黃芪多糖對HD11雞巨噬細胞轉錄組和代謝組的影響
2024-04-02 07:12:04陳富斌徐國偉馮海鵬郭志廷韓松偉劉佳惠古雪艷張景艷李建喜HuubSavelkoul
畜牧獸醫學報 2024年3期
關鍵詞:差異
陳富斌,徐國偉,王 磊,劉 琴,馮海鵬 ,張 康,郭志廷,韓松偉,劉佳惠,古雪艷,張景艷*,李建喜*,Huub F. J. Savelkoul
(1.中國農業科學院蘭州畜牧與獸藥研究所,蘭州 730050;2.甘肅農業大學動物醫學院,蘭州 730070;3.荷蘭瓦赫寧根大學細胞生物與免疫學教研組,荷蘭瓦赫寧根市)
黃芪多糖(APS),作為豆科黃芪屬植物的主要成分之一,是免疫活性最強的一類生物大分子[1],具有免疫調節、促進免疫器官發育[2-3],增加免疫球蛋白和T淋巴細胞的數量、增強疫苗保護效力等生物學功能[4-5],已被廣泛應用于畜禽生產中[6]。天然免疫是機體重要的防御屏障,對抵抗病原微生物感染發揮至關重要的作用[7-8]。巨噬細胞是機體天然免疫調控的主要執行者,并參與特異性免疫過程[9],能消滅侵入機體的細菌、吞噬異物顆粒,消除體內衰老、損傷的細胞和細胞間質變性、殺傷腫瘤細胞,并參與免疫反應[10]。研究表明,APS對正常小鼠的細胞免疫、體液免疫及巨噬細胞功能均有明顯的增強作用[11]。然而有關其對雞巨噬細胞(HD11細胞)的調節作用機制研究尚不深入。HD11細胞是一種通過復制缺陷型禽白血病病毒MC29毒株轉化雞骨髓細胞形成的永生化細胞系,常被作為開展家禽天然免疫調控研究的模式細胞。作者研究發現,12.5~800.0 μg·mL-1APS對HD11細胞增殖活性有促進作用;100 μg·mL-1APS處理HD11細胞12 h,可能通過識別TLR2、TLR4及TLR15促進HD11細胞增殖與極化,發揮免疫增強作用[12-13]。然而相關分子機制有待于進一步的研究證實。
研究表明,巨噬細胞的糖酵解、氧化磷酸化等糖代謝途徑與其極化、免疫增強等生物學過程有緊密的相關性[14-15]。劉小琳等[16]研究發現在RAW264.7細胞中PI3K/AKT信號通路通過激活mTOR信號通路,激活糖酵解,從而激活巨噬細胞M1極化過程。轉錄組學與代謝組學是一門新興技術,該技術的應用可為疾病發生、藥理學作用等領域的深入研究提供重要依據[17-18]。通過轉錄組學和代謝組學分析發現,在β-葡聚糖(β-glucan)和卡介苗(BCG)誘導的單核巨噬細胞訓練免疫形成中糖酵解代謝過程的重編程發揮關鍵作用[19]。當糖酵解途徑被抑制后,天然免疫細胞的表觀遺傳修飾及免疫記憶均會被抑制[20]。在進一步研究發現,50 μg·mL-1APS可誘導HD11細胞形成訓練免疫,并對隨后LPS再刺激產生免疫增強的記憶特征。然而APS的這種訓練免疫作用是否與代謝重編程有相關性,尚未見文獻報道。因此,本試驗擬通過轉錄組與代謝組分析,探究50 μg·mL-1APS是否通過膜受體的識別,進而介導相關信號通路和代謝重編程發揮對HD11的免疫調節作用。
1 材料與方法
1.1 材料
試驗所用APS由中國農業科學院蘭州畜牧與獸藥研究所中獸醫實驗室制備,測得試驗 用APS總多糖含量為95.31%±3.55%;還原性多糖含量1.96%±0.03%;蛋白質含量為6.50%±0.80%;HD11細胞由西北農林科技大學動物醫學院王晶鈺老師饋贈;細胞培養液RPMI 1640、雞血清、胎牛血清和胰蛋白酶-EDTA購自Gibco公司;青鏈霉素和PBS購自索萊寶公司。
1.2 方法
1.2.1 HD11細胞復蘇 將凍存的HD11細胞在37.5 ℃下水浴解凍,1 000 r·min-1離心5 min,棄上清,取沉淀,接于1640培養基(含10%胎牛血清、2.5%雞血清和1%青鏈霉素)重懸并移入細胞培養瓶中,標記為第1代,置于41 ℃、體積分數為5%的CO2培養箱中進行培養,每隔48 h更換1次培養基,在顯微鏡下觀察細胞的形態及生長狀況。
1.2.2 HD11細胞傳代 當細胞融合度達80%~90%以上時,棄上清,用預熱的PBS洗一遍,使用胰蛋白酶-EDTA消化3 min,在鏡下可見細胞回縮呈圓形、輕拍細胞瓶可見細胞滑動時,棄去胰蛋白酶-EDTA,加1640培養基(含10%胎牛血清、2.5%雞血清和1%青鏈霉素)1 000 r·min-1離心5 min,棄上清,加入1640培養基輕輕吹打混懸細胞,使用細胞計數器計數,并調整濃度后,分裝到新的細胞培養瓶中,標記為第2代,貼壁培養24 h。傳至3~5代可進行后續試驗。
1.2.3 HD11細胞的處理與分組 將消化后的HD11細胞1 000 r·min-1離心5 min,棄上清,取沉淀,接于1640培養基(含10%胎牛血清、2.5%雞血清和1%青鏈霉素)中,調整細胞濃度為1×106·mL-1,按5 mL·瓶-1接于T25細胞培養瓶中,置于41 ℃、體積分數為5%的CO2培養箱中培養36 h,棄上清。將細胞分為空白對照組(CON組)和APS處理組(A50組)。①空白對照組細胞中加入1640培養基(含10%胎牛血清、2.5%雞血清和1%青鏈霉素)培養12 h;②50 μg·mL-1APS試驗組中用1640培養基(含10%胎牛血清、2.5%雞血清和1%青鏈霉素)將APS濃度調整為50 μg·mL-1后加入細胞中培養12 h;收集各組上清和細胞,并進行后續試驗。
1.2.4 轉錄組學、代謝組學及二者關聯分析 將培養的細胞去除培養基,用預冷的PBS清洗2~3次,棄去上清,最后加入1 mL預冷的80%的甲醇水溶液,使用細胞刮將培養容器中所有細胞都刮干凈,轉入2 mL凍存管中,置于液氮中速凍15 min后,保存于-80 ℃冰箱中;轉運過程中,將細胞樣本置于足量干冰中運輸。
1.2.5 統計學分析 轉錄組學差異基因的篩選標準是|log2(Fold change)|≥1且Padj≤0.05。代謝組學差異代謝物的篩選主要參考VIP、FC和P-value 3個參數,VIP是指PLS-DA模型第一主成分的變量投影重要度,VIP值表示代謝物對分組的貢獻;FC指差異倍數(Fold change),為每個代謝物在比較組中所有生物重復定量值的均值的比值;P-value是通過T-test計算得到,表示差異顯著性水平。采用一維方差分析,設定閾值為VIP>1.0,FC>1.5或FC<0.667的方法篩選差異代謝物。并計算出差異代謝物在兩組間的差異變化倍數 FC(fold change),通過差異變化倍數判斷差異代謝物的上調或者下調。
2 結 果
2.1 APS處理組與空白對照組的轉錄組分析
2.1.1 差異表達基因分析 轉錄組測序結果中的共表達維恩圖展示了每個組中唯一表達的基因數量,重疊區域展示了兩個組中共表達的基因數量。APS處理組(A50)與空白對照組(CONT)的唯一表達基因數量與共表達基因數如圖1所示,空白對照組總共鑒定出11 116個表達基因,而APS處理組共鑒定出11 021個表達基因,其中二者共同表達的基因數為10 592個。

圖1 共表達韋恩圖Fig.1 Co-expression Venn diagram
如圖2所示,以|log2(Fold change)|≥1且Padj≤0.05為標準,APS處理組與空白對照組相比較,共篩選到差異基因845個,其中上調基因415個,下調基因430個。

圖2 差異表達基因統計Fig.2 Statistics of differentially expressed genes
2.1.2 差異表達基因KEGG富集分析 KEGG通路數據庫包括各種生物通路中分子相互作用網絡和生物體所特有的變化形式,從而確定蛋白質參與的最重要的信號轉導通路和生物代謝通路。將APS處理組與空白對照組的差異表達基因進行KEGG通路富集分析發現,APS處理組與空白對照組的224個差異基因富集于113條KEGG通路,差異表達基因主要富集在細胞因子-細胞因子受體相互作用、Toll樣受體信號通路、吞噬體、AGE-RAGE等信號通路,其中細胞因子-細胞因子受體相互作用、Toll樣受體信號2條通路最為顯著性(圖3)。

點的大小代表注釋到GO term上的基因數,顏色從紅到紫代表富集的顯著性大小The size of the dots represents the number of genes annotated onto the GO term, and the colors from red to purple represent the significance size of enrichment
2.1.3 關鍵信號通路中差異基因的表達情況 細胞因子-細胞因子受體信號通路與Toll樣受體信號通路是兩條對HD11胞免疫調節作用影響的重要途徑,與細胞的免疫調節等密切相關。對APS處理組與空白對照組的差異基因表達進行分析,發現在細胞因子-細胞因子受體信號通路中富集了19個差異基因,其中上調基因17個,包括IL-8、CXCL13、CSF1、TNFSF8等,下調基因2個,包括IL-12B和CCR5(表1);在Toll樣受體信號通路中主要富集了11個差異基因,其中上調基因5個,包括IL-8、IL8L1、TLR1B、CCL4和TLR1A,下調基因6個,包括TLR2A、TLR2B、IL-12B、FOS、TLR3和MAP2K7(表2);在其他代謝通路富集的差異基因中ACSS2、iNOS、ATP6V0、MTA為上調基因,H6PD、PRPS2、CA、MSA為下調基因(表3)。

表1 細胞因子-細胞因子受體相互作用的基因表達情況

表3 其他代謝途徑基因的表達情況
2.2 APS處理組與空白對照組代謝組分析
2.2.1 代謝組學多元統計分析 主成分分析(principal component analysis, PCA)是將一組可能存在相關性的變量,通過正交變換轉換為一組線性不相關變量的統計方法,轉換后的這組變量即稱為主成分。依據PCA模型顯示,第一主成分可顯示30.68%的代謝物信息,第二主成分可顯示25.55%的代謝物信息,APS處理組與空白對照組代謝物的第一主成分和第二主成分在空間中的分布相對獨立,存在顯著差異(圖4)。

圖4 代謝組學樣本PCA分析圖Fig.4 PCA analysis of metabonomic samples
(圖5 Fig.5)
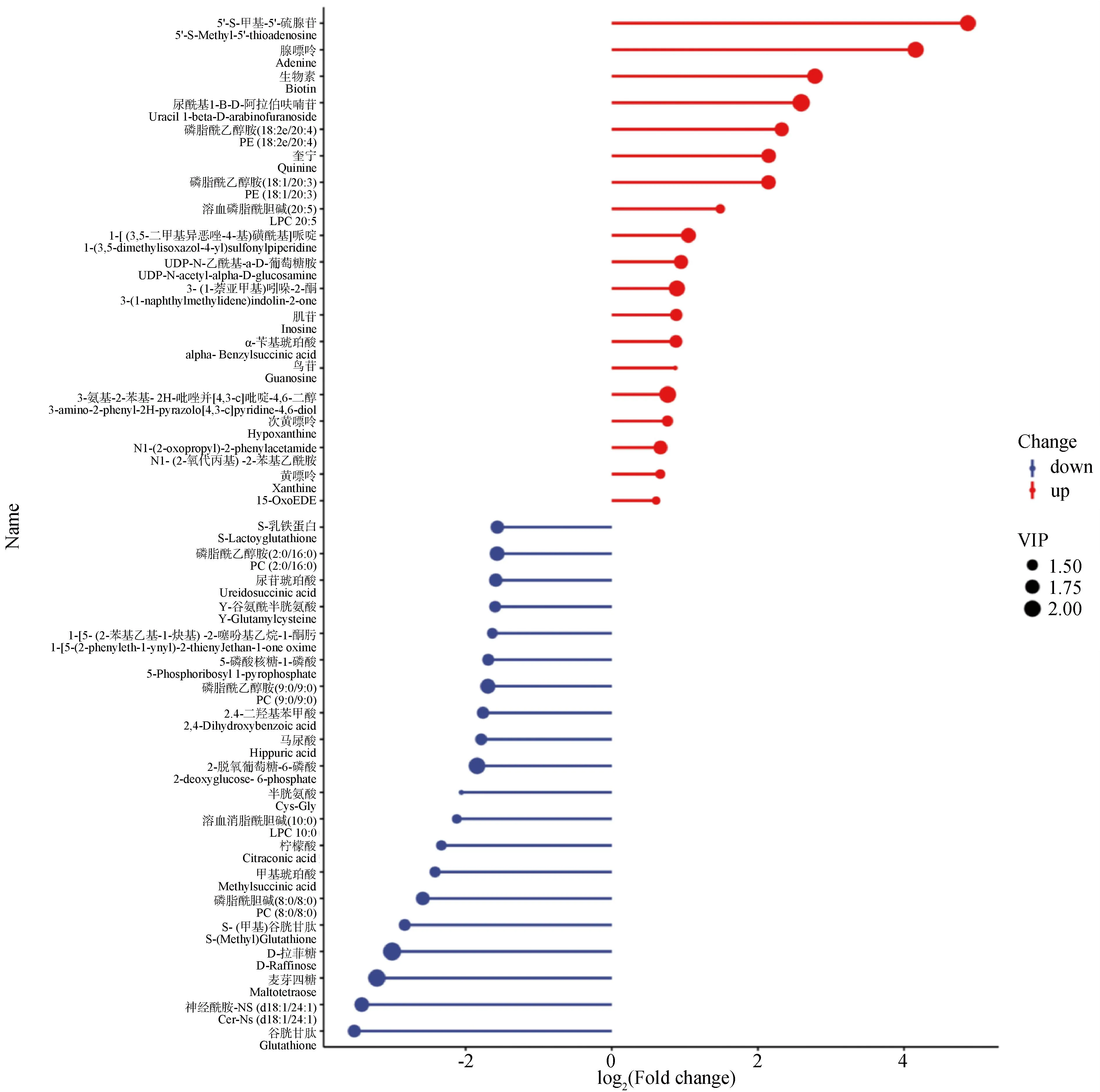
2.2.2 差異代謝物分析 利用液相色譜-質譜技術,參比空白對照組,APS處理組共篩選出623個差異代謝物,其中差異顯著的代謝物總數為89個。在差異顯著的代謝物中,下調的差異代謝物為70個,上調的差異代謝物為19個。根據組間差異比較組合得到的差異代謝物繪制火柴桿圖,能夠較為清晰的表示代謝物的上、下調以及差異倍數變化較大的物質。以log2(Fold change)進行排序,分別取前20的上調/下調代謝物進行火柴桿圖的展示。其中5′-S-甲基-5′-硫腺苷、腺嘌呤、生物素等顯著上調;谷胱甘肽、神經酰胺、麥芽四糖等顯著下調(圖5)。
2.2.3 差異代謝物KEGG通路富集分析 對APS處理組和空白對照組的差異表達代謝進行KEGG通路富集分析,主要涉及的通路包括細胞轉化、環境信息過程、遺傳信息過程、代謝和有機系統5類,包括氨基酸代謝、類脂物代謝、輔助因子和維生素的代謝、核苷酸代謝等20條通路發生變化(圖6)。

圖6 KEGG富集柱狀圖Fig.6 KEGG enrichment bar graph
2.3 APS處理組與空白對照組的轉錄組和代謝組聯合分析
2.3.1 基因分析差異結果 轉錄組與代謝組是檢測特定狀態下生物個體、組織或細胞的基因和代謝物的表達水平。APS處理組與空白對照組比較在上調中有415個顯著差異的基因,有19個顯著差異的代謝物;在下調中有430個顯著差異的基因,有70個顯著差異的代謝物。
2.3.2 代謝與轉錄KEGG富集分析 基于代謝與轉錄同一比較對的KEGG富集結果,以KEGG pathways為條目,獲取代謝、轉錄共同富集到的通路,可信度排名前十的代謝通路主要有嘌呤代謝、碳代謝、磷酸戊糖途徑、谷胱甘肽代謝、組氨酸代謝、氨基糖和核苷酸糖代謝、α-亞麻酸代謝、ABC轉運體、鐵死亡和酪氨酸代謝(圖7)。

Meta 代謝(圓點是代謝數據),Tran 轉錄(三角是轉錄數據;P value是通路富集(轉錄或代謝)的P value;value是通路中富集的代謝物或基因的個數,通路中富集的是該通路中富集到的差異代謝物或差異基因與該通路中注釋到的代謝物或基因個數的比值Meta metabolism (dots are metabolic data), Tran transcription (triangles are transcriptional data; P value is the pvalue of pathway enrichment (transcription or metabolism); Count is the number of metabolites or genes enriched in the pathway, and Ratio is the ratio of differential metabolites or genes enriched in the pathway to the number of metabolites or genes annotated in the pathway
3 討 論
APS是一種具有免疫調節活性的多糖,其對機體巨噬細胞的調節作用機制研究是近年來的研究熱點。然而有關其對雞巨噬細胞的調節作用機制尚不明確。基于前期研究結果,作者推測APS可能是通過相關信號通路和糖代謝相關途徑的調控,對HD11細胞發揮免疫增強和訓練免疫作用。因此,本研究利用轉錄組和代謝組學技術,對50 μg·mL-1處理的HD11細胞與對照組HD11細胞進行差異分析,旨在為APS的天然免疫調節機制研究提供理論和科學依據。在轉錄組結果中,APS處理組與空白對照組共篩選出差異基因845個,APS處理組415個基因表達較空白對照組上調、430個基因表達下調;APS處理組與空白對照組的224個差異基因富集于113條KEGG通路,差異表達基因主要在細胞因子-細胞因子受體相互作用、Toll樣受體信號通路、吞噬體、AGE-RAGE等信號通路上富集。
炎癥因子主要是由單核/巨噬細胞或淋巴細胞產生的細胞因子,包括IL-1或、IL-6、IL-8、TNF-或等[21-22],參與機體多種疾病的炎癥、感染和免疫等多種生理病理過程[23]。結果發現,APS處理組促進IL-8、CXCL13及CCL4的轉錄,抑制CCR5、IL-12的轉錄。Toll樣受體是一類天然免疫模式識別受體,在病原微生物免疫和誘導快速防御反應激活中具有重要作用[24]。TLRS在不同宿主中存在多態性,且由于宿主生存環境存在的差異,導致TLRS信號途徑介導的宿主免疫系統調節分子機制具有差異性[25]。在Toll樣受體信號通路中,以往植物多糖的研究主要集中在TLR4上[26-28],而本研究結果顯示,APS促進HD11雞巨噬細胞IL-8L1、TLR1B、CCL4和TLR1A的轉錄,抑制TLR2A、TLR2B、IL-12B、FOS、TLR3和MAP2K7的轉錄。作者在前期研究中也發現,與哺乳類動物不同,APS和LPS抑制TLR2和TLR4的轉錄,且APS對TLR2 mRNA更加敏感[29]。另在LPS處理12 h的HD11細胞轉錄組分析中,也發現TLR2信號通路受到主要富集,其中,TLR1和TLR2為下調差異基因;CCL5、IL-8為上調差異基因[30-31]。通過與已有試驗結果和相關文獻的比對,作者推測50 μg·mL-1APS對HD11細胞的免疫調節作用與TLR2信號通路調控相關。
近年來研究表明,糖代謝重編程是巨噬細胞極化的根本原因[32],其作為免疫刺激與表觀遺傳修飾的重要鏈接,也是訓練免疫形成的重要機制[33]。在轉錄組結果中,作者還發現ACSS2 (糖酵解關鍵酶基因)、iNOS(參與細胞傳導和免疫調節的關鍵酶基因)、ATP6V0(在細胞代謝功能和抑炎性表型都具有重要作用)、MTA(一種甲基轉移酶抑制劑)顯著性上調,H6PD(H6PDH,是磷酸戊糖途徑NAPDH合成的關鍵酶,與G6PDH具有相似的反應特點)、PRPS2(核苷酸合成途徑中一種重要的限速酶)顯著性下調。在代謝組結果中,作者發現APS處理組顯著性上調差異表達代謝物主要有5′-S-甲基-5′-硫腺苷、腺嘌呤、生物素等;顯著性下調差異表達代謝物主要有谷胱甘肽、神經酰胺、麥芽四糖等,且差異代謝物主要在5-磷酸核糖-1-焦磷酸代謝、腺嘌呤、次黃嘌呤等通路上富集。在轉錄組與代謝組聯合分析結果中,作者發現嘌呤代謝、碳代謝(包括三羧酸循環、糖酵解)、磷酸戊糖途徑、谷胱甘肽代謝等是APS處理組的主要富集代謝途徑。研究發現在免疫作用刺激下,細胞中代謝重新編程包括葡萄糖攝取增加,糖酵解速率提高,磷酸戊糖途徑上調以及通過TCA循環的氧化磷酸化水平降低[34]。因此,可以看出50 μg·mL-1APS對HD11細胞的免疫調節作用與糖酵解、磷酸戊糖的代謝重編程相關。
4 結 論
50 μg·mL-1APS對HD11細胞的免疫調節作用機制與TLR2受體信號通路、糖酵解、磷酸戊糖的代謝重編程相關,但其分子調節機制仍需在蛋白水平、關鍵代謝基因及代謝產物表達水平上進一步驗證。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
