。
1 材料與方法
1.1 試驗設計及飼糧組成 采取單因素多水平試驗設計方法,挑選20匹年齡1歲,體重(295.25±25.15)kg的伊犁馬,分成4組,每組 5匹,分別為對照組、試驗Ⅰ組、試驗Ⅱ組、試驗Ⅲ組。試驗飼糧由精料補充料和干草組成。不同組馬匹所喂精料補充料不同。精料補充料配方及營養水平見表1,委托新疆伊犁州昭蘇縣駿力農牧科技有限公司生產加工。在試驗飼糧的基礎上,各試驗組補喂不同水平的KIC,KIC以鈣鹽的形式進行補喂(購自張家港市思普生化有限公司,純度為99%,有效濃度為87%)。玉米和次粉先按照1:1進行混勻,混勻后與KIC鈣鹽按照1:1制作成顆粒料,將顆粒料分為兩份與精料補充料混合補喂。試驗Ⅰ、Ⅱ、Ⅲ組每匹馬KIC有效補喂量分別為6、12、18 g/d。本試驗中KIC的補喂量參考了Yarrow等(2007)在運動員上的應用,選用低劑量0.02 g/kg的研究結果,進行為期25 d的補喂試驗。精料補充料和干草中氨基酸含量見表2。
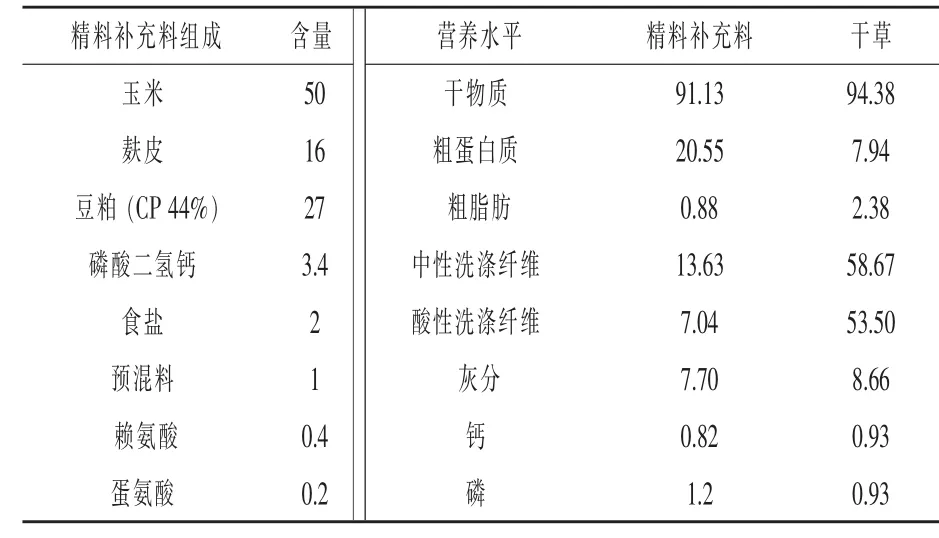
表1 精料補充料配方及營養水平(干物質基礎)%
1.2 試驗動物飼養管理 試驗期間所有馬匹均在同一馬廄,單欄飼養。干草每天每匹馬(8±0.8)kg, 分別在 08:30、13:30、18:30 和 23:30 分 4 次飼喂。精料補充料每天每匹馬3 kg,分別于每天09:00和21:00分2次飼喂。先粗后精,先采食一定量干草,再補給精料補充料,待精料補充料采食完畢后,將馬匹趕入活動圈,繼續采食一定量的干草,自由飲水。每天按時打掃馬廄,清除糞便,更換墊草。
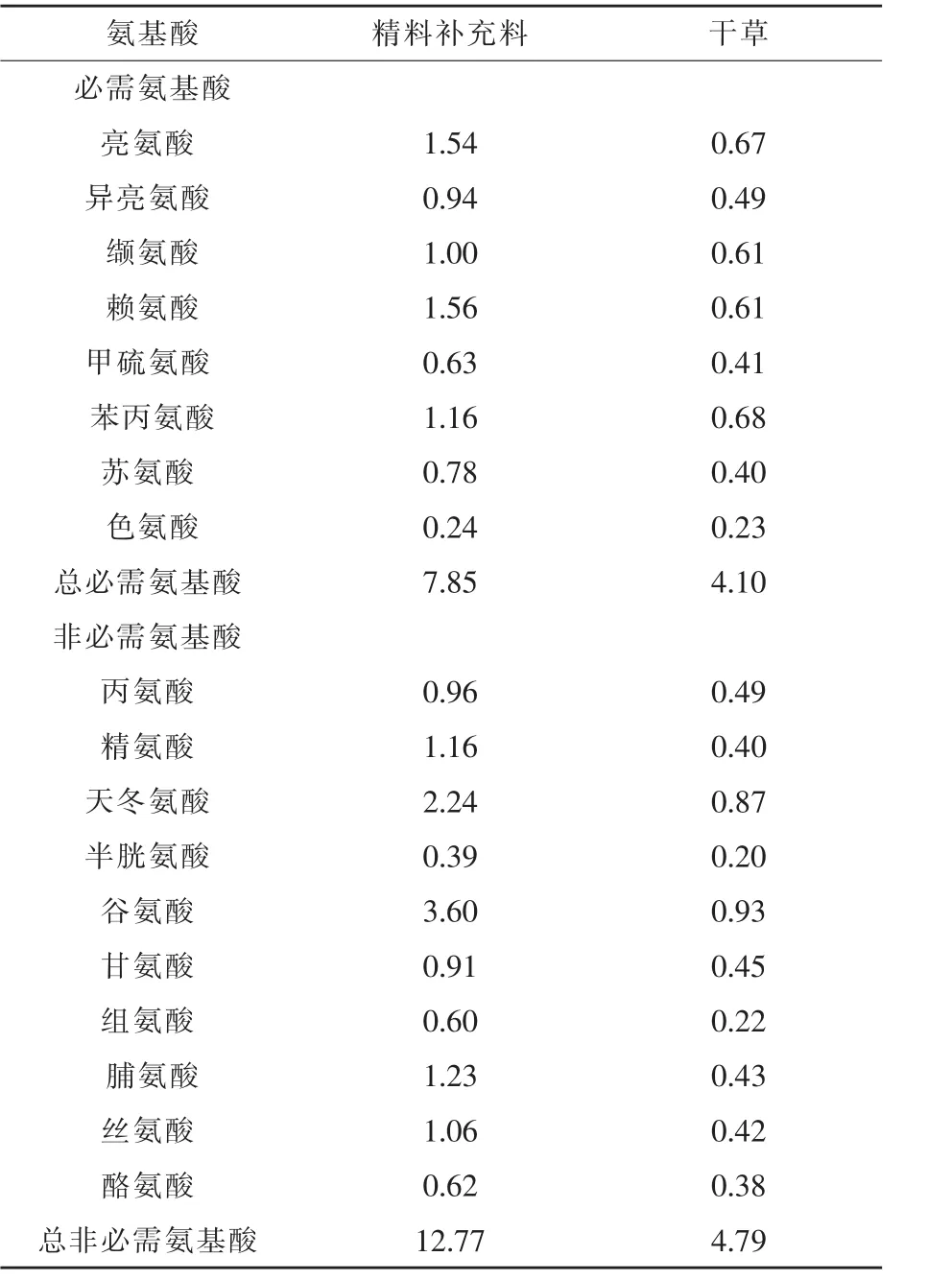
表2 精料補充料和干草中氨基酸含量(干物質基礎)%
1.3 樣品采集及指標的測定
1.3.1 血漿α-酮異己酸和β-羥基-β-甲基丁酸測定 在試驗第25天采集血液,血液采集時間以補喂前 0 h 開始(8:30)以及補喂后的 1 h(10:00)、2 h(11:00)和 4 h(13:00),通過頸靜脈采集血液置于肝素鈉抗凝管中,以3500 r/min離心10 min,收集上清血漿,分裝于1.5 mL Eppendorf管中,分裝完畢后在-20℃冰箱中進行凍存。
血漿α-酮異己酸和β-羥基-β-甲基丁酸濃度均由北京普瑞華盛生物科技有限公司采用高效液相色譜串聯質譜聯用儀進行定量分析檢測。
1.3.2 血漿游離氨基酸測定 在試驗第25天采集血液,通過頸靜脈于補喂后2 h(11:00)采集血液至肝素鈉抗凝管中,以3500 r/min離心10 min,收集上清血漿,分裝于1.5 mL Eppendorf管中,分裝完畢后在-20℃冰箱中進行凍存。
血漿5-羥色胺濃度由北京華英生物技術研究所采用酶聯免疫吸附法測定;血漿游離氨基酸濃度由北京普瑞華盛生物科技有限公司采用高效液相色譜串聯質譜聯用儀進行定量分析檢測。
1.4 數據處理與分析 數據采用Excel 2010進行初步整理,試驗結果以“平均值和標準誤”表示。試驗數據采用SPSS 19.0統計軟件中的單因素方差分析(one-way ANOVA)進行差異顯著性檢驗,差異顯著用Duncan氏法進行多重比較。采用正交多項式對比,檢驗日糧中α-酮異己酸濃度增加的線性和二次效應。以P<0.05為差異顯著的判斷標準,P<0.01為差異極顯著水平,0.05<P<0.10為有差異趨勢。
2 結果與分析
2.1 伊犁馬對α-酮異己酸的吸收 日糧α-酮異己酸補喂水平與伊犁馬血漿中α-酮異己酸濃度的關系見圖 1,當補喂 0 g/horse·meal(指每匹馬每頓補喂的克數,下同)的KIC(對照組)時,血漿中KIC濃度最低;3 g/horse·meal(試驗Ⅰ組)時,血漿中 KIC 濃度升高。且當 6 g/horse·meal(試驗Ⅱ組)時,血漿中KIC濃度最高。隨著KIC補喂水平的增加,血漿中KIC濃度的變化呈先上升后下降的趨勢 (擬合方程為y=-0.0365x2+0.577+11.016,R2=0.9653)。結果顯示,補喂不同水平的KIC對伊犁馬血漿中KIC濃度無顯著影響。
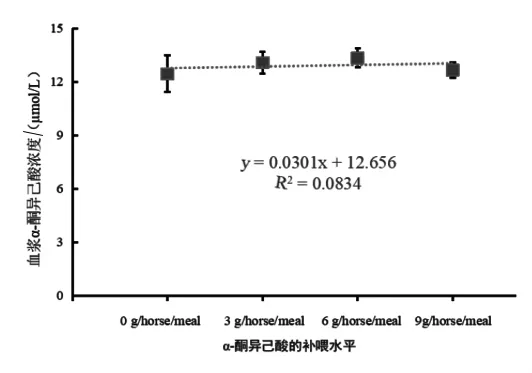
圖1 日糧α-酮異己酸補喂水平與伊犁馬血漿中α-酮異己酸濃度的關系
補喂α-酮異己酸對伊犁馬血漿α-酮異己酸濃度的影響見表3。與對照組相比,補喂前(即0 h)各組間血漿KIC濃度差異不顯著(P>0.05)。與對照組相比,補喂后1 h試驗Ⅰ、Ⅱ組和Ⅲ組血漿KIC濃度分別提高了11.2%、7.86%和21.51%(P>0.05);補喂后2 h試驗Ⅰ組、試驗Ⅱ組和試驗Ⅲ組血漿KIC濃度分別提高了8.66%、18.77%和8.84%,但均差異不顯著(P>0.05);補喂后4 h各試驗組均差異不顯著(P>0.05)。3個試驗組馬匹血漿KIC濃度都在補喂后1 h出現峰值。
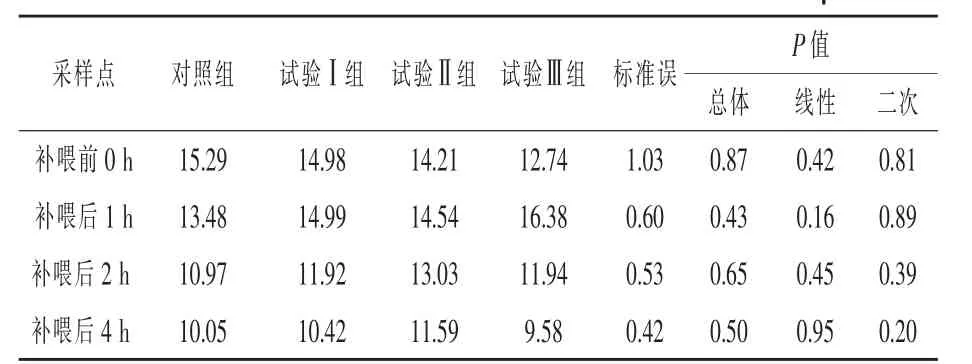
表3 補喂α-酮異己酸對伊犁馬血漿中α-酮異己酸濃度的影響 μmol/L
2.2 補喂α-酮異己酸對伊犁馬血漿β-羥基-β-甲基丁酸濃度的影響 日糧α-酮異己酸補喂水平與伊犁馬血漿中β-羥基-β-甲基丁酸濃度的關系見圖2,當KIC補喂水平為3 g/horse·meal(試驗Ⅰ組)時,血漿中HMB濃度最低;且當6 g/horse·meal(試驗Ⅱ組)時,血漿中HMB 濃度最高。隨著KIC補喂水平的增加,血漿中HMB濃度的變化呈先下降后上升再下降的波浪趨勢。結果顯示,補喂不同水平的KIC對伊犁馬血漿中HMB濃度無顯著影響。
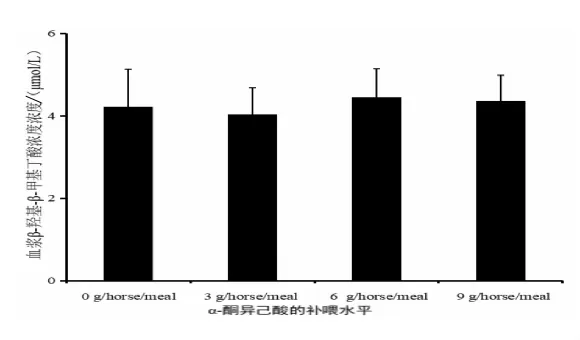
圖2 日糧α-酮異己酸補喂水平與伊犁馬血漿中β-羥基-β-甲基丁酸濃度的關系
補喂α-酮異己酸對伊犁馬血漿β-羥基-β-甲基丁酸濃度的影響見表4。與對照組相比,補喂前0 h各組間血漿HMB濃度差異不顯著 (P>0.05)。與對照組相比,補喂后1 h血漿HMB濃度各試驗組均差異不顯著(P>0.05);補喂后2 h試驗Ⅰ、Ⅱ、Ⅲ組血漿HMB濃度分別提高了5.84%、15.74%和14.97%(P>0.05);補喂后4 h各試驗組分別提高了8.95%、18.93%和14.58%,但均差異不顯著(P > 0.05)。
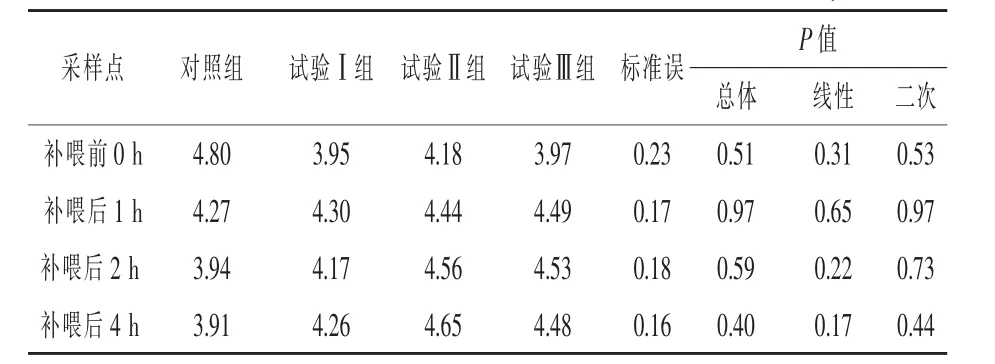
表4 補喂α-酮異己酸對伊犁馬血漿中β-羥基-β-甲基丁酸濃度的影響 μmol/L
2.3 補喂α-酮異己酸對伊犁馬血漿游離氨基酸及其代謝物濃度的影響 補喂α-酮異己酸對伊犁馬血漿游離氨基酸濃度的影響見表5。與對照組相比,各試驗組血漿KIC濃度分別提高了8.66%、18.77%和8.84%,但均差異不顯著(P>0.05);各試驗組血漿HMB濃度均差異不顯著 (P>0.05);試驗Ⅱ組和試驗Ⅲ組血漿亮氨酸濃度分別提高了22.40%和22.24%,且達到顯著水平(P<0.05);試驗Ⅰ組和試驗Ⅲ組血漿異亮氨酸濃度降低了23.08%和32.34%,且達到極顯著水平(P<0.01);試驗Ⅰ組和試驗Ⅲ組血漿纈氨酸濃度分別降低了15.75%和14.13%,且達到顯著水平(P<0.05);試驗Ⅲ組血漿賴氨酸、甲硫氨酸、精氨酸和谷氨酰胺濃度分別降低了 28.43%、22.22%、21.81%和23.60%,且均達到顯著水平(P<0.05);各試驗組血漿5-羥色胺濃度分別降低了7.5%、28.03%和10.28%,且試驗Ⅱ組比試驗Ⅰ組和試驗Ⅲ組分別降低了22.22%和24.68%,組間差異均達到極顯著水平(P<0.01);試驗Ⅲ組血漿三甲基組氨酸濃度降低了18.74%,但差異不顯著(P>0.05)。試驗Ⅲ組血漿天冬酰胺濃度比試驗Ⅱ組降低了26.50%,且組間差異達到顯著水平(P<0.05),試驗Ⅱ組血漿支鏈氨基酸濃度比試驗Ⅰ組降低了18.80%,且達到顯著水平(P<0.05)。
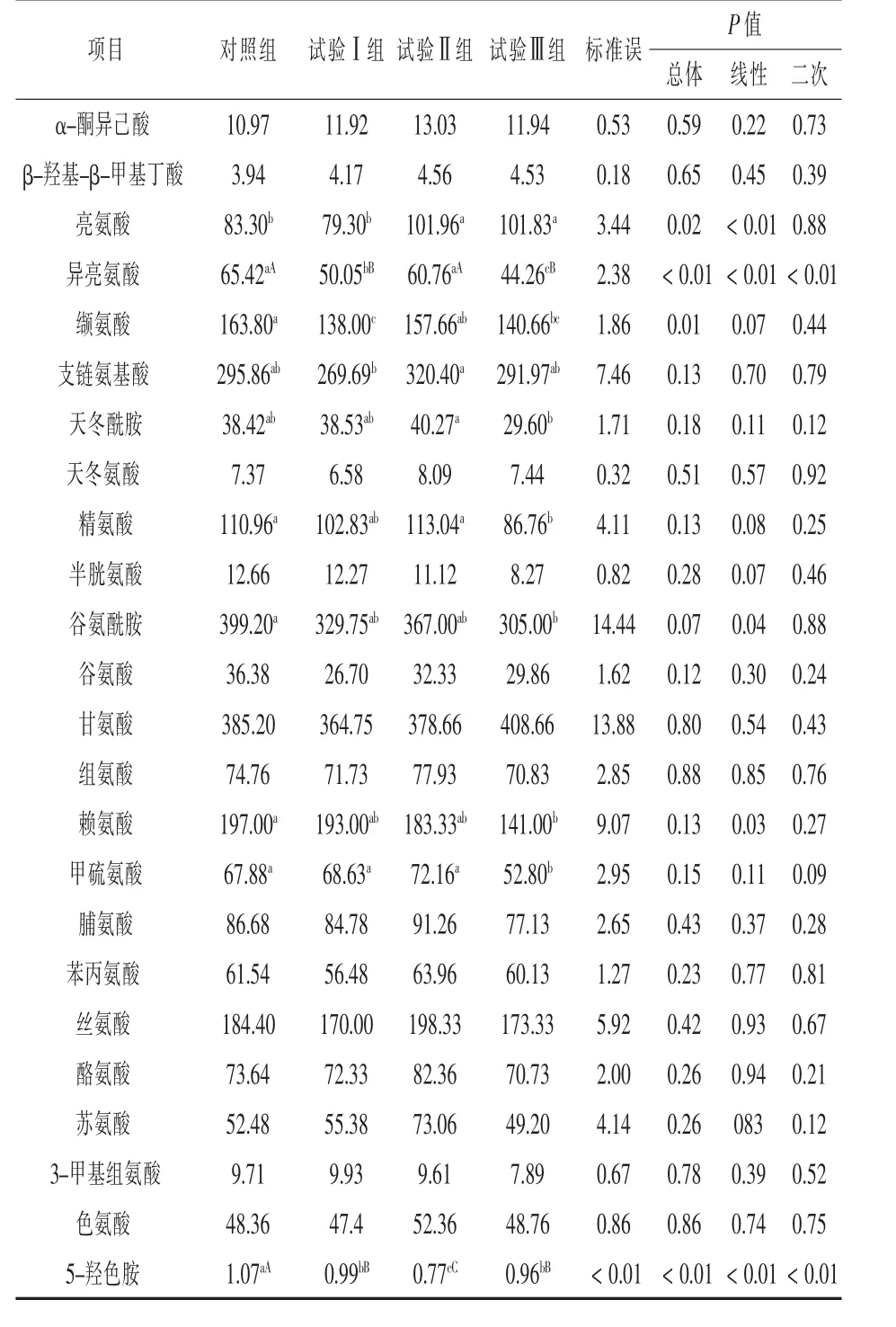
表5 補喂α-酮異己酸對伊犁馬血漿氨基酸濃度的影響 μmol/L
3 討論
3.1 伊犁馬對α-酮異己酸的吸收 在動物機體內KIC有3種來源,從含有KIC的食物中直接攝取;也可以直接攝取KIC復合鹽或通過攝取亮氨酸代謝產生KIC,進而提高血漿中KIC濃度。因此,本試驗探究了日糧中補喂不同水平KIC與伊犁馬血漿中KIC濃度的關系。本試驗中,隨著KIC補喂水平的增加,血漿中KIC濃度的變化呈先上升后下降的趨勢。且當KIC補喂水平為6 g/horse·meal(試驗Ⅱ組)時,血漿中KIC濃度最高。結果表明,補喂不同水平的KIC對伊犁馬血漿中KIC濃度的影響不顯著。KIC作為中間代謝物,在血漿中濃度處于相對穩定的狀態。
測定動物飼喂前后血漿中間代謝物濃度的變化是動物營養消化代謝研究中常用的重要指標。從肝臟釋放到血液中的KIC用于合成骨骼肌中的蛋白質或形成丙氨酸和谷氨酰胺。肝外組織中,KIC與亮氨酸可以相互轉化,然后KIC可以重新合成亮氨酸或進一步氧化。本試驗中,補喂KIC 1 h后血漿KIC濃度各試驗組均達到了峰值。說明通過補喂KIC提高了伊犁馬血漿中KIC濃度。本試驗中,各試驗組在補喂前及補喂后1、2、4 h血漿KIC濃度均呈先上升后下降趨勢,KIC作為亮氨酸的中間代謝產物,在機體中的存留時間較短,通常會在肝臟組織中代謝或參與亮氨酸的相互轉化。KIC的半衰期極短,血液中的KIC的代謝進行的很快(Giron等,2016),可能是因為KIC的氧化速率很快,這就解釋了各組血漿中KIC的濃度在飼喂后1 h達到峰值,然后下降的原因。Duan等(2019)給仔豬補喂飼糧1.25%的KIC-Ca,1 h后血漿KIC濃度升高。Escobar等(2010) 發現給仔豬靜脈滴注 400 μmol/kg·h 的KIC,1 h后血漿KIC濃度升高,與本試驗結果一致。本試驗中,對照組血漿中KIC濃度的變化呈下降趨勢,各試驗組血漿KIC濃度的變化表現為“單峰”模式(在補喂后1 h血漿KIC濃度達到峰值),且試驗Ⅲ組血漿KIC濃度的變化表現為“單峰”模式最明顯。
3.2 補喂α-酮異己酸對伊犁馬血漿中β-羥基-β-甲基丁酸濃度的影響 機體內源性產生的 β-羥基-β-甲基丁酸(HMB)的量很少(Zhong等,2019)。KIC在肝細胞溶質中發生不可逆的氧化脫羧反應,大約5%的KIC通過KIC二氧化酶氧化生成HMB,生成的HMB大約有20%隨尿液排出體外 (Molfino等,2013)。本試驗中,隨著KIC補喂水平的增加,血漿中HMB濃度的變化呈先下降后上升再下降的趨勢,呈現出波浪狀。值得注意的是,伊犁馬中補喂低水平KIC(3 g/horse·meal)與未添加KIC相比,血漿中KIC濃度下降,可能是受到性別、個體差異和環境等外界因素的影響。當KIC補喂水平為6 g/horse·meal(試驗Ⅱ組)時,血漿中HMB濃度最高。結果表明,補喂不同水平的KIC伊犁馬血漿中HMB濃度的變化不顯著。HMB作為KIC的代謝產物,通過內源性添加KIC,不會影響血漿中HMB濃度。
無論是內源性還是額外攝入的HMB,大約有30%的HMB被肌肉吸收 (Freund等,1978)。這就表明在高濃度時,HMB代謝的主要場所是肌肉組織(Van 和 Nissen,1992)。 研究表明,HMB不僅能夠提高肌肉功能,還能提高脂肪代謝速率并降低蛋白質分解速率,促進組織細胞修復,減輕肌肉損傷并延緩肌肉疲勞(Norton和Layman,2006)。補喂KIC提高了血漿KIC濃度,從而導致血漿HMB濃度有所升高。本試驗中,對照組血漿HMB濃度的變化呈下降趨勢,試驗I組和試驗Ⅲ組在補喂前及補喂后1、2、4 h血漿HMB濃度的變化呈先上升后下降趨勢,但試驗Ⅱ組血漿中HMB濃度呈上升趨勢。說明KIC在調節伊犁馬血漿HMB濃度中可能存在劑量效應。Duan等(2019)給仔豬補喂1.25%KIC-Ca,1 h后血漿HMB濃度升高,與本試驗結果一致。血漿KIC濃度的變化表現出的“單峰”模式,并沒有使血漿HMB濃度出現相同的趨勢,說明通過補喂KIC,機體代謝產生的HMB是十分有限的。因此補喂KIC在一定程度上增加了血漿HMB濃度,且試驗II組血漿HMB濃度較高。
3.3 補喂α-酮異己酸對伊犁馬血漿游離氨基酸及其代謝物濃度的影響
3.3.1 補喂α-酮異己酸對伊犁馬血漿亮氨酸及支鏈氨基酸濃度的影響 亮氨酸作為必需氨基酸之一,必須通過日糧攝入的方式來維持機體正常生長及生產活動正常進行(Wu,2014)。本試驗中,通過給伊犁馬飼糧中補喂KIC可顯著提高血漿中亮氨酸濃度,與對照組相比,試驗Ⅱ組和試驗Ⅲ組血漿亮氨酸濃度分別提高了22.40%和22.24%。血漿亮氨酸濃度升高,導致血漿纈氨酸和異亮氨酸濃度顯著下降。由于三種氨基酸之間互為拮抗關系,亮氨酸增加必定會影響其他一種或兩種氨基酸在機體內的吸收和利用 (Papet等,1988)。支鏈氨基酸對氨基酸與蛋白質的代謝具有調節作用,尤其是亮氨酸可以通過激活雷帕霉素靶蛋白(mTOR)信號通路調節骨骼肌的蛋白質合成(Wilkinson 等,2013;Suryawan 等,2011),而異亮氨酸和纈氨酸可以保證三種支鏈氨基酸之間的平衡(Suryawan 等,2013)。 Duan 等(2019)給仔豬補喂1.25%的KIC-Ca提高了血漿亮氨酸濃度,血漿纈氨酸、異亮氨酸濃度降低,與本試驗結果一致。本試驗中,試驗Ⅰ組與試驗Ⅱ組相比,血漿中支鏈氨基酸濃度顯著下降。由于馬匹血漿中游離氨基酸濃度會受到性別、環境和溫度等因素影響 (Hackl等,2006),因此補喂不同水平的KIC對血漿支鏈氨基酸濃度的影響有待進一步研究。
3.3.2 補喂α-酮異己酸對伊犁馬血漿3-甲基組氨酸濃度的影響 3-甲基組氨酸是組氨酸通過蛋白質合成甲基化后生成的氨基酸,主要存在于機體內骨骼肌蛋白的肌球蛋白和肌纖蛋白中。骨骼肌中蛋白質周轉主要包括:骨骼肌收縮蛋白、肌動蛋白、肌球蛋白的周轉,會導致血漿中3-甲基組氨酸釋放,3-甲基組氨酸不能被機體重新利用合成蛋白,最終被排出體外(Rathmacher和Nissen,1998)。在一定程度上3-甲基組氨酸也可作為肌原纖維蛋白水解的指標(桑丹等,2009)。本試驗中,血漿中3-甲基組氨酸濃度無顯著變化,但整體呈下降趨勢,且試驗Ⅲ組血漿3-甲基組氨酸濃度降低了18.74%。研究發現,給禁食的雛雞補喂2.25 mg/kg的KIC降低了血漿3-甲基組氨酸濃度(Nakashima 等,2007),Sapir等(1983)給術后患者每天靜脈滴注70 mmol的KIC-Na降低了血漿3-甲基組氨酸濃度,與本試驗結果相似。由于本試驗中KIC的補喂量分早晚兩次補喂,因此,伊犁馬血漿中3-甲基組氨酸濃度的變化可能與KIC的補喂量有關。本試驗中沒有收集馬尿,進而得到3-甲基組氨酸的排出量,因而沒有直接證據表明動物機體內蛋白質的降解情況。但從本試驗中的馬匹增重情況可以得出,與對照組相比,各試驗組的平均日增重有升高趨勢。因此,KIC可能有提高伊犁馬增重的趨勢。
3.3.3 補喂α-酮異己酸對伊犁馬血漿其他氨基酸及其代謝物濃度的影響 血漿游離氨基酸濃度反映了動物攝入飼糧氨基酸的水平。谷氨酰胺是馬駒血漿中濃度最多的氨基酸 (Manso等,2009)。本試驗中,試驗Ⅲ組降低了血漿谷氨酰胺、賴氨酸、甲硫氨酸濃度。說明補喂KIC可能會影響伊犁馬對某些氨基酸的吸收。賴氨酸和甲硫氨酸分別作為第一、第二限制性氨基酸,血漿賴氨酸和甲硫氨酸濃度降低,說明KIC維持了與限制性氨基酸的平衡,增加了伊犁馬對限制性氨基酸的吸收利用。血漿精氨酸濃度降低,推測血漿中的精氨酸可能是參與了機體其他氨基酸的合成。研究發現,通過給仔豬靜脈滴注400 μmol/kg·h的 KIC可降低血漿精氨酸、谷氨酰胺濃度(Escobar等,2010);Duan 等(2019)也證實了仔豬補喂1.25%的KIC-Ca可降低血漿中精氨酸濃度,與本試驗結果一致。因此,KIC可能促進了機體蛋白質或氨基酸的利用率。腦中游離色氨酸濃度的變化與支鏈氨基酸濃度有關,因為支鏈氨基酸能競爭抑制色氨酸在血腦屏障上的載體——白蛋白通道(Leite等,2010)。色氨酸是5-羥色胺的前體物質,大腦吸收過多血液運輸傳遞的游離色氨酸會引起腦內5-羥色胺增加,從而引起中樞疲勞(Davis等,2000)。本試驗中血漿5-羥色胺濃度顯著下降。說明KIC可能在抑制機體中樞疲勞方面具有積極作用。本試驗中,各組血漿天冬氨酸、天冬酰胺和精氨酸濃度的變化呈下降趨勢,但試驗Ⅱ組相比其他各試驗組血漿天冬氨酸、天冬酰胺和精氨酸濃度較高。關于補喂不同水平的KIC對氨基酸代謝的影響有待進一步研究。
4 結論
本試驗結果表明,補喂α-酮異己酸提高了伊犁馬血漿中α-酮異己酸濃度,且各試驗組均在補喂后1 h達到峰值;提高了血漿亮氨酸濃度;降低了血漿中支鏈氨基酸和5-羥色胺的濃度。通過補喂不同水平的α-酮異己酸,改變了伊犁馬對某些氨基酸的吸收利用。
猜你喜歡
現代臨床醫學(2022年4期)2022-09-29 07:38:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
火花(2019年12期)2019-12-26 01:00:28
云南醫藥(2019年3期)2019-07-25 07:25:14
人大建設(2019年12期)2019-05-21 02:55:32
海南醫學(2016年8期)2016-06-08 05:43:00
學苑創造·A版(2015年11期)2016-01-14 09:03:27
醫學研究雜志(2015年9期)2015-07-01 17:28:15
鄭州大學學報(理學版)(2014年3期)2014-03-01 04:21:05
