苯丙烯菌酮對水稻稻瘟病病原真菌細胞壁膜的作用
2019-01-19 06:17:39劉雪關(guān)麗杰
江蘇農(nóng)業(yè)科學 2019年22期
關(guān)鍵詞:作用機制
劉雪 關(guān)麗杰

摘要:初步探究苯丙烯菌酮對水稻稻瘟病病菌的抑菌機制。采用紫外分光光度法測定參與水稻稻瘟病病菌細胞壁降解的2個關(guān)鍵酶(幾丁質(zhì)酶和β-1,3-葡聚糖酶)活性以及菌體細胞外的磷濃度;采用熒光分光光度法測定PI-DNA復合物(碘化吡啶)熒光強度;采用氣質(zhì)聯(lián)用法(GC-MS)測定甾醇類物質(zhì)含量變化。結(jié)果表明,用10 mg/L苯丙烯菌酮處理水稻稻瘟病病菌后,菌體內(nèi)的幾丁質(zhì)酶和β-1,3-葡聚糖酶活性均升高,藥劑處理1 h后酶活性明顯升高,分別是對照組的1.69倍和3.76倍,細胞外的磷濃度在1 h后明顯升高,3 h時是對照組的1.29倍,24 h時PI-DNA復合物熒光強度急劇升高,為對照組的19.71倍,然后用10 mg/L苯丙烯菌酮處理6 h后6-乙酰氨基麥角甾醇含量與對照組相比減少了96.83%,酵母甾醇則消失。結(jié)果表明,苯丙烯菌酮可以破壞水稻稻瘟病菌細胞壁和細胞膜功能結(jié)構(gòu),起到抑菌作用。
關(guān)鍵詞:水稻稻瘟病菌;殺真菌劑;苯丙烯菌酮;作用機制;植物病原真菌
中圖分類號: S435.111.4+1 ?文獻標志碼: A ?文章編號:1002-1302(2019)22-0117-04
水稻稻瘟病是一類重要的水稻病害,并且水稻稻瘟病病菌侵染能力強,能在短時間內(nèi)快速傳播[1-2],同時易發(fā)生菌絲融合現(xiàn)象,形成多核體,這種變異增加了水稻病害防治工作的難度[3]。全球每年因為水稻減產(chǎn)造成很大的經(jīng)濟損失,迫切需要一種新型環(huán)境友好的殺菌劑來解決水稻病害[4]。苯丙烯菌酮別稱異補骨脂查爾酮,是天然植物補骨脂提取物中的一個重要活性成分。有研究表明,二聚黃酮類化合物對人類致病酵母菌和絲狀真菌均有較好的抑菌效果[5],查爾酮類化合物也具有優(yōu)良的抗細菌真菌活性[6]。據(jù)報道,異補骨脂查爾酮具有多種藥理活性,包括抗細菌、抗真菌、抗病毒、抗炎和抗腫瘤作用[7]。Zhang等在抑菌試驗中發(fā)現(xiàn),枯草芽孢桿菌 KB-1122對稻瘟病菌生長有明顯的抑制作用,經(jīng)過二維聚丙烯酰胺凝膠電泳和蛋白組分析試驗發(fā)現(xiàn),甘油醛-3-磷酸脫氫酶和絲氨酸蛋白激酶與枯草桿菌KB-1122的抑菌作用密切相關(guān)[8]。檸檬醛可以提高幾丁質(zhì)酶活性,從而破壞水稻稻瘟病菌細胞壁完整性,達到抑菌目的[9]。硝基苯乙烯類化合物能明顯抑制稻瘟病菌的黑色素合成相關(guān)酶3HNR的活性,顯示出抑菌效果[10]。
盆栽試驗和田間試驗已經(jīng)證實了苯丙烯菌酮乳油劑對稻瘟病菌有較好的防治效果[11]。掃描電鏡結(jié)果顯示,經(jīng) 10 mg/L 苯丙烯菌酮處理后,水稻稻瘟病菌菌絲界限變得模糊,且相互交連發(fā)生融合,菌體形態(tài)發(fā)生了明顯變化。本試驗為進一步明確苯丙烯菌酮對水稻稻瘟病菌的殺菌機制,通過測定細胞壁水解酶(幾丁質(zhì)酶和β-1,3葡聚糖酶)活性[12-13]、細胞膜通透性試驗、PI熒光檢測和甾醇類物質(zhì)成分分析試驗,探究苯丙烯菌酮對稻瘟病菌菌體細胞壁和細胞膜的作用[14-15],為苯丙烯菌酮殺菌作用提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
1.1.1 主要試劑 苯丙烯菌酮(異補骨脂查爾酮),購自成都普菲德生物技術(shù)有限公司;丙酮,購自福晨(天津)化學試劑廠;葡萄糖,購自天津市大茂化學試劑廠;酵母浸粉,購自安琪酵母股份有限公司;瓊脂,購自福建省金燕海洋生物科技股份有限公司。
1.1.2 供試菌種 水稻稻瘟病菌(Pyricularia oryzae Cav.),由沈陽化工研究院生物測定中心提供。
1.1.3 培養(yǎng)條件 稻殼培養(yǎng)基(1 000 mL):稻殼30 g,葡萄糖5.0 g,酵母浸粉1.4 g,瓊脂20~25 g,MgSO4 0.25 g。
PDA液體培養(yǎng)基(1 000 mL):馬鈴薯200 g,葡萄糖20 g。
1.1.4 主要儀器 DHG-9149A型電熱恒溫鼓風干燥箱,購自上海精宏實驗設(shè)備有限公司;MPLR-702四孔恒溫水浴鍋,購自金壇市大地自動化儀器廠;SHZ-DZⅢ循環(huán)水式真空泵,購自河南省鞏義市予華儀器有限責任公司;SZ-93自動雙重純水蒸餾器,購自上海亞榮生化儀器廠;SCIENTZ-ⅡD超聲波細胞破碎儀,購自寧波新芝生物科技股份有限公司;SPX-250B-Z生化培養(yǎng)箱,購自上海博迅實業(yè)有限公司醫(yī)療設(shè)備廠;SW-CJ-IG單人凈化工作臺,購自蘇州凈化設(shè)備有限公司;TBL-16G-A高速冷凍離心機,購自上海安亭科學儀器廠;TSQ-280振蕩培養(yǎng)箱,購自上海精宏實驗設(shè)備有限公司。
1.2 試驗方法
1.2.1 苯丙烯菌酮對稻瘟病菌細胞壁的影響
1.2.1.1 菌絲的制備 取5 mL水稻稻瘟病菌孢子懸液接入PDA培養(yǎng)液中,置于28 ℃、150 r/min振蕩培養(yǎng)6 d后進行處理,苯丙烯菌酮處理濃度為10 mg/L,對照加等量丙酮,每個處理設(shè)3次重復。藥劑處理后,在時間為0、1、3、6、12、24 h收集菌絲,抽干水分后在-20 ℃下保存。
1.2.1.2 酶液的提取 稱取1 g菌絲,加入5 mL Tris-HCl(0.05 mol/L,pH值7.5)緩沖液,超聲破碎30 min(工作5 s間歇3 s)。然后將破碎的細胞勻漿轉(zhuǎn)移至EP管中,4 ℃、12 000 r/min 離心15 min,取上清液置于-80 ℃儲存?zhèn)溆谩?/p>
1.2.1.3 幾丁質(zhì)水解酶含量測定 向潔凈的試管中加入 0.2 mL 膠狀幾丁質(zhì),用0.1 mL乙酸鈉緩沖液(濃度為 0.1 mol/L,pH值4.5)和不同時間處理酶液0.3 mL,于40 ℃下水浴1 h(空白用三蒸水代替酶液),流水冷卻至室溫。12 000 r/min 離心10 min,移取上清液0.25 mL,加40 μL 10%脫鹽蝸牛酶,37 ℃保溫存1 h,冷卻。然后加0.1 mL硼酸鉀溶液(0.8 mol/L),沸水浴3 min,冷卻至室溫,加2 mL 1%二甲基胺硼烷(DMAB)溶液于37 ℃水浴20 min,冷卻,于波長585 nm處測定吸光度。根據(jù)N-乙酰葡萄糖胺標準曲線y=0.059x-0.007(r2=0.997)進行計算。以單位鮮質(zhì)量樣品在單位時間內(nèi)產(chǎn)生的N-乙酰葡萄糖胺為1個酶活力單位。
U=(D-Do)×VtFW×Vs×t。
式中:D為樣品中幾丁質(zhì)酶水解生成的N-乙酰葡萄糖胺含量;Do為空白組N-乙酰葡萄糖胺含量;Vt為酶液總體積,mL;Vs為反應酶液體積,mL;FW為樣品的鮮質(zhì)量,g;t為反應時間,h。
1.2.1.4 β-1,3-葡聚糖水解酶含量測定 β-1,3-葡聚糖水解酶含量測定的反應體系包括0.05 mol/L pH值5.0的乙酸鈉緩沖液(含1%昆布多糖)、0.16 mL 0.05 mol/L乙酸鈉(pH值5.0)緩沖液和300 μL各處理酶液(空白用三蒸水代替酶液),充分混勻,37 ℃保溫1 h,使其充分水解為葡萄糖,采用DNS法測定還原糖的量,向各反應體系中加入 3,5-二硝基水楊酸試劑沸水浴5 min,流水立即冷卻,定容至 3 mL,于540 nm波長下測D值[16]。根據(jù)葡萄糖標準曲線計算酶活性:y=0.173x-0.009,r2=0.998。以單位鮮質(zhì)量樣品在單位時間內(nèi)產(chǎn)生的還原糖為1個酶活力單位。
U=(D-Do)×VtFW×Vs×t。
式中:D為樣品中β-1,3葡聚糖水解酶水解生成的還原糖含量;Do為空白組還原糖含量;Vt為酶液總體積,mL;Vs為反應酶液體積,mL;FW為樣品的質(zhì)量,g;t為反應時間,h。
1.2.2 苯丙烯菌酮對稻瘟病菌細胞膜的影響
1.2.2.1 菌懸液的制備 取5 mL水稻稻瘟病菌孢子懸液接入PDA培養(yǎng)液中,于28 ℃、150 r/min振蕩培養(yǎng)6 d后進行處理,苯丙烯菌酮處理濃度為10 mg/L,對照加等量丙酮,每個處理設(shè)3個重復。藥劑處理后0、1、3、6、12、24 h取樣。
1.2.2.2 磷濃度標準曲線 磷濃度標準曲線的制備:準確稱取經(jīng)100 ℃干燥至恒質(zhì)量的磷酸二氫鉀43.9 g,加水溶解并定容至250 mL,精確量取10 mL于100 mL容量瓶中,稀釋至刻度,得到0.004 mg/mL的參比溶液。分別取參比試劑0、1、3、5、7、9 mL于25 mL具塞試管中,加6 mL定磷試劑[水 ∶ 3 mol/L 硫酸 ∶ 2.5%鉬酸銨 ∶ 10%維生素C為 2 ∶ 1 ∶ 1 ∶ 1],加水稀釋至25 mL,搖勻,45 ℃水浴20 min,冷卻至室溫,在660 nm波長處測定吸光度,并以磷濃度為橫坐標、吸光度為縱坐標繪制標準曲線y=0.242 8x+0.009 6,r2=0.996。
1.2.2.3 胞外磷濃度含量測定 分別在藥劑處理后0、1、3、6、12、24 h取菌懸液8 mL,3 000 r/min離心10 min,精確量取上清液5 mL于平底燒瓶中,加9 mol/L硫酸溶液5 mL,直火緩慢加熱15 min,冷卻至室溫,滴加過氧化氫5 mL,繼續(xù)加熱15 min,冷卻,轉(zhuǎn)移至100 mL容量瓶中,稀釋至刻度,精確量取5 mL,余下步驟同“1.2.2.2”節(jié)磷濃度標準曲線測定方法。根據(jù)標準曲線計算磷濃度,磷濃度單位為μg/mL。
1.2.2.4 PI-DNA熒光檢測 用滅菌后的超純水配制 50 μg/mL 的PI溶液,置于棕色瓶內(nèi),4 ℃下避光保存。取 5 mL 菌懸液于離心管中,加入1 mL PI溶液和1 mL藥劑,充分混合,在37 ℃下進行溫育反應,分別在反應0、1、3、6、12、24 h時測定其熒光值(激發(fā)波長和發(fā)射波長分別為535、615 nm)。
1.2.2.5 甾醇成分分析 取菌絲0.5 g于50 mL具塞試管中,加2.5 mL PBS和6 mL新鮮配制的皂化劑,充分混勻,80 ℃ 水浴皂化1 h。加6 mL石油醚60 ℃提取3次,收集餾分,加水洗滌。醚層60 ℃水浴揮干石油醚,得到未皂化脂,加色譜純環(huán)己烷(1 mL/g濕菌)后于-20 ℃保存[17]。
色譜條件:5% HP-5MS(Phenyl Siloxane)30.0 m×0.250 mm×0.25 μm;柱溫:初溫度100 ℃,最終溫度300 ℃,進樣口溫度250 ℃;載氣:氦氣1.0 mL/min。質(zhì)譜標準庫:NBS譜庫。
2 結(jié)果與分析
2.1 苯丙烯菌酮對水稻稻瘟病菌細胞壁的影響
2.1.1 苯丙烯菌酮對水稻稻瘟病菌幾丁質(zhì)酶活性的影響 由圖1可知,經(jīng)10 mg/L苯丙烯菌酮處理后,菌體內(nèi)幾丁質(zhì)酶活性明顯高于對照組。藥劑處理1 h,幾丁質(zhì)酶活性為對照組1.69倍,處理3、6、12、24 h后分別是對照組的1.41倍、1.54倍、1.33 倍、1.43倍。幾丁質(zhì)是真菌細胞壁的主要成分,是由N-乙酰葡糖胺通過β連接聚合而成的結(jié)構(gòu)同多糖。幾丁質(zhì)酶活性升高導致幾丁質(zhì)水解速度加快,細胞壁幾丁質(zhì)層受損,其功能受到影響,使菌體無法正常生長。
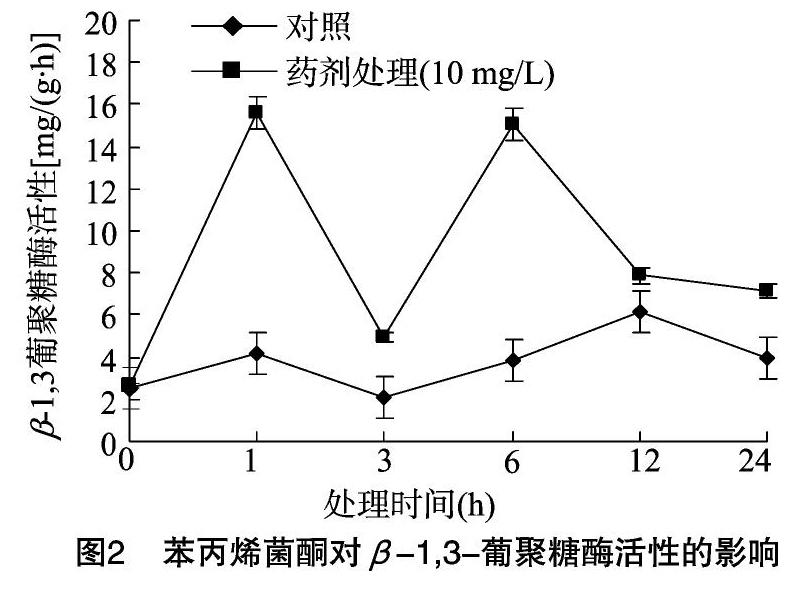
2.1.2 苯丙烯菌酮對水稻稻瘟病菌β-1,3-葡聚糖酶活性的影響 由圖2可知,藥劑處理組菌體內(nèi)β-1,3-葡聚糖酶活性明顯高于對照,在藥劑處理后1、3、6、12、24 h后分別是對照組的3.76倍、2.33倍、3.91倍、1.29倍、1.81倍。葡聚糖是以葡萄糖為單糖組成的同型多糖,葡萄糖單元之間以糖苷鍵連接,是真菌細胞壁的重要組成部分。經(jīng)苯丙烯菌酮處理后,菌體內(nèi)β-1,3葡聚糖酶活性升高,葡聚糖發(fā)生水解,引起稻瘟病菌細胞壁降解,所以藥劑通過破壞真菌細胞壁達到抑菌目的。
2.2 苯丙烯菌酮對稻瘟病菌細胞膜的影響
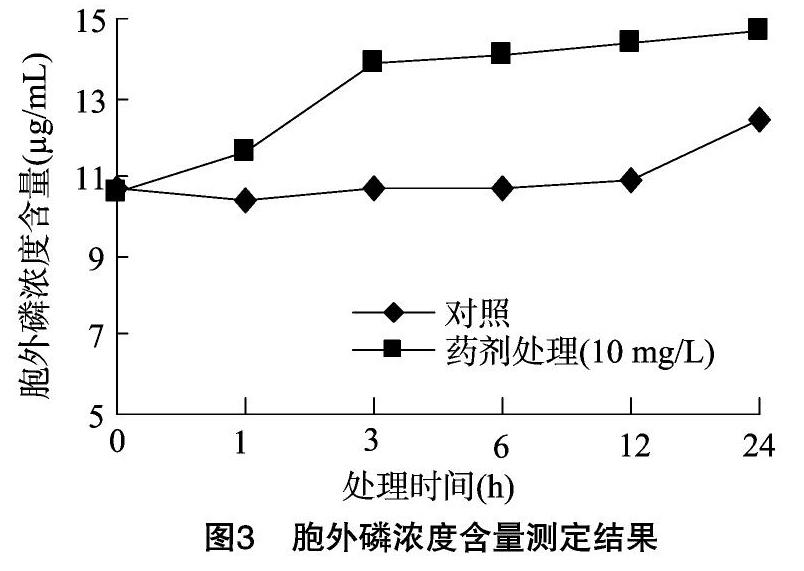
2.2.1 苯丙烯菌酮對稻瘟病菌細胞膜通透性的影響 磷是真菌細胞內(nèi)的重要營養(yǎng)物質(zhì),主要以多磷酸鹽形式存在,正常細胞胞外磷濃度遠遠小于胞內(nèi)磷濃度,但當細胞膜受到損傷時,胞液滲漏可使胞外磷濃度升高,分別在藥劑處理菌懸液0、1、3、6、12、24 h后取菌懸液測定胞外磷濃度,結(jié)果如圖3所示。
由圖3可知,在藥劑處理1 h后胞外磷濃度明顯升高,藥劑處理1、3、6、12、24 h后胞外磷濃度分別是對照組的1.12倍、1.29倍、1.31倍、1.31倍、1.17倍。由此可知,殺菌劑苯丙烯菌酮處理菌體細胞后,真菌細胞膜發(fā)生一定程度的破損,導致營養(yǎng)物質(zhì)泄露。
2.2.2 PI-DNA熒光檢測結(jié)果 PI是一種可對DNA染色的細胞核染色劑,它是一種溴化吡啶的類似物,在嵌入雙鏈DNA后釋放紅色熒光。PI不能穿透完整細胞膜,只有當細胞處于凋亡晚期或者死亡時,染色劑才可以進入細胞并與核酸物質(zhì)結(jié)合形成PI-DNA復合物,結(jié)果如圖4所示。
由圖4可以看出,在藥劑處理菌懸液1 h內(nèi),復合物熒光值沒有明顯變化,處理1 h后熒光值緩慢上升,處理3、6、12 h時分別是對照組的6.35倍、3.24倍、8.30倍,24 h時復合物熒光值急劇升高為對照組的19.71倍。由此可以推測,苯丙烯菌酮處理稻瘟病菌后,菌體胞膜嚴重受損,染色劑進入細胞內(nèi)1 h后,菌體開始慢慢凋亡,24 h時細胞達到凋亡晚期,細胞膜完全破裂,細胞最終死亡。
2.2.3 苯丙烯菌酮對甾醇類物質(zhì)含量的影響 由圖5至圖7可知,10 mg/L苯丙烯菌酮處理后菌體酵母甾醇消失,6-乙酰氨基麥角甾醇成分減少。表1顯示,苯丙烯菌酮處理組與空白組、對照組相比分別減少了96.83%、97.07%。麥角甾醇是真菌細胞膜的重要成分,與細胞膜流動性和穩(wěn)定性密切相關(guān)。麥角甾醇含量降低,細胞膜會形成小孔,而酵母甾醇是麥角甾醇的前體物質(zhì)。由此可見,苯丙烯菌酮可能作用于麥角甾醇合成途徑,破壞稻瘟病菌細胞膜結(jié)構(gòu),起到抑菌作用。
3 結(jié)論
通過對真菌細胞壁水解相關(guān)酶活性的測定,結(jié)果表明,用10 mg/L苯丙烯菌酮處理稻瘟病菌后幾丁質(zhì)酶和β-1,3葡聚糖酶,其活性均升高,藥劑處理1 h,幾丁質(zhì)酶活性為對照組1.69倍;β-1,3葡聚糖酶在藥劑處理1、3、6、12、 24 h后分別是對照組的3.76倍、2.33倍、3.91倍、1.29倍、1.81倍。幾丁質(zhì)和葡聚糖都是真菌細胞壁的重要組成部分,其水解酶活性升高會加快細胞壁降解,導致細胞壁結(jié)構(gòu)受損,進而影響其功能。稻瘟病菌胞外磷濃度在藥劑處理1 h后含量增加,是對照組的1.12倍;PI-DNA復合物熒光強度在藥劑處理24 h時顯著升高為對照組的19.71倍;膜通透性試驗和PI熒光檢測試驗表明,苯丙烯菌酮可以破壞菌體細胞膜功能,導致營養(yǎng)物質(zhì)泄露,并且細胞膜完整性遭到破壞,菌體最終死亡。甾醇成分分析結(jié)果顯示,細胞膜上麥角甾醇合成途徑受阻,細胞膜形成小洞,流動性和穩(wěn)定性下降。綜上,苯丙烯菌酮可以破壞稻瘟病菌細胞壁和細胞膜結(jié)構(gòu)功能及其完整性,從而達到抑菌目的。
參考文獻:
[1]戴余有,邢 海,裘珺琳,等. 吡唑醚菌酯·稻瘟靈防治水稻稻瘟病應用研究[J]. 綠色科技,2017(23):154-156.
[2]周志偉. 水稻稻瘟病發(fā)生情況及綜合防治現(xiàn)狀[J]. 農(nóng)民致富之友,2016(5):53.
[3]李 楊,王耀雯,王育榮,等. 水稻稻瘟病菌研究進展[J]. 廣西農(nóng)業(yè)科學,2010,41(8):789-792.
[4]Lee W,Lee D G. An antifungal mechanism of curcumin lies in membrane-targeted action within Candida albicans[J]. IUBMB Life,2014,66(11):780-785.
[5]Wang S Y,Sun Z L,Liu T,et al. Flavonoids from Sophora moorcroftiana and their synergistic antibacterial effects on MRSA[J]. Phytotherapy Research,2014,28(7):1071-1076.
[6]Kulkarni R R,Tupe S G,Gample S P,et al. Antifungal dimeric chalcone derivative kamalachalcone E from Mallotus philippinensis[J]. Natural Product Research,2014,28(4):245-250.
[7]Kuete V,Sandjo L P. Isobavachalcone:an overview[J]. Chinese Journal of Integrative Medicine,2012,18(7):543-547.
[8]Zhang C X,Zhang X X,Shen S H. Proteome analysis for antifungal effects of Bacillus subtilis KB-1122 on Magnaporthe grisea P131[J]. World Journal of Microbiology and Biotechnology,2014,30(6):1763-1774.
[9]Li R Y,Wu X M,Yin X H,et al. Naturally produced citral can significantly inhibit normal physiology and induce cytotoxicity on Magnaporthe grisea[J]. Pesticide Biochemistry Physiology,2015,118:19-25.
[10]Chen H,Han X,Qin N,et al. Synthesis and biological evaluation of novel inhibitors against 1,3,8-trihydroxynaphthalene reductase from Magnaporthe grisea[J]. Bioorganic & Medicinal Chemistry,2016,24(6):1225-1230.
[11]關(guān)麗杰,姜翠杰,趙禮慧,等. 0.1%異補骨脂查耳酮乳油防治水稻稻瘟病藥效試驗[J]. 農(nóng)藥,2012,51(3):228-230.
[12]Dalonso N,Goldman G H,Miranda Gern R M. β-(1→3),(1→?6)-Glucans:medicinal activities,characterization,biosynthesis and new horizons[J]. Applied Microbiology and Biotechnology,2015,99(19):7893-7906.
[13]Walker S S,Xu Y,Triantafyllou I,et al. Discovery of a novel class of orally active antifungal β-1,3-D-glucan synthase inhibitors[J]. Antimicrobial Agents and Chemotherapy,2011,55(11):5099-5106.
[14]Leconte O,Bonfils J P,Sauvaire Y. Protetive of iridals from saponin injury in Candida albicans cells[J]. Phytochemistry,1997,44(4):575-579.
[15]于 洋,孔繁翔,王美林,等. 應用流式細胞技術(shù)研究銅對藻細胞膜完整性及脂酶活性的影響[J]. 應用與環(huán)境生物學報,2006,12(5):706-709.
[16]陳見暉,周 衛(wèi). 鈣對蘋果果實過氧化物酶、β-1,3-葡聚糖合成酶和β-1,3-葡聚糖分解酶活性的影響[J]. 中國農(nóng)業(yè)科學,2004,37(3):400-405.
[17]張軍東. 抗真菌天然產(chǎn)物的篩選與分子機制研究[D]. 上海:第二軍醫(yī)大學,2006.
猜你喜歡
海峽科技與產(chǎn)業(yè)(2016年12期)2017-01-16 19:06:39
中國信息技術(shù)教育(2016年24期)2017-01-03 22:35:16
商業(yè)經(jīng)濟研究(2016年22期)2016-12-27 10:51:08
中國實用醫(yī)藥(2016年29期)2016-12-26 08:53:55
農(nóng)家科技下旬刊(2016年9期)2016-12-15 08:33:09
中國中藥雜志(2016年20期)2016-11-19 12:24:52
人間(2016年26期)2016-11-03 17:51:57
中國實用醫(yī)藥(2016年24期)2016-10-17 04:45:48
科技視界(2016年17期)2016-07-15 18:29:24
商(2016年6期)2016-04-20 09:17:58
