腦紅蛋白在脊髓損傷后星型膠質細胞內的表達及意義
2019-03-08 00:44:32明超曾云熊敏
山東醫藥 2019年4期
明超,曾云,熊敏
(湖北醫藥學院附屬東風總醫院,湖北十堰 442000)
脊髓損傷(SCI)分為原發性和繼發性,原發性SCI以神經細胞壞死和神經纖維斷裂、血-脊髓屏障破壞、血管損傷等表現為主[1];繼發性SCI以細胞凋亡和進展性軸突丟失為主,主要包括微循環缺血缺氧、氧化應激損傷、興奮性氨基酸中毒、炎癥反應等[2]。研究表明氧化應激損傷在繼發性SCI的發生發展中發揮重要作用[3]。氧自由基(ROS)的堆積會造成脊髓組織氧化應激損傷,主要包括膜脂質過氧化反應、核酸和蛋白質損傷、氧化呼吸鏈酶系統抑制、神經纖維退變及脫髓鞘等,最終導致脊髓功能紊亂和損傷進一步加重[4,5]。星形膠質細胞(AS)是中樞神經系統(CNS)內分布最廣、數量最多的一類細胞。研究表明AS可通過分泌神經營養因子、減輕氧化應激損傷及缺血缺氧預適應等機制保護神經元[6]。SCI后星形膠質瘢痕可減輕神經炎癥反應,抑制軸突及神經纖維脫髓鞘而保護神經細胞免受二次損傷[7]。Anderson等[8]提出膠質瘢痕有助于SCI后軸突再生。研究認為,SCI后AS的活化是保護脊髓組織功能及修復的潛在靶點[9]。Ngb是一種定位于細胞質的攜氧球蛋白,主要分布于神經系統、視網膜細胞及內分泌腺組織中[10]。研究發現Ngb具有清除氧化應激毒性物質、調控凋亡通路、激活細胞生存信號等功能[11]。在腦損傷、腦瘧疾及自身免疫性腦炎中,Ngb除了表達于神經元,亦能表達于損傷區域的反應性AS中;且與星形膠質瘢痕形成有關[12,13]。Ngb在SCI后對神經元的保護作用已有諸多報道,而在AS中的作用卻未見確切報道。2017年1月~2018年2月,我們擬從動物模型上探討SCI后AS中Ngb的表達變化,在細胞模型中研究Ngb在保護AS免受氧化應激損傷中的作用。
1 材料與方法
1.1 材料 雌性SD大鼠30只(3月齡,體質量250~300 g)、SD新生乳鼠10只由湖北醫藥學院實驗動物中心提供。主要試劑:H2O2(美國Sigma公司);兔抗大鼠GFAP及小鼠抗大鼠Ngb抗體(英國Abcam公司);驢抗兔Cy3及驢抗小鼠FITC(武漢Antgene公司);胎牛血清FBS(以色列BI公司);DMEM高糖培養基(美國Thermo公司);TRIzol試劑(美國MRC公司);PCR引物(上海生工生物有限公司設計合成);逆轉錄試劑盒(美國Fermentas公司)。主要儀器:IX71 熒光顯微鏡(日本Olympus公司);CO2細胞培養箱(美國Thermo公司);FACS流式細胞儀(美國BD公司);全波段酶標儀(美國Biotek公司);熒光定量PCR儀(澳大利亞RotorGene公司)。
1.2 SCI模型制作 將30只SD大鼠隨機分成2組,假手術組(5只)及SCI組(25只)。SCI組采用改良Allen′s 法建立脊髓損傷模型[14],10%水合氯醛(350 mg/kg)麻醉大鼠,俯臥固定脫毛,以浮肋與第13胸椎連接處作為定位標志,后正中線縱行3 cm切口,依次暴露椎旁軟組織,咬除棘突、椎板,顯露 T10脊髓。脊髓沖擊棒(10 g)于25 mm處垂直自由下落撞擊T10硬脊膜,持續壓迫約10 s,造成中度脊髓挫傷,分層縫合后分籠喂養。SCI成功標準:脊髓組織出血,硬脊膜膨隆呈紫紅色,鼠尾痙攣性擺動,雙下肢及軀體回縮樣撲動,呈遲緩性癱瘓。本實驗均獲湖北醫藥學院動物研究倫理委員會批準。
1.3 脊髓組織取材及HE染色 術后第1、3、7、14、28天分別于SCI組隨機選取5只大鼠及假手術組選取3只大鼠,麻醉后暴露心臟,經升主動脈插管灌注生理鹽水250 mL,待右心耳流出無色清亮液體后繼以4% 多聚甲醛溶液灌注,隨后打開椎管,分離出損傷處脊髓,4%多聚甲醛固定24 h后,石蠟包埋切片(4 μm)行HE染色。在SCI后1 h取出脊髓組織,可見其連續性中斷、表面嚴重變性壞死,損傷周圍亦有不同程度的變性壞死,假手術組大鼠脊髓完整,未見明顯出血壞死。
1.4 脊髓星型膠質細胞取材及鑒定 取1~2 d的SD新生乳鼠,經75%乙醇浸泡10 min后處死,超凈臺中取出脊髓,剝離脊膜及血管,置于冰PBS溶液中剪碎,0.25%胰酶溶液室溫消化15 min,12%FBS-DMEM終止消化制成細胞懸液。200目濾網過濾、離心、重懸后接種至T25培養瓶。以差速貼壁法去除成纖維細胞。24 h后換液,以后每隔2~3 d換液1次。取6~8 d培養至90%融合的細胞于37 ℃恒溫搖床中160 r/min水平振蕩,16~18 h去除少突膠質細胞及小膠質細胞。待細胞傳至第三代時(約21 d),取少量細胞以GFAP及DAPI雙染細胞免疫熒光法鑒定AS及其純度。在染色視野范圍內,可見GFAP陽性細胞胞體突起之間排列緊密,GFAP和DAPI共表達的細胞占有核細胞總數的90%以上,故而選取第三代AS用于后續實驗。
1.5 GFAP及Ngb免疫熒光染色 第三代AS以1×105/mL密度接種于明膠包被的24孔板中,待細胞融合至80%左右時預冷PBS漂洗3次,4%多聚甲醛4 ℃固定30 min,PBS漂洗5 min×3次,后續步驟同脊髓組織免疫熒光染色。免疫熒光染色觀察AS內GFAP和Ngb表達。
1.6 AS細胞活力檢測 采用MTT法。第三代AS以5×104/mL密度接種于96孔板中,每孔200 μL,置于CO2培養箱中培養72 h。待細胞融合至80%后,分別加入不同濃度(0、50、100、150、200、400 μmol/L)H2O2處理不同時間(6、12 h)用于MTT實驗。全波段酶標儀測量波長490 nm時各孔吸光度(A)值。細胞活力=(藥物組A值-調零孔A值)/(對照孔A值-調零孔A值)×100%。
1.7 AS內源性ROS生成測定 采用流式細胞術。第三代AS按1.25×105/mL密度接種于6孔板中,每孔2 mL,細胞培養箱中培養72 h后,不同濃度(50、100、200、400 μmol/L)H2O2處理不同時間(6、12 h),胰酶消化、重懸后加入終濃度為10 μmol/L的DCFH-DA探針,37 ℃孵育1 h。無血清培養基洗滌、離心后,2%FBS-DMEM重懸細胞,流式細胞儀檢測。
1.8 Ngb、SOD2、CAT mRNA表達檢測 采用實時熒光定量PCR法。取不同濃度(50、100、200、400 μmol/L)H2O2處理不同時間(6、12 h)后的AS,TRIzol法抽提總mRNA,應用逆轉錄試劑盒將1 μg總RNA逆轉錄成cDNA,稀釋至5 ng/μL,隨后加入SYBR Green PCR Master Mix及相應引物進行熒光定量PCR反應。引物序列分別為:Ngb正向引物5′-GGCCATCCAAGAGAGGTGAT -3′,Ngb反向引物5′-CCATGCCTCCTCACTACCAA-3′;SOD2正向引物5′- CTGCTGGGGATTGATGTGTG-3′,SOD2反向引物5′-CTACAAAACACCCACCACGGCAT-3′;CAT正向引物5′-GCGGATTCCTGAGAGAGTGG-3′,CAT反向引物5′- GAATCGGACGGCAATAGGAG-3′;反應條件:95 ℃ 2 min,95 ℃ 5 s,60 ℃ 30 s,72 ℃ 30 s,72 ℃ 5 min共35個循環。以2ΔΔCt法計算目的基因相對表達量。
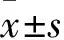
2 結果
2.1 SCI后AS內GFAP及Ngb表達 假手術組脊髓組織內少量Ngb表達,而GFAP陽性細胞呈散在分布,白質內AS胞體小,胞突細長且直,分支較少。在脊髓邊緣的白質內可見極少量GFAP和Ngb共表達于細胞質中,且顯色很弱。SCI組術后1 d,可見脊髓組織結構破壞嚴重,損傷區域周圍Ngb陽性細胞數目開始增加,與假手術組相比,GFAP和Ngb共表達稍有增加。術后3 d,Ngb表達繼續增加,損傷區域周圍GFAP和Ngb共表達明顯增加;術后1周,GFAP表達開始顯著增加,并聚集于損傷區域邊緣。與之相似,GFAP和Ngb共表達亦顯著增加,明顯高于臨近未損傷區域;術后14 d及28 d內,GFAP的增長速度最快,28 d時達到峰值,此時AS胞體突起增多、延長,突起之間形成致密網絡,形成囊腔與空洞的邊界,而周圍區域GFAP和Ngb共表達卻在14 d時達到峰值,28 d時共表達減弱。
2.2 H2O2不同濃度及時間處理AS細胞活力變化 見表1。H2O2處理6 h或12 h后,隨著H2O2濃度的增加,AS活力逐漸降低(F6 h=137.36,P<0.01;F12 h=143.61,P<0.001)。H2O2處理6 h后,100與150 μmol/L間AS活力無統計學差異(P=0.158);與之相似,12 h后150與200 μmol/L間亦無統計學差異(P=0.168)。因此確立H2O2濃度為50、100、200、400 μmol/L用于后續實驗。隨暴露時間延長,同一濃度下AS活力持續降低,而50 μmol/L H2O2時卻無統計學差異。

表1 H2O2不同濃度及時間處理AS活力變化
注:6 h時與H2O2未處理細胞相比,*P<0.01;12 h時與H2O2未處理細胞相比,#P<0.001。
2.3 H2O2處理6、12 h誘導ROS生成情況 見圖1及圖2。H2O2可誘導AS產生ROS,且在6 h內,隨著H2O2濃度的增加,ROS生成逐漸增加(峰值右偏),差異有統計學意義(F=262.33,P<0.01)。暴露于H2O212 h的AS,ROS生成先升后降(F=297.30,P<0.01),并在100 μmol/L時達到峰值。

圖1 H2O2處理6 h誘導ROS生成情況

圖2 H2O2處理12 h誘導ROS生成情況
2.4 不同時點H2O2誘導Ngb、SOD2、CAT mRNA表達變化 見圖3A、B、C。暴露于50~400 μmol/L H2O26 h或12 h后,Ngb mRNA表達呈先升后降趨勢(F6 h=67.48,P<0.01;F12 h=51.47,P<0.01)。
且在100 μmol/L 6 h或12 h時均達到峰值(t=4.72,P=0.042)。與之相似,CAT、SOD2mRNA表達亦呈先升后降趨勢(CAT:F6 h=95.31,P<0.001;F12 h=101.20,P<0.001,SOD2:F6 h=94.73,P<0.001;F12 h=41.54,P<0.01),而暴露于100 μmol/L H2O212 h時,SOD2表達量高于6 h(t=6.78,P=0.032)。Ngb、CAT、SOD2mRNA表達在50 μmol/L或100 μmol/L時達到峰值,隨后持續降低,400 μmol/L時達到最低值,呈現劑量-時間依賴效應(P均<0.05)。

圖3 H2O2處理AS抗氧化應激基因表達
3 討論
氧化應激損傷是促氧化劑和抗氧化劑平衡失調、自由基產生過多、機體或組織傾向于氧化,導致中性粒細胞浸潤、蛋白酶分泌增加,產生大量氧化中間產物的一種狀態。Bermudez等[15]研究發現,氧化應激在SCI的急、慢性期均持續存在。Amri等[19]研究發現,100 μmol/L的H2O2可使大腦皮層AS發生皺縮、膠質網絡發生塌陷、細胞密度減少;然而加入Ngb后可阻止這些改變的發生,同時Ngb還可通過減少ROS形成及促炎因子的表達而發揮抗凋亡作用[16]。
哺乳動物CNS的再生能力貧乏,嚙齒類動物(完全橫斷性脊髓損傷除外)在SCI急性期過后脊髓功能卻發生一定程度的自然恢復。就CNS損傷后的細胞反應而言,人類進化選擇似乎更傾向將小傷害隔離同時預防損傷區域感染,而不是去修復大且無法復原的損傷,AS的行為正好彰顯了這一特點。SCI后相比于AS而言,神經元細胞更易受到ROS攻擊,AS具有保護神經元免受氧化應激損傷的作用。一方面與AS含有更多的谷胱甘肽及維生素E等抗氧化物質有關,另一方面亦與細胞進化的自然選擇有關。
研究表明SCI能觸發AS增殖與活化,而活化的AS可抑制SCI區域的擴大、清除細胞碎片、重塑損傷區域[17]。Nathan等[18]研究結果顯示,脊髓環境可改變AS的命運。Okada等[7]研究證實在SCI恢復過程中,敲除AS內Socs3活化抑制基因后,AS遷移和隔離損傷區域及限制炎癥播散的能力均增強。本研究證實,在SCI的自然恢復過程中,SCI可促進AS中Ngb表達。SCI早期Ngb表達增加可能與其增加損傷區域的氧供有關。在SCI后7 d,損傷區域周圍GFAP和Ngb共表達開始明顯增加,14 d達到峰值,此時膠質瘢痕形成的速度最快,Ngb的這種變化趨勢與AS功能相匹配,由此推斷Ngb可能促進了AS的活化,從而促進SCI的自然恢復。而SCI后1~28 d Ngb呈現先升后降的表達趨勢,說明除了AS的活化,Ngb還可能扮演抗氧化應激損傷的作用,此與Amri等[16]及Fabrizius等[19]研究結果一致。
ROS是反映氧化應激的指標。本實驗結果顯示,不同濃度H2O2處理脊髓AS 6 或12 h后,細胞出現不同程度的氧化應激損傷,AS活力逐漸降低,呈現劑量-時間依賴效應。實時熒光定量PCR結果顯示,H2O2誘導AS氧化應激損傷后可促進SOD2、CAT、Ngb基因表達;且隨著H2O2濃度增加及暴露時間延長,內源性抗氧化應激酶SOD2及CAT mRNA水平呈現先升后降趨勢。與此同時,Ngb mRNA亦出現類似于SOD2、CAT基因的表達趨勢,且在100 μmol/L時達到峰值。說明Ngb可能發揮類似作用,與SOD2、CAT共同參與AS的抗氧化應激損傷。Douiri等[20]將外源性Ngb加入H2O2誘導損傷后的AS中發現,Ngb可明顯增加SOD2及CAT mRNA表達,從而發揮抗氧化應激損傷作用;而抑制AS內源性抗氧化酶系統的活性則可明顯削弱內源性保護分子促進細胞存活的能力。
本研究結果證實SCI可促進AS表達Ngb,并且Ngb可能參與了SCI后AS的抗氧化應激損傷作用,從而促進SCI的自然恢復過程。此為星型膠質細胞與腦紅蛋白抗氧化應激損傷治療脊髓損傷奠定了理論基礎。
猜你喜歡
今日農業(2021年9期)2021-11-26 07:41:24
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
電測與儀表(2015年5期)2015-04-09 11:30:52
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
