
蘇蓋特布拉克剖面微生物巖地層特征及沉積演化
2021-11-23 07:54:58楊兆偉郭睿婷
承德石油高等專科學校學報 2021年5期
昌 琪,郭 峰,尚 凱,楊兆偉,郭睿婷,韋 悅
(1.中國石化西北油田分公司 勘探開發研究院,新疆 烏魯木齊 830011;2.西安石油大學 地球科學與工程學院,陜西 西安 710065)
微生物碳酸鹽巖在中新元古界、寒武系和奧陶系最為發育,在美國Alabama、俄羅斯Siberi、巴西Santos地區、阿曼Salt盆地、哈薩克斯坦以及中國四川盆地和華北地區均有重大油氣發現[1-6]。目前微生物碳酸鹽巖也逐漸成為主要的目的層之一[7-11]。塔里木盆地柯坪地區肖爾布拉克組發育有廣泛分布且較為典型的微生物礁[12-21],但在盆地內目前未獲得大規模的油氣發現。塔里木盆地多口鉆井(塔中1、塔中23、方1等)均已證實該盆地良好的微生物礁型油氣藏勘探前景,但是對于微生物礁發育條件及典型微生物礁沉積模式尚缺乏系統認識。本次研究以塔西北寒武系典型剖面蘇蓋特布拉克高精度野外露頭實測為基礎,詳細刻畫微生物礁發育特征、顯微組構、沉積模式。該剖面微生物礁分布廣泛、代表性、可對比性上佳,前人對本區沉積儲層研究多以概述性為主[15-21],本次以中國石化西北油田分公司與西安石油大學共建野外教學實習剖面為基礎,按1 ∶100精度實測了該剖面,并樹立了耐久性、明顯可見的標識。研究成果對塔西北寒武系微生物碳酸鹽巖勘探具有一定的指導意義。
1 地層特征
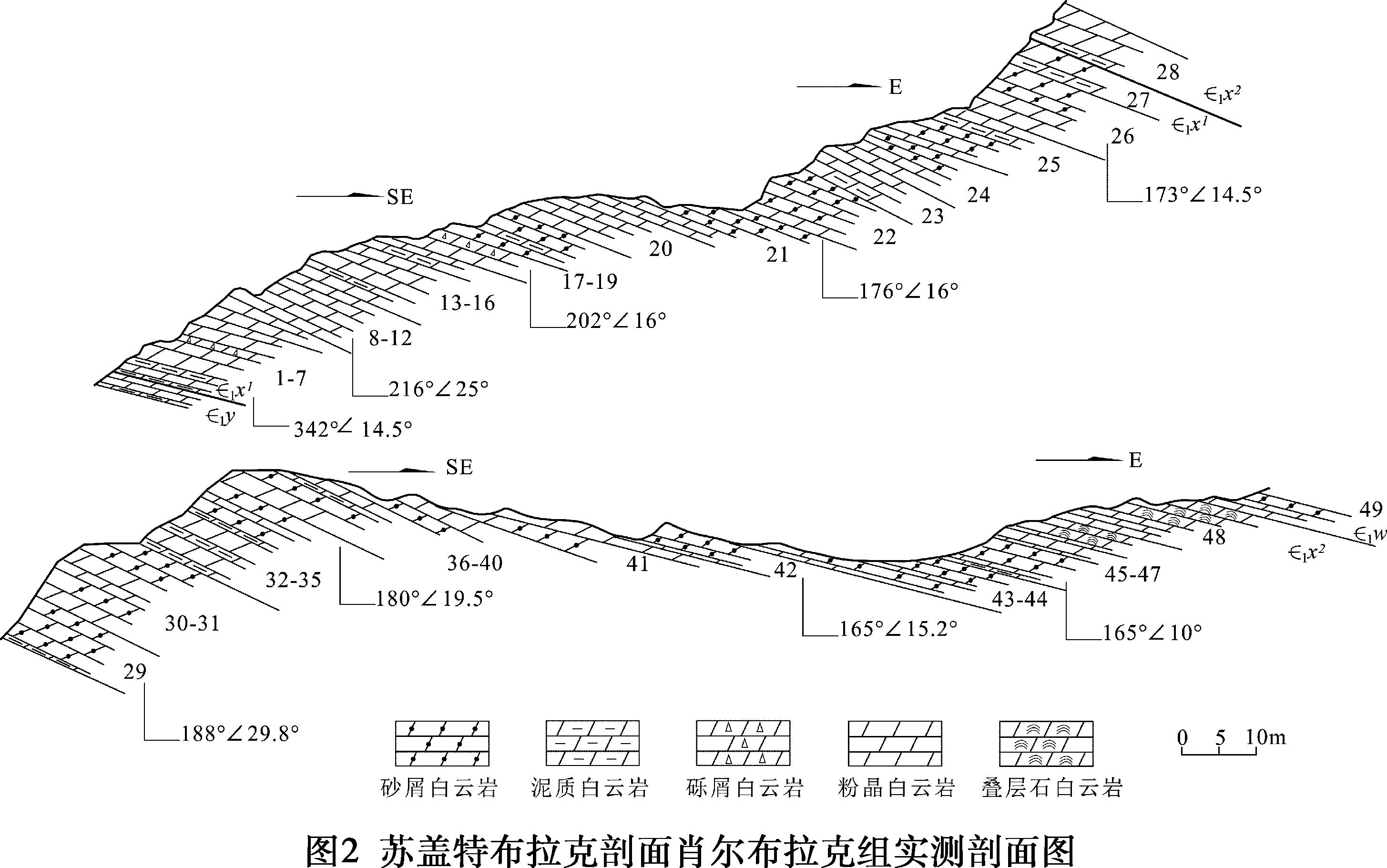
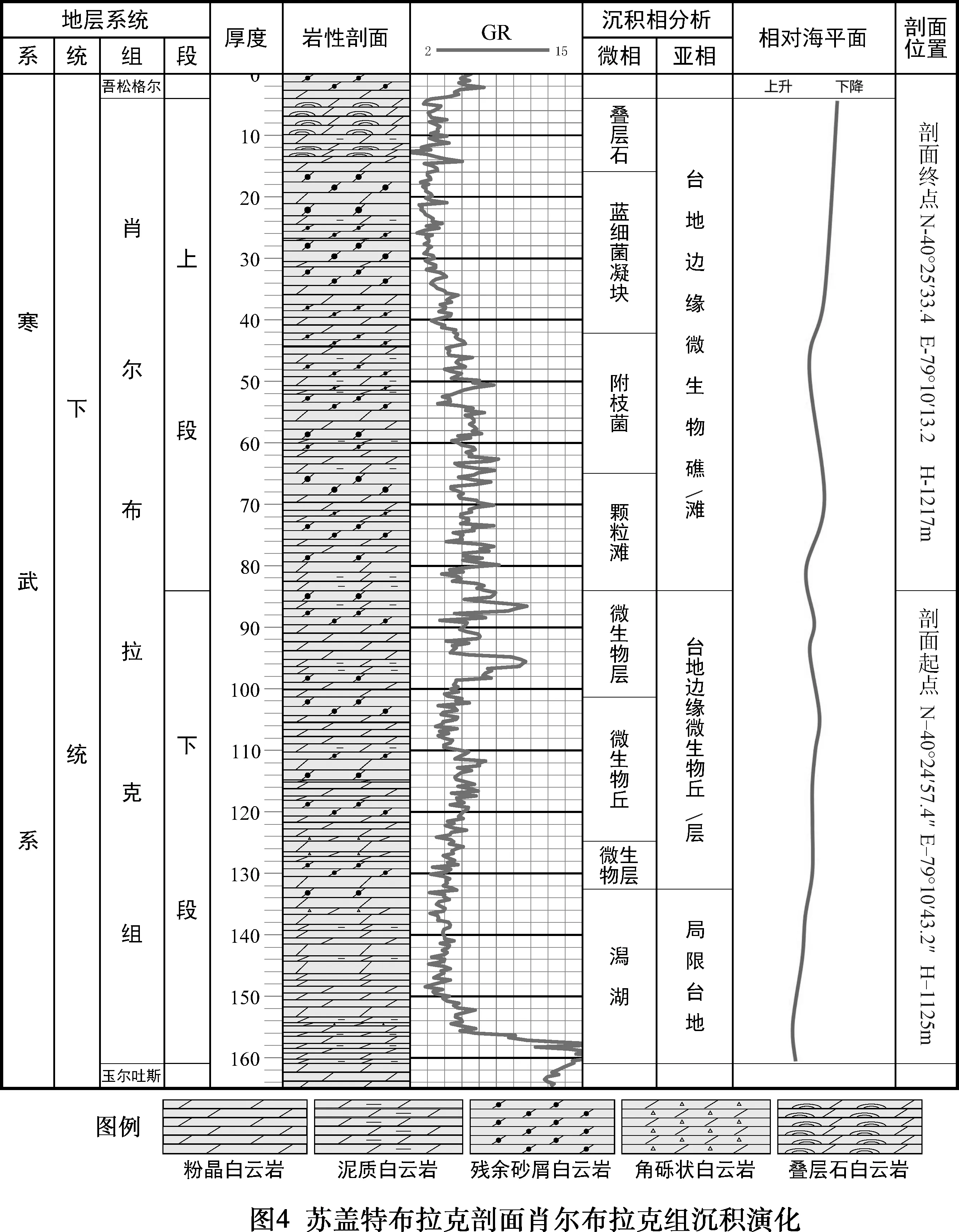
關于蘇蓋特布拉克剖面(見圖1),多個單位先后都進行過不同程度的地質調查和地層古生物工作,特別是1974年新疆地質局區測大隊柯坪地區地層組,為本次地層分析提供了重要基礎(見表1)。本次研究按1 ∶100實測該剖面,真厚度156.96 m,并根據巖性及沉積環境特征將肖爾布拉克組分上、下兩段。下段(1~27層,厚度76.87 m):巖性主體為灰色中層狀粉晶白云巖、殘余砂屑白云巖,富含硅質條帶或團塊,屬于局限臺地潟湖及臺地邊緣微生物丘沉積。上段(28~48層,厚度80.09 m):微生物礁灘發育層段,灰色中厚層狀殘余砂屑白云巖、泥晶白云巖,常見黑色硅質結核或條帶,屬于臺地邊緣生物礁(灘)環境(見圖2)。
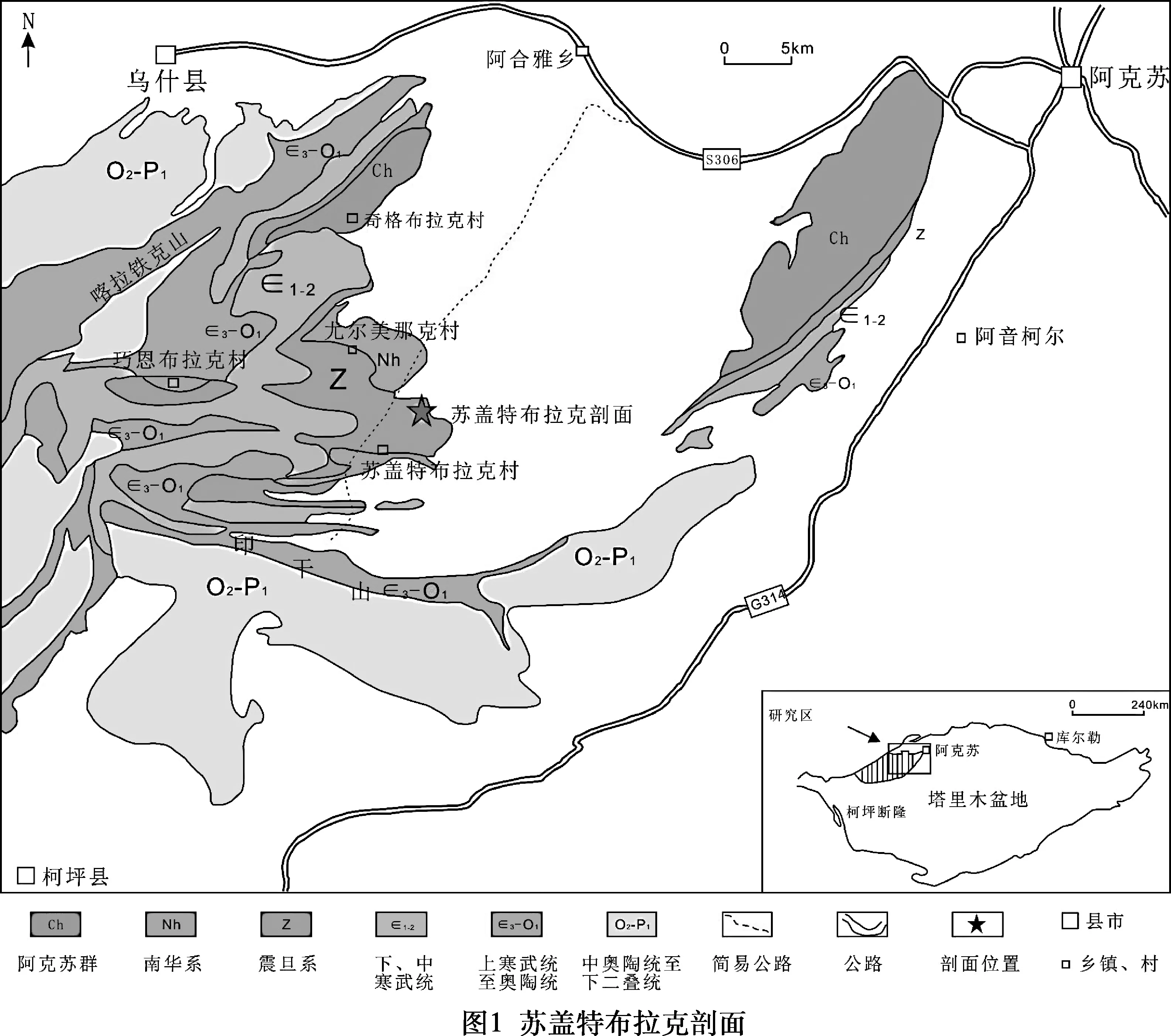
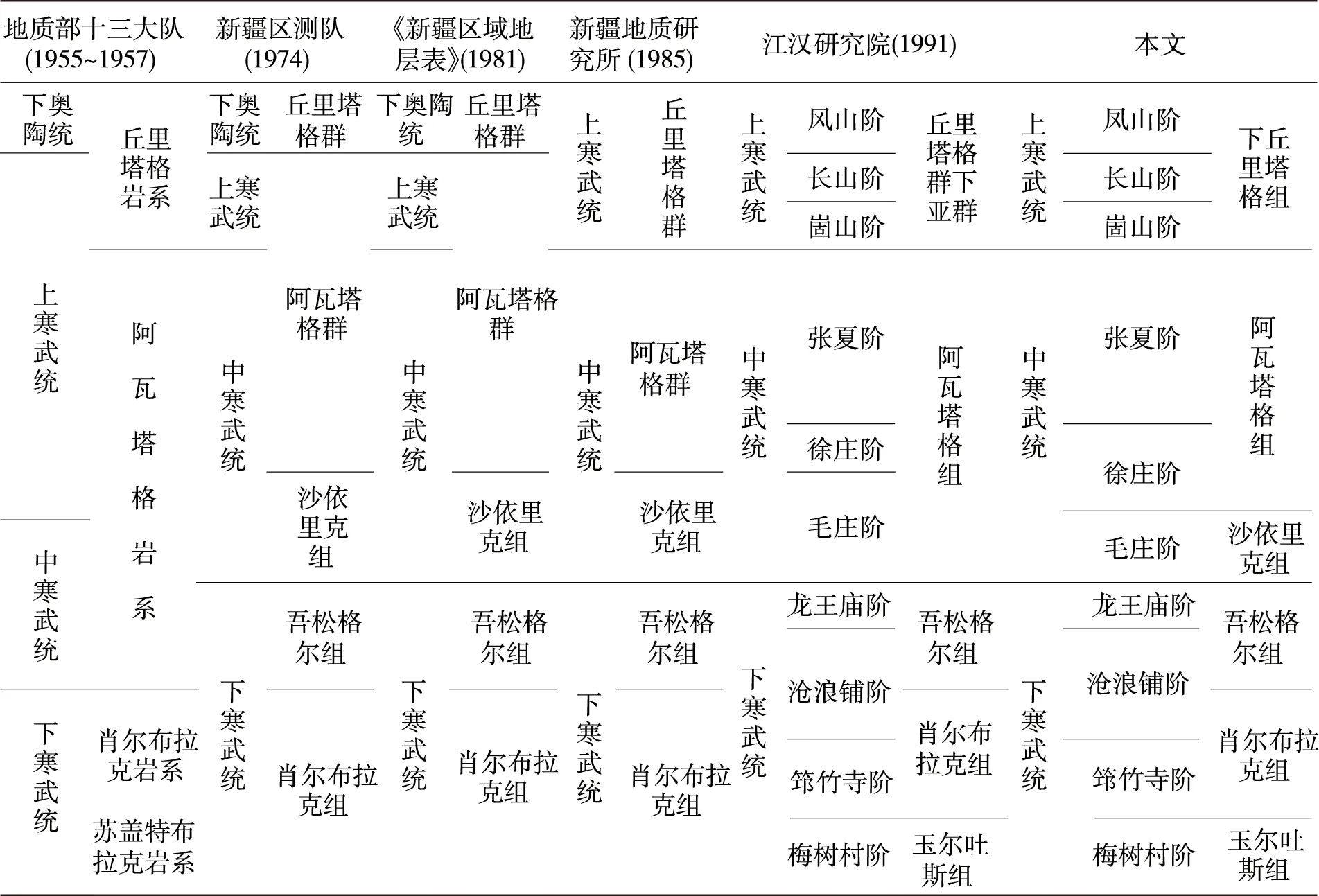
表1 柯坪地區寒武系劃分沿革表
在野外或鏡下可見到三葉蟲、腕足類、小殼化石,尤其是三葉蟲,分兩個三葉蟲帶:Shizhudiscus帶和Ushbaspis(=Metaredlichioides)帶。近底部Shizhudiscus(石柱盤蟲)帶,層位穩定,是與下伏玉爾吐斯組分界的標準化石,主要見于揚子區早寒武世早期。
2 沉積特征
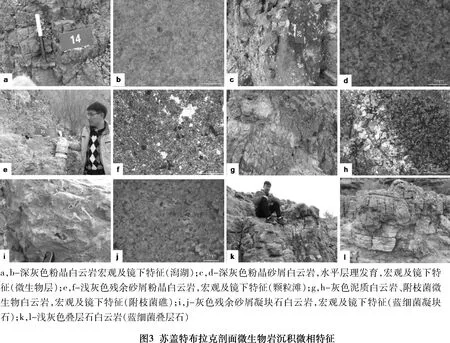
柯坪地區寒武系主要為海相碳酸鹽巖沉積,奧陶系時期塔里木盆地經歷了發育史上最大的一次海侵。通過對露頭資料以及取樣分析的綜合分析認為,塔里木盆地蘇蓋特布拉克剖面肖爾布拉克組屬于碳酸鹽巖臺地沉積(見表2、圖3),其中,下段相對于上段水體稍深,巖性主體為灰色中薄層狀粉晶白云巖及殘余砂屑白云巖,富含硅質條帶或團塊,下部為屬于局限臺地潟湖,上部為臺地邊緣微生物丘沉積。上段總體色淺,為微生物礁灘發育段,巖性在橫向上有變化,在東北部的礁主體剖面上,主要為粉晶白云巖、殘余凝塊粉晶白云巖,含附枝菌;在西南部則為灰色薄層狀泥晶白云巖,見硅質結核,屬于臺地邊緣生物礁(灘)環境。
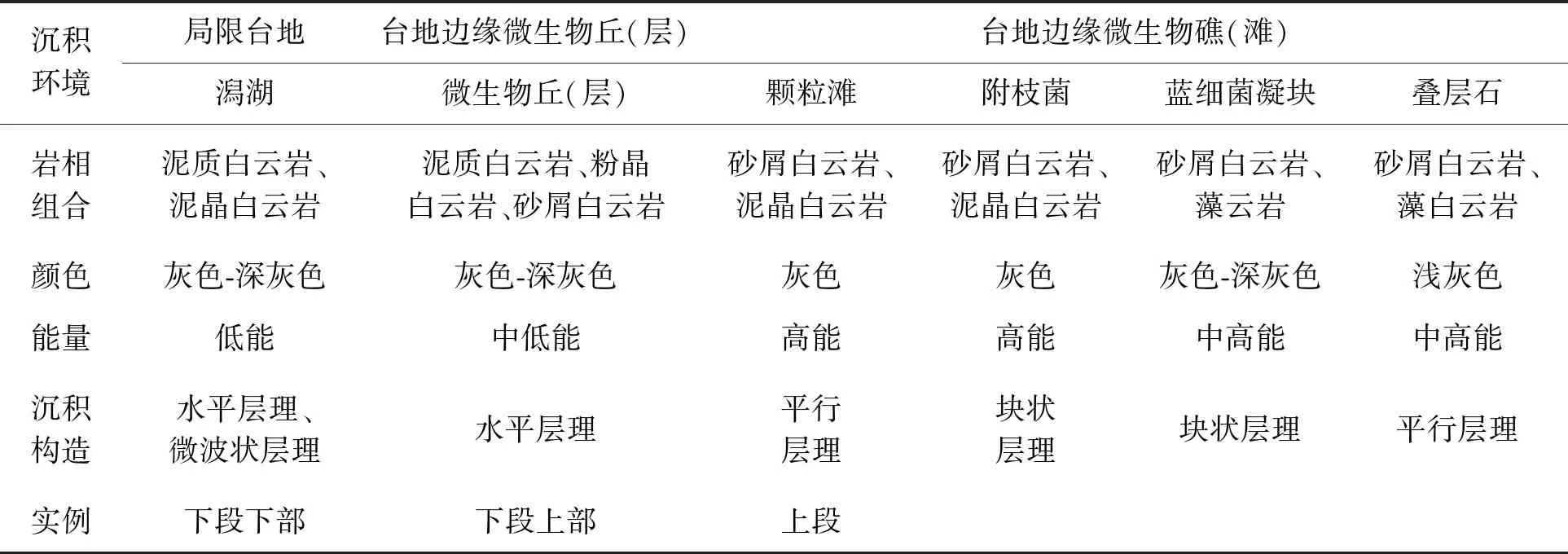
表2 蘇蓋特布拉克剖面肖爾布拉克組沉積微相類型及特征
2.1 潟湖
潟湖發育于肖爾布拉克組下段下部(1-18層),巖性主要為灰色泥晶白云巖夾泥質白云巖,水平層理發育,可見微波狀紋層,局部見硅質條帶,反映水體相對較弱的沉積環境(見圖3a~圖3b)。
2.2 臺地邊緣微生物丘(層)
肖爾布拉克組下段上部(19-27層)為微生物丘主要發育段,巖性以灰色粉晶白云巖夾殘余砂屑白云巖為主,局部發育波狀紋層,可見黑色硅質結核或條帶(見圖3c~圖3 d)。
2.3 臺地邊緣微生物礁(灘)
肖爾布拉克組上段是微生物礁(灘)主要發育層位。造礁生物主要為藍細菌類。
顆粒灘(28~31層):深灰色殘余砂屑粉晶白云巖夾泥質白云巖為主。砂屑以中細砂屑為主,細粉晶結構,構成微生物礁的基底。泥粉晶白云巖多發育水平層理。鏡下可見暗色顆粒結構,粒徑約0.1~0.3 mm,整體呈似球形或似橢球狀,主要為磨圓的泥晶碎屑或生物球粒(見圖3e~圖3f)。
附枝菌礁(32~38層):藍細菌為鈣化藍細菌,且有明顯密集的分枝特點的附枝菌。主要為灰色殘余砂屑粉晶白云巖(單層厚度大,0.5~1.5 m)與薄層狀泥晶白云巖(單層厚度小,0.1~0.4 m)不等厚互層,有兩期微生物礁發育。
附枝菌呈現樹枝狀和簇球狀兩種形態:其一呈分枝狀或叢狀,為縱切附枝菌,這種結構呈現出孤立分叉的葉枝狀體,單個枝狀體長0.2~1.1 mm,寬0.15~0.7 mm,顏色較深,內部不發育微晶結構,由隱晶質白云巖組成,暗色葉枝狀體間發育的孔隙多被亮晶白云巖充填;葉枝狀體形狀類似小灌木叢。其二態種形是呈團簇狀,為附枝菌的橫切面,即附枝菌葉狀體橫斷面的圓形斑點簇,圓形斑點的內部不發育其他微細結構。這種微生物結構在附枝菌為主要部分。團簇狀附枝菌生長形態表現為排列密集的圓形斑點,略呈放射狀。團簇體的直徑0.3~1.2 mm不等,每個團簇體內部發育一個不規則形狀的偏心空隙(見圖3 g~圖3 h)。團簇體內部發育多個暗色斑點,呈圓形-橢圓形,為附枝菌葉狀體的橫斷面。
宏觀上原地生長的附枝菌礁上覆于似顆粒灘之上,附枝菌微生物礁宏觀上表現為似層狀微生礁建造,以點礁形式生長,呈凸起形狀,指示水深較淺,水動力條件較高的臺地邊緣沉積環境。
藍細菌凝塊石(39~45層):凝塊石微相發育在原地生長的附枝菌礁之上,呈暗色斑塊狀,主要有中層狀和塊狀兩種類型(見圖3i~圖3j)。中層狀凝塊結構微生物巖(39~41層)從下到上巖層厚度略有增加,無明顯的丘狀構造,微觀下此類微生物巖為半自形細粉晶白云巖。塊狀凝塊結構微生物巖(42~45層)是以深灰色為主的殘余砂屑粉晶白云巖組成,凝塊石層厚約30~50 cm,在巖石表面有不規則的黑色團塊,團塊大小約(0.5~1.5)cm((0.5~0.8)cm分布不均勻且無明顯的定向性。
藍細菌疊層石(46~48層):藍細菌疊層石主要發育在本剖面的頂部,宏觀上呈厚層塊狀灰白色產出,厚11.93 m(見圖3k~圖3l)。
枝狀或叢狀附枝菌群常生長于礁體內部,在礁體外緣,附枝菌受到較強的水動力破壞作用,大多不能正常生長,而呈半閉合房室狀或凝塊狀生長,以抵抗水動力侵蝕。故上段附枝菌→凝塊石→疊層石的沉積演化也說明微生物礁的生長是一個水動力條件逐漸增強的過程。
3 沉積演化
塔里木盆地在早寒武世早期從盆內向東北、西南方向依次發育緩坡和盆地沉積。肖爾布拉克時期緩坡向臺地環境轉化,在海退背景下臺地范圍不斷向外擴展。
在海退大環境下,蘇蓋特地區肖爾布拉克組沉積水體也呈現向上變淺的規律,與下段底部發育一套泥質白云巖、泥晶白云巖,向上為可見紋層結構的殘余砂屑粉晶白云巖,該巖相組合特征反映了沉積環境從水動力較弱、水體較深的局限臺地相逐漸過渡為臺地邊緣微生物丘(層)沉積相;上段發育多期前積的微生物礁,反映海平面位置發生周期性變化(見圖4),層狀凝塊結構沉積水體較深,向上呈現為塊狀,反映水深變淺;頂部疊層石的出現指示其沉積環境由能量較低、水體較深沉積環境逐漸過渡為水動力高能的相帶沉積。故蘇蓋特布拉克剖面呈現為肖下段潟湖→臺地邊緣微生物丘(層)→肖上段顆粒灘→附枝菌礁→凝塊石→疊層石的演化,表明該剖面微生物礁的生長于一個水體能量逐漸增強,海平面不斷下降的沉積環境中(見圖5)。

4 結束語
隨著國內外碳酸鹽巖油氣勘探開發進程的日益加快,多學科綜合研究業已成為解決微生物礁與油氣分布相關問題的主要途徑,并萌芽了“生物礁地質學”。全球微生物碳酸鹽巖油氣發現主要集中在前寒武的非洲、阿拉伯板塊,印度、巴基斯坦和澳大利亞,以及西伯利亞的里菲系[22-23]。大規模微生物巖儲層多位于下古生界-前寒武系,與生命演化進程相對應。塔里木盆地中奧陶世以后,微生物在碳酸鹽沉積上起次要地位,生物建造主要是個體較大的骨架生物(如托盤類、藻類)。塔里木盆地、四川盆地、渤海灣盆地以及鄂爾多斯盆地的深部海相碳酸鹽巖油氣層將是以微生物碳酸鹽巖儲層為主角[10,11,14,22]。雖然建設性的成巖作用和構造破裂可以改善儲層性質,但對于微生物碳酸鹽巖沉積構造和組構是影響儲層發育的基本要素。盡管地質學家們對微生物碳酸鹽巖研究持續近兩百年,但其研究方興未艾[23]。
