Exendin-4對hIAPP轉基因鼠胰島功能的改善作用
2022-07-11 07:59:38劉辰鄒慶寶趙濟全
中國醫藥科學 2022年11期
劉辰 鄒慶寶 趙濟全



[摘要]目的探究Exendin-4對hIAPP轉基因鼠的保護作用及其可能的作用機制。方法正常小鼠和hIAPP轉基因鼠分別注射PBS和Exdendin-4,共分為四組:正常小鼠對照組(WT+PBS)、正常小鼠注射Exendin-4組(WT+Ex-4)、hIAPP小鼠對照組(homo+PBS)、hIAPP小鼠注射Exendin-4組(homo+Ex-4),每組6只。對四組小鼠進行2個月每天一次的腹腔注射,給藥結束后進行葡萄糖耐受實驗;體外培養Min6胰島β細胞,取對數生長期細胞分別加入2.5、5、10、20μM的hIAPP完全培養基,檢測細胞周期相關蛋白的表達;分離小鼠原代胰島細胞,AnnexinV-FITC檢測細胞凋亡;提取小鼠胰腺組織蛋白,檢測周期相關蛋白表達水平變化。結果homo+PBS組小鼠葡萄糖耐受能力顯著降低,差異有統計學意義(P<0.05),hIAPP體外處理Min6細胞會劑量依賴式下調ABC蛋白及周期相關蛋白CCND2和CCND3的蛋白表達。相反,homo+Ex-4組的小鼠葡萄糖耐受能力顯著增強,差異有統計學意義(P<0.05),原代胰島細胞的凋亡比例明顯降低,細胞周期蛋白CCND2和CCND3的表達水平增加。結論Exendin-4對hIAPP轉基因鼠造成的胰島損傷具有保護作用,其機制與調控胰島細胞增殖和凋亡有關。
[關鍵詞]Exendin-4;胰島淀粉樣多肽;細胞增殖;細胞凋亡
[中圖分類號]R587.1
[文獻標識碼]A
[文章編號]2095-0616(2022)11-0027-04
2型糖尿病胰島β細胞病變的主要特點是β細胞質量和功能減退而不能分泌足夠的胰島素[1]。胰島淀粉樣蛋白沉積是導致胰島β細胞功能逐漸喪失的重要因素[2]。淀粉樣蛋白的獨特成分是胰島淀粉樣多肽(islet amyloid polypeptide,IAPP),人源胰島淀粉樣多肽(human IAPP,hIAPP)是由胰島β細胞合成的具有37個氨基酸的短肽。hIAPP的過度沉積會導致胰島β細胞的凋亡并損傷葡萄糖代謝,從而導致2型糖尿病的發生[3]。Exendin-4含有39個氨基酸殘基,屬于GLP-1受體長效激動劑。研究發現Exendin-4可以通過多種途徑改善胰島β細胞的功能,起到治療糖尿病的作用[4]。然而,這種作用是否發生在由內源性hIAPP形成的淀粉樣蛋白沉積的條件下尚不清楚,本研究旨在探討Exendin-4在體內對hIAPP轉基因鼠的保護作用及其可能的分子機制。
1材料與方法
1.1材料
hIAPP轉基因小鼠購自JacksonLab;血糖儀和血糖試紙購自三諾生物傳感公司;hIAPP購自肽谷生物科技有限公司;Exendin-4購自MCE公司;小鼠Min6胰島β細胞由本實驗室保存;胎牛血清、DMEM培養基購自Gibco公司;AnnexinV-FITC/PI雙染凋亡檢測試劑盒購自凱基生物公司;ABC和β-catenin抗體購自CST公司;CCND2、CCDN3和CDK4購自ABclonal公司;β-Actin抗體購自Proteintech公司;PVDF膜、ECL發光液購自Millipore公司。
1.2方法
1.2.1小鼠Exendin-4處理將6~8周齡雄性小鼠分為四組:正常小鼠對照組(WT+PBS)、正常小鼠注射Exendin-4組(WT+Ex-4)、hIAPP轉基因小鼠對照組(homo+PBS)、hIAPP轉基因小鼠注射Exendin-4組(homo+Ex-4)。Exendin-4按照10nmol/kg的劑量給藥。對四組小鼠進行2個月每天一次的腹腔注射,給藥結束后進行葡萄糖耐受實驗(oral glucose tolerancetest,OGTT)。
1.2.2小鼠葡萄糖耐受實驗實驗前先將小鼠禁食16h,而后檢測小鼠血糖水平作為起始血糖值,然后,對小鼠按照2g/kg體重的標準進行葡萄糖腹腔注射,分別在30、60、120min尾靜脈取血測定血糖水平。
1.2.3小鼠胰腺組織取材每組取3只小鼠用3%的戊巴比妥鈉麻醉后,打開胸腔,將灌流針刺入心尖,先用生理鹽水進行沖洗,然后灌注4%的多聚甲醛,當肝臟發白時灌流結束。迅速摘取胰腺組織于-80°C保存,用于蛋白檢測。
1.2.4小鼠原代胰島細胞分離每組取3只小鼠頸椎脫臼處死后用酒精浸泡消毒。在生物安全柜中打開腹腔,十二指腸的開口處灌注Hank’s鹽,當胰腺膨脹后,摘除胰腺組織裝入盛有預冷Hank’s鹽的平皿中,剪成1~3mm的小塊,在37°C培養箱中消化15min后加入預冷Hank’s鹽中止消化,1500r/min室溫離心3min洗滌3次,用PBS重懸后置于冰上用于后續流式細胞儀檢測。
1.2.5Annexin V-FITC檢測細胞凋亡將分離好的小鼠胰島細胞加入5μl Annexin-V-FITC和10μl碘化丙啶(PI)試劑,室溫避光孵育20min,隨后立即進行流式細胞儀檢測,Annexin V-FITC為綠色熒光,碘化丙啶(PI)為紅色熒光。
1.2.6Westernblot檢測相關蛋白表達RIPA裂解液裂解細胞,BCA法進行蛋白定量。SDS-PAGE凝膠電泳進行蛋白分離,半干法轉運至PVDF膜,室溫封閉1h,加入一抗抗體4°C孵育過夜。TBST洗滌后,二抗室溫孵育1h,加入ECL發光液,使用天能成像系統進行檢測。
1.3統計學方法
采用SPSS19.0統計學軟件進行數據處理,計量資料用均數±標準差(x±s)表示,采用t檢驗,P<0.05為差異有統計學意義。
2結果
2.1Exendin-4提高hIAPP小鼠的葡萄糖耐受能力
腹腔注射葡萄糖后,各組小鼠的血糖水平均升高。相比WT+PBS組,homo+PBS組上升更為顯著,差異有統計學意義(P<0.05),而homo+Ex-4組血糖水平相比homo+PBS組明顯降低,差異有統計學意義(P<0.05),隨著時間的增加,WT+PBS組的血糖幾乎恢復到正常水平,而homo+PBS組仍較高,homo+Ex-4組血糖水平迅速下降。見表1。
2.2hIAPP抑制胰島β細胞周期相關蛋白的表達
不同濃度的hIAPP體外刺激小鼠Min6胰島β細胞48h。Western blot結果顯示,hIAPP可以明顯抑制ABC、CCND2以及CCND3的蛋白表達,并且呈現劑量依賴,而CDK4和β-catenin的蛋白表達水平無明顯變化,見圖1。
2.3Exendin-4抑制hIAPP轉基因小鼠的胰島細胞凋亡
小鼠原代胰島細胞凋亡分析結果顯示,相比于WT+PBS組(1.43%),homo+PBS組的早期凋亡細胞數明顯增加(21.3%),而homo+Ex-4組可以明顯降低由hIAPP導致的原代胰島細胞凋亡比例(9.76%),WT+Ex-4組無明顯變化,見圖2。
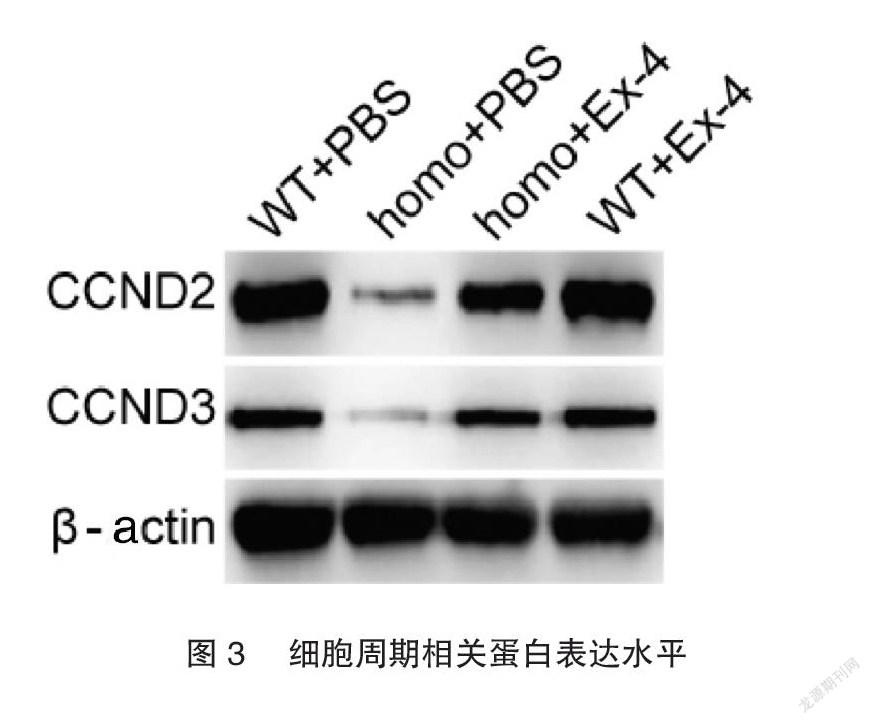
2.4Exendin-4促進hIAPP轉基因小鼠的胰島細胞周期相關蛋白表達
小鼠原代胰腺細胞Western blot結果顯示,homo+PBS小鼠原代胰島細胞中CCND2和CCND3的蛋白表達顯著降低,而Exendin-4可以逆轉由hIAPP導致的CCND2和CCND3的降低,見圖3。
3討論
2型糖尿病的主要特征是胰島β細胞的功能下降所引起的慢性高血糖,通常伴有胰島素敏感性降低[5-6]。盡管β細胞的功能改變在糖尿病的發病中起到關鍵作用,但β細胞數量的減少同樣對糖尿病的病理進展具有重要影響[7]。與非糖尿病患者相比,2型糖尿病患者體內β細胞數量減少了40%~65%[8]。目前認為增殖能力降低且凋亡增多是造成β細胞數量減少的主要因素。在2型糖尿病患者的胰島組織中發現有大量的淀粉樣蛋白沉積,經鑒定發現其主要成分是IAPP[9]。胰島周圍高濃度的IAPP通過形成大量淀粉樣沉積導致β細胞的功能紊亂[2]。在動物模型中,過表達hIAPP能夠顯著增加淀粉樣蛋白沉積,導致β細胞功能障礙和凋亡[10-11]。本研究發現,hIAPP轉基因小鼠的葡萄糖耐受能力明顯減弱,表現出了糖尿病癥狀。
Exendin-4是一種葡萄糖濃度依賴性促胰島素分泌劑,可通過促進胰島β細胞的增殖、調控細胞自噬以及抑制細胞凋亡來改善胰島的功能[4,12]。但是Exendin-4是否在體內對hIAPP轉基因鼠具有保護作用尚缺乏研究。在本研究中,體內注射Exendin-4可以明顯增強hIAPP轉基因小鼠的葡萄糖耐受能力,提示Exendin-4具有改善IAPP誘導的細胞凋亡作用。
細胞周期是在眾多蛋白分子調控下最終產生兩個新的子細胞的過程[13],細胞周期蛋白(cyclin)通過與細胞周期依賴性激酶(cyclin-dependent kinases,CDKs)相互協作,共同維持和協調細胞周期的運行[14-15],細胞周期的正常運轉對于維持細胞數量和功能至關重要[16]。在本研究中,hIAPP在體外會明顯抑制小鼠β細胞中細胞周期相關蛋白CCND2和CCND3的表達,而對細胞周期依賴性激酶CCRK和CDK4無影響。在hIAPP轉基因小鼠原代胰島細胞檢測中發現,體內注射Exendin-4可以有效逆轉由hIAPP導致的周期蛋白CCND2和CCND3的表達丟失,提示Exendin-4可以促進體內胰島細胞增殖。另外,本研究還發現,hIAPP轉基因小鼠的胰島組織中凋亡細胞比例明顯增加,而Exendin-4處理組細胞凋亡比例顯著降低,說明Exendin-4具有抑制胰島細胞凋亡的功能。
綜上所述,Exendin-4可以恢復hIAPP轉基因小鼠的胰島功能,其作用機制可能與Exendin-4促進小鼠體內胰島細胞增殖并且抑制胰島細胞凋亡有關。本研究為Exendin-4的臨床應用提供了新的理論指導。
[參考文獻]
[1] TAYLOR R.Type 2 diabetes: etiology and reversibility[J]. Diabetes Care,2013,36(4):1047-1055.
[2] BISHOYI AK,ROHAM PH,RACHINENI K,et al. Human islet amyloid polypeptide (hIAPP) -a curse in type II diabetes mellitus: insights from structure and toxicity studies[J].Biological Chemistry,2021,402(2): 133-153.
[3] RALEIGH D,ZHANG X,HASTOY B,et al.The β-cell assassin: IAPP cytotoxicity[J].Journal of Molecular Endocrinology,2017,59(3):R121-R140.
[4] YAP MKK,MISUAN N.Exendin-4 from Heloderma suspectum venom: From discovery to its latest application as type II diabetes combatant[J].Basic Clin Pharmacol Toxicol,2019,124(5):513-527.
[5] MEZZA T,CINTI F,CEFALO CMA,et al.β-Cell Fate in Human Insulin Resistance and Type 2 Diabetes: A Perspective on Islet Plasticity[J].Diabetes,2019,68(6): 1121-1129.
[6] ?HAN JCN,LIM LL,WAREHAM NJ,et al.The Lancet Commission on diabetes: using data to transform diabetes care and patient lives[J].Lancet,2021,396 (10267):2019-2082.
[7] WEIR GC, GAGLIA J,BONNER-WEIR S.Inadequate β-cell mass is essential for the pathogenesis of type 2 diabetes[J].Lancet Diabetes Endocrinol,2020,8(3): 249-256.
[8] RAHIER J,GUIOT Y,GOEBBELS RM,et al.Pancreatic beta-cell mass in European subjects with type 2 diabetes[J]. Diabetes Obes Metab,2008,10 Suppl 4:32-42.
[9] MIYAZATO M,NAKAZATO M,SHIOMI K,et al. Identification and characterization of islet amyloid polypeptide in mammalian gastrointestinal tract[J]. Biochemical and Biophysical Research Communications, 1991,181(1):293-300.
[10] COSTES S,LANGEN R,GURLO T,et al.β-Cell failure in type 2 diabetes: a case of asking too much of too few?[J].Diabetes,2013,62(2):327-335.
[11] BLENCOWE M,FURTERER A,WANG Q,et al. IAPP-induced beta cell stress recapitulates the islet transcriptome in type 2 diabetes[J].Diabetologia, 2022,65(1):173-187.
[12] NAUCK MA,QUAST D R,WEFERS J,et al.GLP-1 receptor agonists in the treatment of type 2 diabetes- state-of-the-art[J].Mol Metab,2021,46:101102.
[13] LI X.Epigenetics and cell cycle regulation in cystogenesis[J].Cellular Signalling,2020,68: 109509.
[14] COLLER HA.Regulation of Cell Cycle Entry and Exit: A Single Cell Perspective[J].Compr Physiol,2019, 10(1):317-344.
[15] MARTíNEZ-ALONSO D,MALUMBRES M.Mammalian cell cycle cyclins[J].Semin Cell Dev Biol,2020, 107:28-35.
[16] INGHAM M,SCHWARTZ GK.Cell-Cycle Therapeutics Come of Age[J].J Clin Oncol,2017,35 (25):2949-2959.
(收稿日期:2021-12-23)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2022年5期)2022-08-24 02:35:42
中老年保健(2022年1期)2022-08-17 06:14:56
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年5期)2021-08-24 07:07:20
中老年保健(2021年11期)2021-08-22 03:15:16
學苑創造·A版(2020年9期)2020-10-13 09:41:02
海峽科技與產業(2016年3期)2016-05-17 04:32:12
