厚殼貽貝幾丁質提取及幾丁質相關基因篩查
2024-01-09 02:21:32袁俊杰李邁唐菊艷袁孟強李千千王曉娜張美溦陳偉王曉通
海洋通報 2023年6期
袁俊杰,李邁,唐菊艷,袁孟強,李千千,王曉娜,張美溦,陳偉,王曉通
(1.魯東大學 農學院,山東 煙臺 264025;2.煙臺市海洋經濟研究院,山東 煙臺 264003)
幾丁質是自然界中廣泛分布的一類生物大分子,由D-N-乙酰氨基葡萄糖通過β-(1,4)糖苷鍵聚合而成[1],根據結構不同,分為α-、β-和γ-幾丁質三種類型[2]。幾丁質通常不以單體形式存在,其常與蛋白質、色素、碳酸鈣等結合形成復合物[2],廣泛存在于軟體動物貝殼和內骨骼中,在貝殼形成和生物礦化中發揮重要作用。目前,已從烏賊、珠母貝、芋螺、江珧等軟體動物中成功提取并鑒定到幾丁質成分[3-7],研究發現軟體動物幾丁質具有良好的生物相容性、抗菌活性[7-8],在生物醫學等方面有重要的應用前景。
幾丁質作為軟體動物貝殼有機組分之一,與其他蛋白質、糖蛋白、多糖等物質組成有機框架,在此基礎上進行生物礦化[9]。其中,幾丁質合成酶(chitin synthetase,CHS)、幾丁質酶(chitinase,CHIA)、幾丁質結合蛋白(chitin-binding protein,CBP)等與幾丁質框架構建相關的蛋白,均是調控貝殼形成的重要蛋白,參與調控生物礦化[10-14]。在長牡蠣、紫貽貝和大扇貝的貝殼損傷研究中,三種雙殼貝類的貝殼修復后,外套膜差異表達蛋白質中均鑒定到幾丁質結合相關的結構域[15]。在紫貽貝貝殼修復和生物礦化的轉錄組分析中,也篩查到大量上調表達的幾丁質代謝相關基因,包括幾丁質合成酶(CHS)和幾丁質酶(CHIA)等[16]。
厚殼貽貝(Mytilus coruscus)是我國重要的海水養殖貝類,具有較高的經濟價值[17]。根據已有的研究結果:厚殼貽貝的貝殼包括珍珠質層、肌棱柱層和斜棱柱層[18],并通過蛋白質組學鑒定到60 余種殼基質蛋白[19],但幾丁質以及幾丁質相關基因在厚殼貽貝貝殼形成中的作用卻鮮有報道。本研究從形態和組織學兩方面詳細觀察了厚殼貽貝貝殼與外套膜結構,從厚殼貽貝的角質層和貝殼中提取并鑒定到幾丁質,并構建外套膜三褶皺轉錄組,根據轉錄組數據篩查到幾丁質合成、代謝、結合相關基因,并對三褶皺差異表達基因進行GO 富集分析,篩查出與幾丁質功能相關的GO term。本研究有助于了解厚殼貽貝貝殼中有機骨架的成分,為幾丁質在貝殼形成和生物礦化中的作用奠定研究基礎。
1 材料與方法
1.1 實驗樣品
實驗所用厚殼貽貝材料購于江蘇省連云港市,實驗前在海水養殖實驗室內充氧暫養,暫養海水水溫20 ℃,鹽度25‰,養殖期間飼喂螺旋藻粉,每天換水一次。
1.2 貝殼及外套膜的形態及組織學觀察
將腹緣端貝殼切下部分后,露出白色貝殼,并保留貝殼邊緣的角質層,觀察貝殼、角質層、外套膜三者生理學結構。將貝殼沿閉殼肌斷開,取腹緣端外套膜邊緣組織,經固定、脫水、透明、包埋后,在切片機(Leica,RM2255)上切成6 μm厚的切片。切片經脫蠟、復水后,用蘇木精-伊紅染液(南京建成,D006)染色,后經脫水、透明,用中性樹脂封片。最后,在顯微鏡(Nikon,CI-L)下觀察切片。
1.3 貝殼中幾丁質的提取
將厚殼貽貝貝殼刷洗干凈后,將表面的角質層剝離,烘干后研磨成粉末,放入2%鹽酸浸泡30 min,用蒸餾水充分漂洗,高速離心分離得到不溶物并吸干水分。向不溶物加入2 mol/L的NaOH溶液,室溫震蕩24 h,之后高速離心30 min,蒸餾水清洗至中性,過濾得到棕褐色不溶物。H2O2漂白后,蒸餾水漂洗。60 ℃烘干,得到固體粉末。
將角質層剝離后的貝殼清洗并烘干,也研磨成粉末,并放入2%鹽酸浸泡脫鈣,漂洗至pH 為中性后,高速離心,向沉淀物加入2 mol/L 的NaOH 溶液,室溫震蕩24 h。高速離心、洗滌、濾紙過濾,得到脫蛋白不溶物。脫色、漂洗、烘干,得到粉末狀固體。
1.4 幾丁質提取物的傅里葉紅外光譜鑒定
各取0.01 g 幾丁質提取物,在NICOLET IS50型傅里葉紅外儀器下,經32次全反射掃描在400~4 000 cm-1波數范圍內測得吸收峰,使用OMNIC軟件繪制吸收峰圖。
1.5 基于外套膜轉錄組的幾丁質相關基因鑒定
隨機選取3 只厚殼貽貝,分別采集外套膜邊緣的外褶皺、中褶皺、內褶皺組織,液氮速凍,放入超低溫冰箱保存。提取總RNA 后,抽提mRNA 構建RNA-seq 文庫并測序。原始數據經質量過濾后,通過Trinity 軟件進行De novo拼接,對組裝好的Unigene進行常見功能數據庫注釋,根據功能注釋和BLAST 比對,篩選出幾丁質合成、代謝、結合相關的基因,進行表達量(FPKM)分析,并對其蛋白序列進行功能域預測(http://smart.embl-heidelberg.de/)。對內、中、外褶皺間差異表達基因進行GO富集分析,篩查出與幾丁質合成、代謝、結合等功能相關的GO term。
2 實驗結果
2.1 厚殼貽貝貝殼及外套膜的組織特征
通過形態觀察,厚殼貽貝貝殼主要由角質層(貝殼硬質蛋白)和貝殼層(碳酸鈣晶體)兩部分組成(圖1(a)),而外套膜邊緣部分主要分為三個褶皺(圖1(b)、(c)),外褶皺(of)、中褶皺(mf)以及內褶皺(if),其中外褶皺形態在三個褶皺中最小,其邊緣緊緊貼附在貽貝的貝殼邊緣,內褶皺靠近其他軟組織部,且是三褶皺中形態最大的,其上分布大量外套膜觸手,而中褶皺則位于內、外褶皺中間,組織大小也介于二者之間。通過石蠟切片進一步觀察,中褶皺又分為一大一小2 層褶皺,靠近外褶皺一側有1 個非常小的褶皺,與外褶皺共同形成一個溝狀結構,此處可分泌貝殼的角質層膜(圖1(c)、(d1))。此外,根據組織切片觀察,發現三個褶皺的上皮細胞類型也有較大差別:殼膜溝處的外褶皺具有單層柱狀上皮細胞,其形態細長,細胞核細長位于基部,細胞質中存在少量褐色的色素(圖1(d1));而靠近殼膜一側的中褶皺(mf1 和mf2)其細胞形態為矮柱狀,細胞核橢圓形位于基部,這些上皮細胞中并未觀察到棕色的色素(圖1(d1)),而在遠離殼膜一側的中褶皺上皮細胞中則可觀察到大量棕色色素(圖1(d2));內褶皺也具有單層柱狀上皮,這些上皮細胞的核細長位于細胞基部,在胞質中可觀察到大量深棕色的色素(圖1(d3)、(d4))。
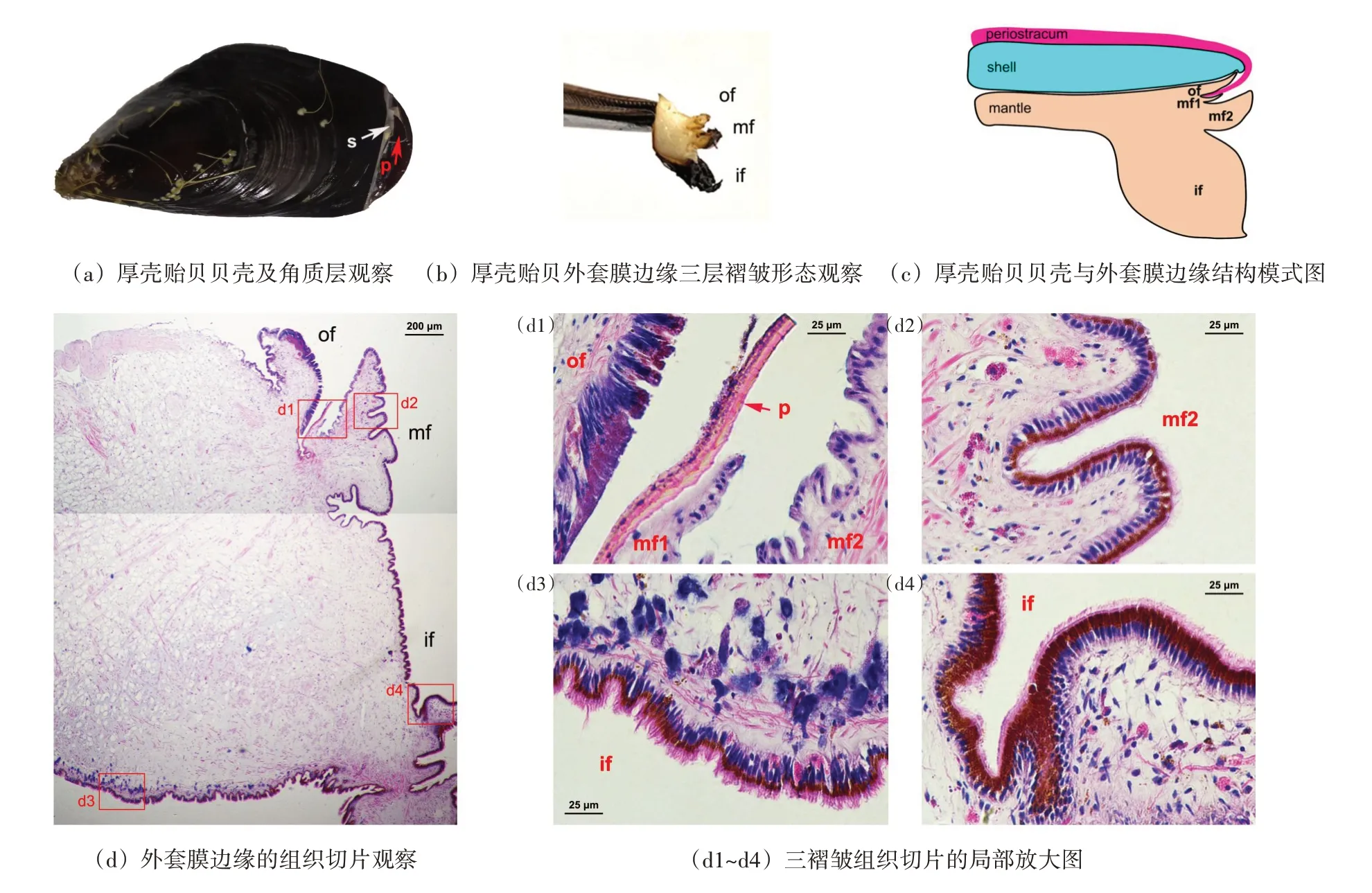
圖1 厚殼貽貝貝殼及外套膜的形態及組織學觀察
2.2 貝殼幾丁質提取及FTIR特征
從厚殼貽貝角質層粉末(圖2(a))中,提取并鑒定到幾丁質(圖2(b)):在 3 272 cm-1處有1個明顯的N-H 伸縮振動吸收峰,在2 800~3 000 cm-1處,有兩個C-H 伸縮振動吸收峰,在1 626 cm-1處有1 個較大的酰胺I 帶吸收峰,在1 516 cm-1處出現1個較大的酰胺Ⅱ帶吸收峰[6],在1 234 cm-1處出現1 個較小的酰胺Ⅲ帶吸收峰。此外,在1 430~1 445 cm-1具有CH2彎曲和CH3變形的特征吸收峰,在1 370~1 390 cm-1具有CH 彎曲與對稱CH3變形而出現特征吸收峰[20]。從貝殼粉末(圖2(c))中,也得到幾丁質提取物,并鑒定到幾丁質特征吸收峰(圖2(d)):在3 269 cm-1處具有N-H 伸縮振動吸收峰,在2 800~3 000 cm-1處,也出現兩個C-H伸縮振動吸收峰,在1 623 cm-1處出現一個酰胺I 帶吸收峰,在1 513 cm-1處有1 個酰胺Ⅱ帶吸收峰,并在1 560 cm-1處還出現一個較小的酰胺Ⅱ帶吸收峰,在1 232 cm-1處出現1 個較小的酰胺Ⅲ帶吸收峰。此外,在1 430~1 445 cm-1也具有CH2彎曲和CH3變形的特征吸收峰,在1 370~1 390 cm-1處也具有CH 彎曲與對稱CH3變形而出現特征吸收峰。
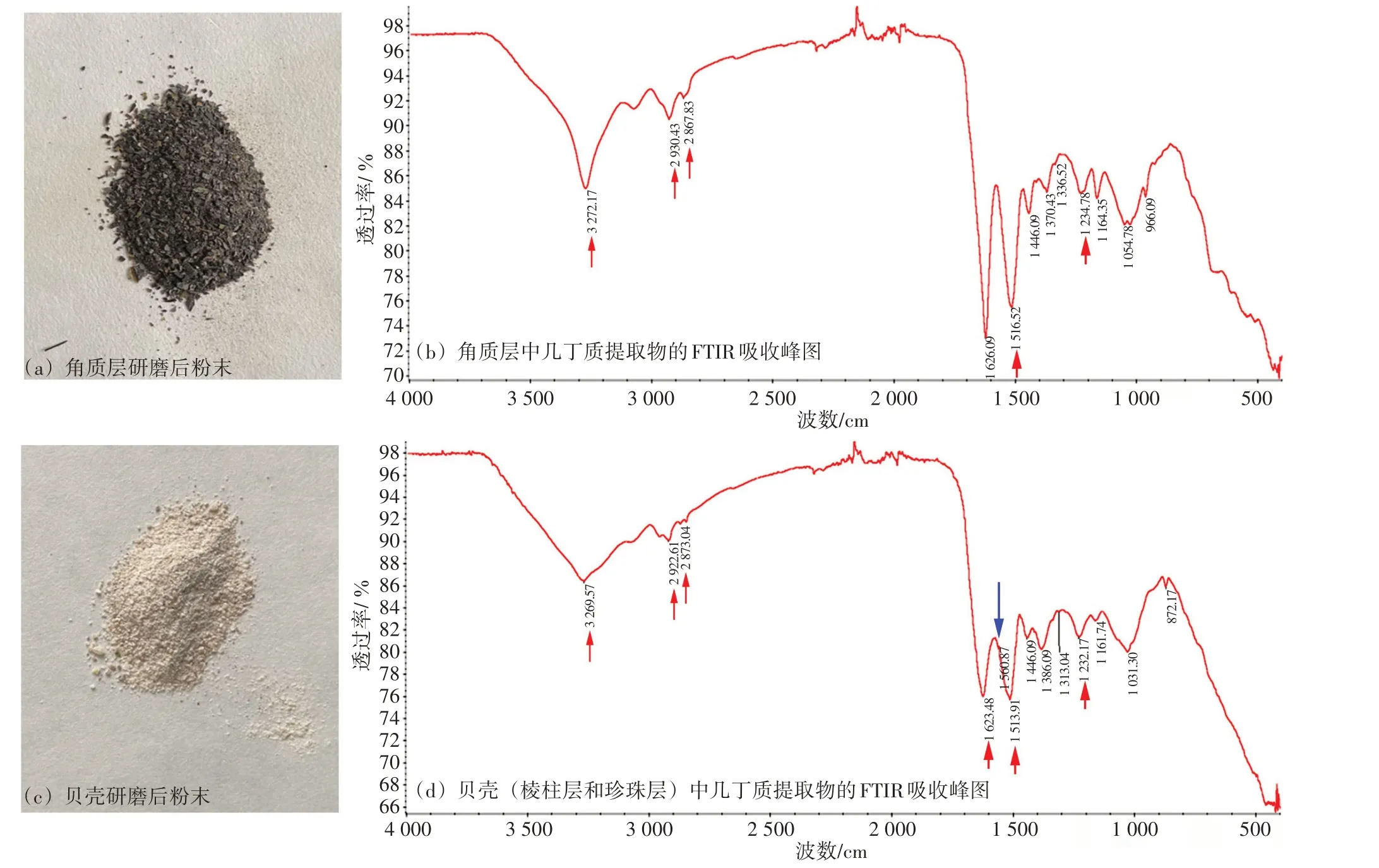
圖2 厚殼貽貝角質層和貝殼中幾丁質的提取及鑒定
2.3 幾丁質相關基因的篩查及表達分析
根據厚殼貽貝外套膜三褶皺的轉錄組數據,篩查出34 個幾丁質相關基因,其中幾丁質合成相關的基因有7 個(圖3(a)),幾丁質降解相關的基因有16 個(圖3(b)),幾丁質結合相關基因有11個(圖3(c))。根據功能域預測,7 個幾丁質合成酶基因(CHS)均具有chitn_synth_2 結構域。而篩查到的幾丁質酶主要包括18家族和20家族糖苷水解酶兩類,在16 個幾丁質酶中,有10 個具有Glyco_18 結構域(18 家族糖苷水解酶),其余6 個具有Glyco_hydro_20 結構域(20 家族糖苷水解酶)。其中McCHIA1、McCHIA4、chitotriosidase-1-like 還具有2 或3 個幾丁質結合結構域(ChtBD2);還 有4 個β 氨 基 己 糖 苷 酶(Bhexosaminidase)除了具有Glyco_hydro_20 結構域外,還具有一個特殊的碳水化合物結合域CHB_HEX結構域。厚殼貽貝11個幾丁質結合相關的基因,均具有1到5個ChtBD2結構域(圖3(c)),其中,Coadhesin 除了ChtBD2 結構域,還具備3 個血小板反應蛋白TSP1 結構域;Chitin binding/VWA1、Chitin binding/VWA2 還各自具備1 個與幾丁質結合相關的VWA 結構域;mucin-2-like 還具有1 個層粘連蛋白LamG 結構域;McCBP1 還具備1個凝血因子5/8 C-末端FA58C結構域。
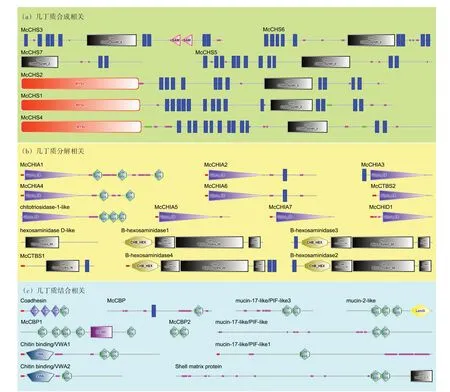
圖3 厚殼貽貝幾丁質相關基因的結構域預測
為進一步研究外套膜三褶皺與幾丁質相關功能的關系,對篩查到的幾丁質相關基因進行表達量分析(圖4),并在三褶皺差異表達基因的GO富集結果中篩查幾丁質相關的GO term(圖5)。根據三褶皺中幾丁質相關基因的表達量分析結果,發現幾丁質合成相關基因主要在內、中褶皺高表達;而幾丁質結合相關基因的表達模式則與之相反,主要在外褶皺高表達;篩查到的16 個幾丁質酶基因,則主要呈現兩種表達模式:一類是在外褶皺中高表達,一類在內、中褶皺中高表達。其中,在外褶皺中高表達的幾丁質酶為殼三糖苷酶(McCHIA),屬于18 家族糖苷水解酶;而在內、中高表達的幾丁質酶多為二-N-乙酰殼二糖酶(McCTBS)、β 氨基己糖苷酶(B-hexosaminidase)等。此外,從外套膜三褶皺差異基因的GO富集分析結果中,篩查出多個幾丁質相關的GO term(圖5),發現內、中褶皺的差異高表達基因主要富集到幾丁質生物合成相關的GO term,而外褶皺則主要富集到幾丁質結合、代謝、角質層色素沉著相關的GO term。
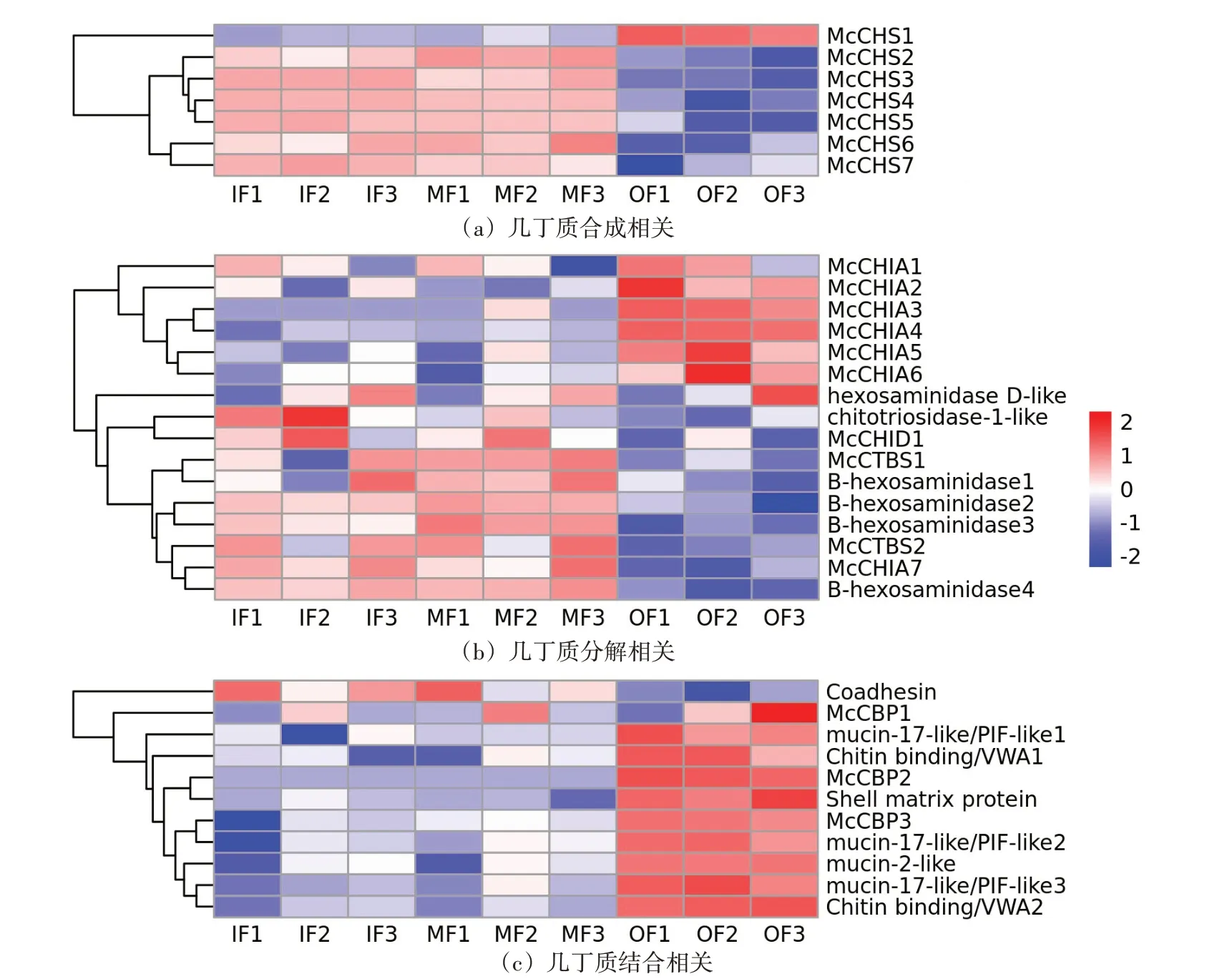
圖4 厚殼貽貝幾丁質相關基因在外套膜三褶皺中的表達分析
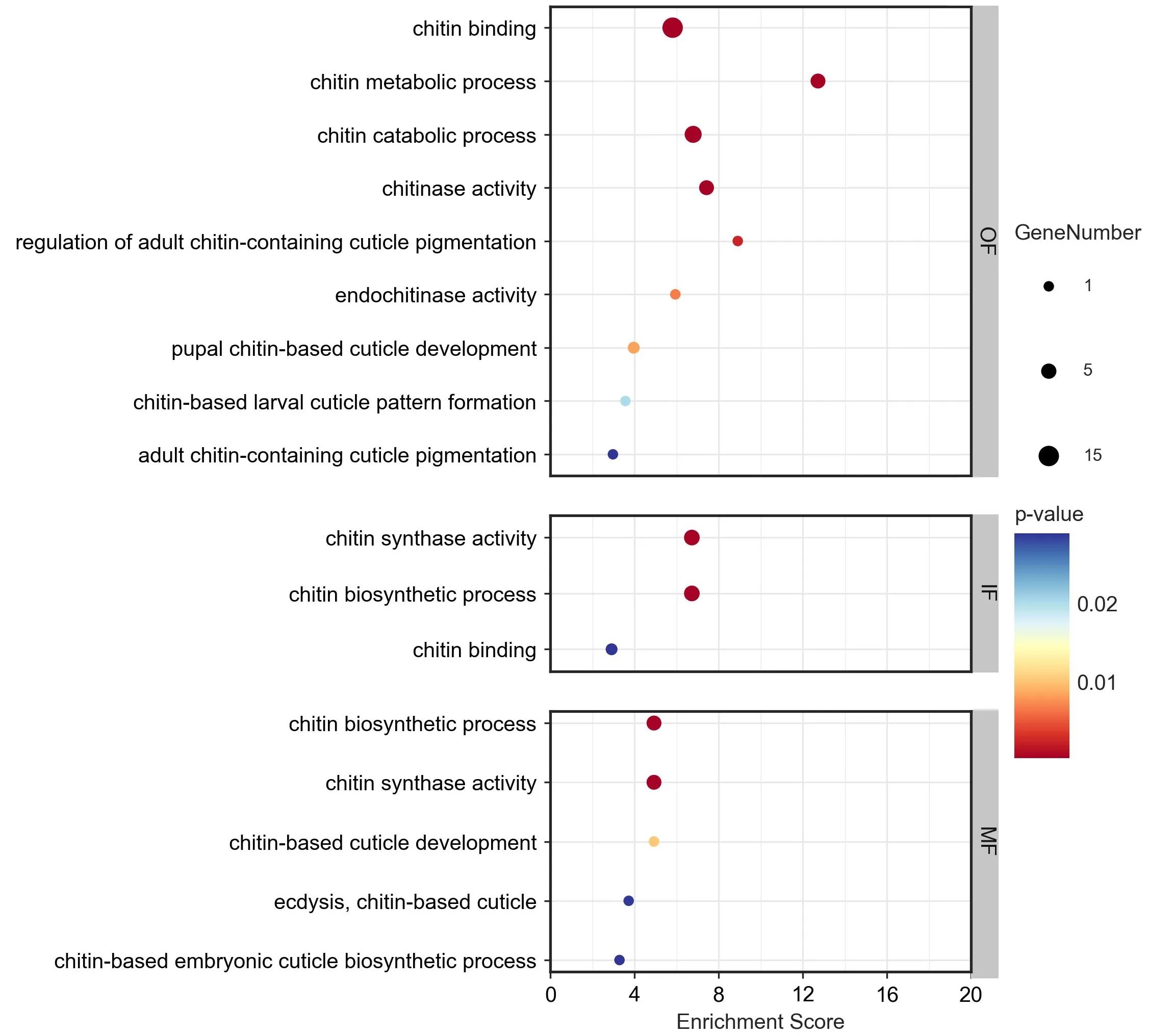
圖5 厚殼貽貝外套膜三褶皺中高表達基因的GO富集分析
3 討論
在自然界中,碳酸鈣晶體會嵌入以幾丁質為主成分的有機基框架中進行生物礦化,因此,本研究在提取幾丁質時,進行了脫鈣、脫蛋白等過程。根據FTIR 結果,本研究成功在厚殼貽貝的角質層和貝殼層提取到幾丁質,二者分別在3 272 cm-1和3 269 cm-1處均存在單一吸收峰,這表明貽貝角質層和貝殼層中提取的幾丁質均為β-幾丁質而非α-幾丁質[3-4,21]。但由于貝殼層與角質層中有機質和礦物質的構成存在差異,兩種幾丁質提取物的FTIR 吸收峰發生了一定程度的左右平移。此外,貝殼層中幾丁質提取物在1 560 cm-1處還多了一個較小吸收峰(圖2(d),藍色箭頭所示),二者在吸收峰的大小上,角質層提取物在1 513~ 1 626 cm-1處的吸收峰也大于貝殼層提取物,這表明,角質層中幾丁質含量要更高于貝殼層中幾丁質含量,且所含其他雜質也較少。幾丁質作為貝殼有機基質的重要組成,在生物礦化中具有關鍵調控作用,在牡蠣、珠母貝、芋螺等貝類貝殼中均鑒定到幾丁質的成分[5-7,21],且這些貝類貝殼中的幾丁質提取物均是具有β-幾丁質的特征吸收峰。
除幾丁質外,貝殼基質蛋白也是貝類貝殼有機基質的重要組成,此前,在長牡蠣[22]、貽貝[19]、海螂[23]、櫛江珧[24]、珠母貝[25]、櫛孔扇貝[26]等雙殼貝類的貝殼基質蛋白研究中,均鑒定到含有幾丁質結合結構域的蛋白質以及幾丁質代謝相關蛋白。在紫貽貝的研究中,也發現幾丁質在幼蟲外殼的形成和功能中發揮著重要作用[25],在翡翠貽貝貝殼蛋白質組分析中,還發現大量具有幾丁質結合結構域的蛋白質[27]。因此,探究幾丁質相關的殼基質蛋白有助于更好地了解厚殼貽貝貝殼有機骨架的形成。本研究利用外套膜邊緣三褶皺的轉錄組數據,篩查出厚殼貽貝中幾丁質合成、代謝、結合相關基因。在篩查到的三類幾丁質相關基因中,幾丁質酶數量最多。自然界中,幾丁質酶主要包括幾類糖苷水解酶(18、19、20、23、48 家族),而在厚殼貽貝中鑒定到了18 家族和20 家族兩類糖苷水解酶。結合這三類幾丁質相關基因在三褶皺中的表達量分析,發現這些幾丁質酶在三個褶皺中的表達模式有所區別:β 氨基己糖苷酶、二-N-乙酰殼二糖酶等幾丁質酶基因主要在內、中褶皺表達;而殼三糖苷酶類的幾丁質酶主要在外褶皺表達。此外,根據表達模式,發現幾丁質合成酶基因主要在內、中褶皺高表達,而幾丁質結合蛋白相關基因的表達模式卻與之相反,主要在外褶皺高表達。這表明外套膜三層褶皺在幾丁質相關的功能上可能存在差異。
此前,在珠母貝[12]、三角帆蚌[14]、長牡蠣[21]等雙殼貝類的研究發現幾丁質結合蛋白能與幾丁質和碳酸鈣結合,并指導碳酸鈣晶體的結晶并參與調控貝殼的形成。在長牡蠣、紫貽貝和大扇貝的貝殼損傷研究中,貝殼修復后外套膜差異表達的蛋白質中,均發現具有幾丁質結合相關的VWA 結構域、LamG 結構域[15-16],這些研究表明具有幾丁質結合相關的VWA 結構域、LamG 結構域蛋白能夠參與貝殼損傷后的修復。結合在厚殼貽貝中的研究結果,幾丁質結合蛋白主要在厚殼貽貝在外褶皺高表達,在這些外褶皺高表達蛋白中也發現幾丁質結合相關的VWA 結構域、LamG 結構域的存在,由此推測,厚殼貽貝可能通過外褶皺調控幾丁質框架的形成,參與貝殼的形成與修復。此外,長牡蠣殼基質蛋白的研究中也鑒定到幾丁質酶和幾丁質合成酶[28],在酸化海水處理的長牡蠣中,幾丁質合成酶表達水平被顯著抑制,從而抑制幾丁質的生物合成來抑制牡蠣鈣化殼的形成[29]。在厚殼貽貝中,幾丁質合成酶主要在內、中褶皺中高表達,貽貝可能通過內、中褶皺來調控幾丁質合成,從而影響貝殼的形成。在貝類幾丁質酶的研究中,珠母貝的幾丁質酶能參與貝殼的形成[30];在靜水椎實螺中,也發現幾丁質酶會影響殼的形成[31]。在厚殼貽貝中,不同類型的幾丁質酶基因在不同褶皺高表達,三個褶皺可能都能通過幾丁質的代謝來影響殼的形成,但具體的作用方式可能不同,還有待深入研究。最后,外套膜三褶皺GO功能富集分析的結果也表明內、中褶皺主要參與調控幾丁質的生物合成過程,而外褶皺可能主要參與調控角質層殼膜的形成。此外,在外褶皺中還富集到“含幾丁質角質層色素沉著”的GO term,結合組織切片觀察結果,殼膜溝處的外褶皺上皮細胞中能觀察到棕色色素,而此處的中褶皺上皮細胞卻未觀察到色素,推測殼膜的色素沉著可能主要由外褶皺來調控。
4 結論
根據貝殼和外套膜邊緣的形態觀察,本研究發現厚殼貽貝外套膜邊緣具有1 個較大的內褶皺及較小的中、外褶皺。在三褶皺中,外褶皺最小,其牢固貼附于殼上,外褶皺和中褶皺之間產生角質層殼膜并延伸包裹在貝殼表面,根據組織切片觀察,三個褶皺的上皮細胞形態也存在明顯不同。此外,本研究還從厚殼貽貝的角質層和貝殼中提取并鑒定到幾丁質成分,并根據三褶皺轉錄組數據鑒定到幾丁質合成酶(CHS)、幾丁質酶(CHIA)和幾丁質結合蛋白(CBP)三類幾丁質相關基因。根據功能域和表達量分析,發現幾丁質合成酶基因、β氨基己糖苷酶、二-N-乙酰殼二糖酶等幾丁質酶基因主要在內、中褶皺高表達,而與之相反的,幾丁質結合蛋白基因和殼三糖苷酶類的幾丁質酶基因主要在外褶皺中高表達。三褶皺間的GO富集分析也表明厚殼貽貝的內、中褶皺可能主要參與調控幾丁質的生物合成,而外褶皺則主要參與調控幾丁質結合、代謝和角質層色素沉著等過程。綜上所述,厚殼貽貝外套膜三褶皺不僅在形態結構上存在顯著差異,其在與幾丁質合成、代謝、結合等功能方面也存在差異,其中,外褶皺可能在厚殼貽貝的幾丁質骨架形成和色素沉著中發揮更重要的作用。本研究有助于了解厚殼貽貝貝殼有機骨架的幾丁質成分和幾丁質合成及代謝等相關基因,為厚殼貽貝貝殼形成和生物礦化的研究奠定了基礎。
