新疆7地區馕用酸面團微生物多樣性分析
2023-02-14 00:57:24于靜許倩李芬牛希躍
食品研究與開發 2023年2期
關鍵詞:酵母菌
于靜,許倩,李芬,牛希躍*
(1.阿克蘇地區疾病預防控制中心,新疆 阿克蘇 843000;2.南疆特色農產品深加工兵團重點實驗室,新疆 阿拉爾 843300;3.塔里木大學 食品科學與工程學院,新疆 阿拉爾 843300;4.塔里木大學 分析測試中心,新疆 阿拉爾 843300)
馕在新疆已有兩千多年的歷史,又稱之為“胡餅”、“爐餅”,是新疆維吾爾等少數民族的傳統主食之一[1-2],其在生活中扮演著重要角色,在新疆有“可以一日無菜,但絕不可以一日無馕”的說法[3-5]。近年來,隨著活性干酵母被引入我國且被廣泛地應用于面制品中,傳統酸面團逐漸呈現劣勢[6]。但新疆馕中的酵頭仍被廣泛應用于自治區內各大、中、小城市(鎮),主要是由于傳統酸面團發酵的馕口感更優,而活性干酵母與傳統酸面團相比則口感、風味、香氣單一[7-8]。傳統酸面團中主要是以酵母菌、乳酸菌為主的多菌種混雜體系[9],由于各種微生物代謝出不同的物質且相互之間能發生反應,從而來影響面團的風味、口感和感官品質。酵母菌在發酵面團中產生的CO2被面團中的面筋包裹而使面團體積增大,這也是面團疏松多孔的原因;乳酸菌能代謝產生乳酸等多種揮發性酸而使面制品擁有多種風味,并且可以降低面團的pH值,同時產生胞外多糖,能改變食品的流變性。乳酸菌能生成一些抑菌物質如細菌素來抑制面食中致病菌和腐敗菌的繁殖,以此來延長貨架期[10-12]。這些都是活性干酵母所不具備的,因此傳統發酵劑仍然存在于全國各地,其制作的面食仍受廣大消費者喜愛。但傳統方式制作面制品存在制作工藝粗放、技術落后、培養條件不穩定、貯藏過程變質明顯等弊端,主要是因為微生物菌群不同、衛生條件不達標、面團中含有有害菌等[13-15]。
目前只有少部分人研究馕中的微生物,并且是研究馕中的酵母菌,而對馕中微生物的多樣性研究相對較少。本文對新疆7個地區馕面團微生物多樣性進行研究,不僅為酸面團中微生物的多樣性研究提供初步的理論依據,也為后續研究區域性面食提供參考。
1 材料與方法
1.1 材料與試劑
1.1.1 樣品來源
酸面團樣品從新疆采集,共計12份,7個采集地區見表1。
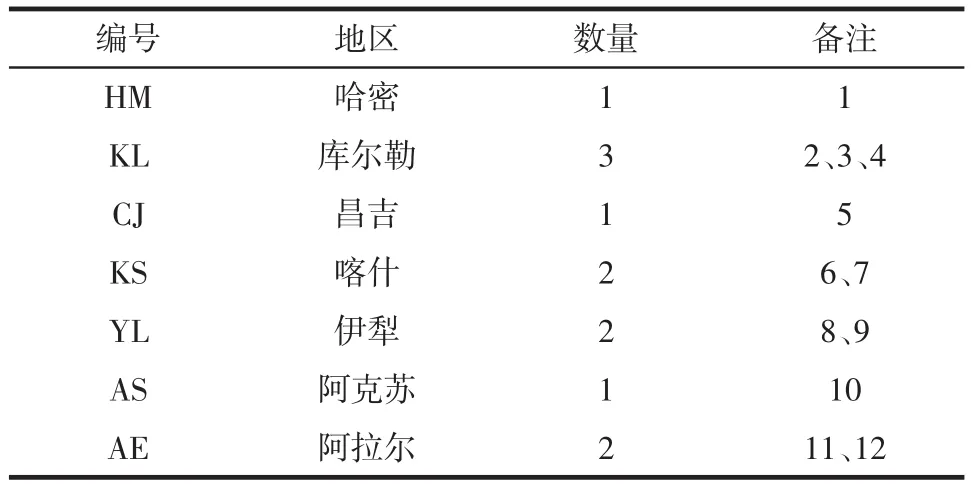
表1 各地區采集的酸面團樣品Table 1 The sourdough samples collected from different areas
1.1.2 培養基及主要試劑
酵母膏胨葡萄糖瓊脂(yeast extract peptone glucose agar medium,YPD)培養基:固體加2%瓊脂、2%蛋白胨、2%葡萄糖、1%酵母膏;WL營養瓊脂培養基:青島日水生物有限公司;MRS培養基:北京奧博星生物技術有限責任公司;革蘭氏染色液:珠海貝索生物技術有限公司;引物27f/1492r、NL1/NL4:上海生工生物工程股份有限公司。
1.2 主要儀器
PHS-3C型pH計:上海儀電科學儀器股份有限公司;TGL-20bR型高速冷凍離心機:上海安亭科學儀器廠;HJ-4A型數顯恒溫磁力攪拌器:江蘇金怡儀器科技有限公司;HPX-9162MBE型電熱恒溫培養箱:上海博迅醫療生物儀器股份有限公司;PowerPac Universal型電泳儀、MYCYCLER型PCR儀及ChemiDoc XRS+型凝膠成像系統(化學發光):伯樂生命醫學產品(上海)有限公司;CX21FS1型生物顯微鏡:日本奧林巴斯光學工業株式會社。
1.3 方法
1.3.1 樣品處理
在無菌室中稱取25 g樣品于225 mL滅菌生理鹽水(0.85%)中,用預先滅菌的磁力攪拌器充分攪拌混勻(干樣品則預先用無菌生理鹽水浸泡),采用10倍梯度稀釋法稀釋至10-7,備用。
1.3.2 乳酸菌的分離純化
樣品稀釋方法同1.3.1。在無菌環境中,取稀釋梯度為10-2~10-4,用滅菌的槍頭從每個梯度各吸取100 μL均勻涂布于MRS固體培養基中,在37℃厭氧條件下48 h~72 h培養。在同一個樣品不同梯度下挑選出不一樣的疑似乳酸菌的菌株接種到液體培養基中,37℃培養24 h~48 h,再進行平板劃線純化,經過數次反復的劃線純化,革蘭氏染色、鏡檢無雜菌為止,再進行過氧化氫酶試驗。將革蘭氏染色陽性、過氧化氫酶陰性菌初步視為乳酸菌;其余則根據鏡檢后的形態及顏色另行歸類[16]。
1.3.3 酵母菌的分離純化
選擇稀釋梯度為10-2~10-4的樣液,吸取100 μL均勻涂布于YPD固體培養基中,在28℃下培養48h~72h,在同一個樣品不同梯度下挑選出不一樣的菌株接種到液體培養基中,在相同溫度條件下培養24 h~48 h再進行平板劃線純化,反復多次劃線后經鏡檢無雜菌為止。
1.3.4 酵母菌形態學的初步認定
將1.3.3中獲得的純酵母菌劃線接種到WL營養瓊脂培養基中,28℃培養5 d~7 d,根據平板上菌落的顏色與形態,來判定酵母菌的類別。
1.3.5 DNA提取與PCR擴增
細菌用十二烷基硫酸鈉法,真菌采用凍融法[17]分別提取相應菌株的DNA,并進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增。細菌進行16S rDNA擴增,采用細菌通用引物27f和1492r,PCR反應體系(50 μL)如下:10buffer5.0μL,dNTPs2.0μL,正、反引物各 2.0μL,ExTaq 酶 0.5 μL, 模板 DNA2.0μL,dwater 36.5 μL。 PCR 反應條件:94℃預變性5 min;94℃變性30 s,56℃退火 30 s,72℃延伸 2 min,30個循環;72℃終延伸10 min,4℃保存。真菌進行26S rDNA擴增,采用酵母菌擴增引物NL1和NL4,PCR反應體系(50 μL)如下:10buffer 5.0 μL,dNTPs 4.0 μL,正、反引物各 3.0 μL,ExTaq 酶 0.5 μL,模板 DNA 2.0 μL,dwater 32.5μL。其PCR反應條件:94℃預變性5 min;94℃變性 30 s,56℃退火 30 s,72℃延伸 1 min,30個循環;72℃終延伸7 min,4℃保存。
1.3.6 瓊脂糖凝膠電泳、DNA測序和構建系統發育樹
采用2.0%的瓊脂糖凝膠對PCR擴增產物進行檢測,合格后測序。細菌和真菌分別登錄Ezbiocloud(http://www.ezbiocloud.net/)和 NCBI(https://www.ncbi.nlm.nih.gov/)數據庫進行已知序列的同源性比對。
1.4 數據處理與分析
利用MEGA5.05軟件進行分析,采用鄰近法(Neighbor-Joining)構建系統發育樹[16]。
2 結果與討論
2.1 乳酸菌的基本形態
2.1.1 乳酸菌的分離純化與鑒定
根據革蘭氏染色、過氧化氫酶試驗,共得到乳酸菌80株,再依據菌株在MRS固體上的基本形態及顏色共挑選出28株菌進行測序鑒定。經過富集培養得到一定量的菌株并提其DNA,以其模板進行16S rDNA擴增,經電泳檢測,結果見圖1。
由圖1可知,條帶清晰明亮,片段大小約1500 bp,符合測序要求。進而將PCR產物測序結果與Ezbiocloud進行同源性比對,并構建系統發育樹,見圖2。
圖1 部分乳酸菌16S rDNA的PCR擴增產物條帶Fig.1 Amplified products of 16S rDNA sequences of several lactic acid bacteria strain
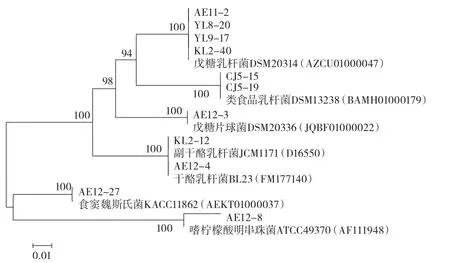
圖2 基于16S rDNA的乳酸菌系統發育樹Fig.2 Phylogenetic tree of the lactobacillus strains according to 16S rDNA sequences
結合圖2可知,處于同一分支上的測序菌株與比對菌株的置信度極高,因此,可以得到測序的23株菌中,乳酸菌有4個屬7個種,分別為戊糖乳桿菌13株、副干酪乳桿菌3株、戊糖片球菌2株、類食品乳桿菌2株、干酪乳桿菌1株、食竇魏斯氏菌1株及嗜檸檬酸明串珠菌1株。數量上,戊糖乳桿菌最多,其次是副干酪乳桿菌。根據文獻的報道,酸面團中乳酸菌主要是以植物乳桿菌、舊金山乳桿菌及短乳桿菌為優勢菌種[18-19]。然而關于新疆馕面團的研究論文中僅有朱曉瑩等[20]研究了馕中的乳酸菌,從其分離的菌株來看,并未發現上述3種菌中的任一種。而本次試驗結果也未出現,可以斷定新疆馕中的優勢乳酸菌與其他地方的存在差異。
不同地區樣品中乳酸菌的分布情況見表2。
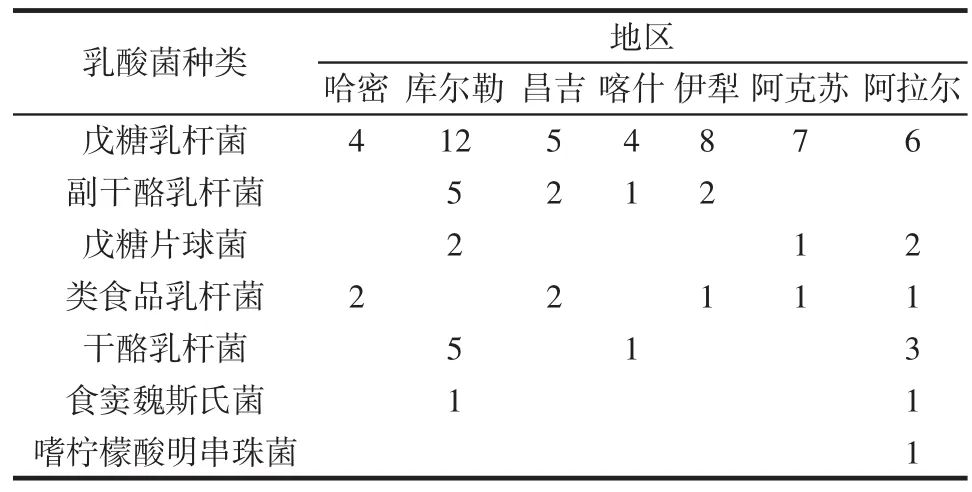
表2 不同地區樣品中乳酸菌的分布情況Table 2 Distribution of lactobacillus from different samples in different regions
由表2可知,菌種分布最多的是庫爾勒及阿拉爾,戊糖乳桿菌廣泛存在于7個地區,嗜檸檬酸明串珠菌只存在于阿拉爾地區,是該地區的特有菌株。
2.1.2 非乳酸菌的分離純化及鑒定
在乳酸菌的篩選過程中獲得了17株非乳酸菌細菌,根據菌株的顏色、形態和狀態進一步區分挑選出不相同的菌株5株,并提取其DNA,經過PCR擴增、電泳后測序,將其結果構建系統發育樹,見圖3。
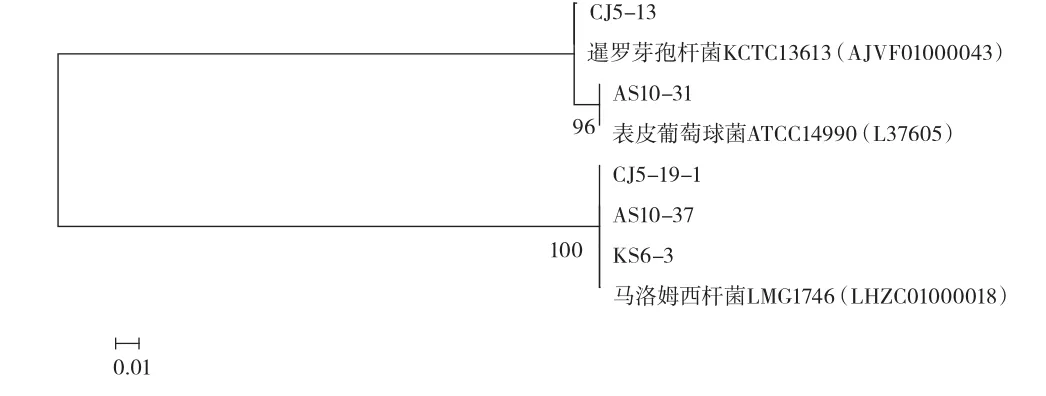
圖3 非乳酸菌16S rDNA序列的系統發育樹Fig.3 Phylogenetic tree of the non-lactobacillus strains according to 16S rDNA sequences
在17株非乳酸菌中,馬洛姆西桿菌僅伊犁的樣品未發現,暹羅芽孢桿菌發現于庫爾勒和昌吉地區,表皮葡萄球菌在庫爾勒、阿克蘇及阿拉爾樣品中都存在。具體數量見表3。
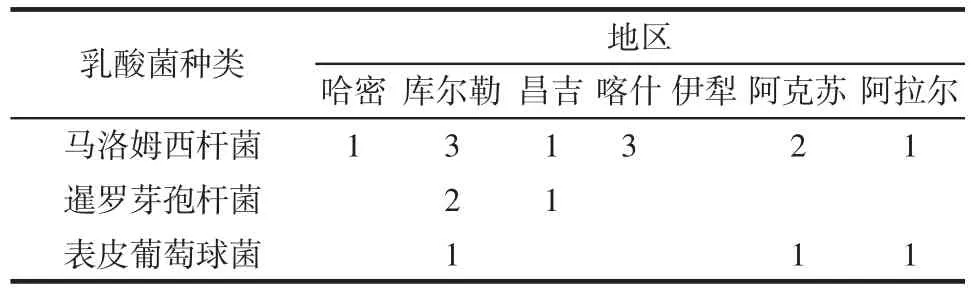
表3 非乳酸菌在各地區的分布Table 3 Distribution of non-lactobacillus stains in various regions
由圖3可知,5株非乳酸菌包含3種菌,分別為(醋酸菌屬)馬洛姆西桿菌3株、表皮葡萄球菌1株、暹羅芽孢桿菌1株。馬洛姆西桿菌屬于醋酸菌的一種,能代謝產生乙醇、二氧化碳及醋酸等物質,代謝物能增加面食的風味同時改善面團的質構,而醋酸可以降低面團的pH值從而抑制腐敗微生物的生長繁殖。暹羅芽孢桿菌尚未了解其特性。表皮葡萄球菌多數為非致病菌,少數致病。這表明了傳統酸面團中可能存在一些有害細菌。
2.2 酵母菌的基本形態
2.2.1 利用WL營養瓊脂培養基對酵母菌的基本分類
通過在WL瓊脂上菌株菌落的形態顏色觀察,結合文獻報道[21-23]比對分析,結果見表4、圖4。不同酵母菌在各地區的分布見圖5。

表4 酵母菌在WL營養瓊脂培養基上的形態描述Table 4 Description of morphology of the yeasts on WL media
由表 4、圖 4 可知,其中類型 5、6、7、9、16、17、18、19可能為釀酒酵母,類型2、12可能為畢赤酵母,類型15可能為紅酵母,其他類型則未提及。將12份樣品經過篩選純化后,共得到229株菌,利用酵母菌在WL培養基上呈現不同顏色進行分類,從而得到19種類型。由圖5可知,其中類型5、7、17、18廣泛分布于7個地區,類型15只存在于庫爾勒地區,庫爾勒地區酵母菌菌落形態分布最多,有15種,喀什地區菌落形態最少分布最少,只有6種。
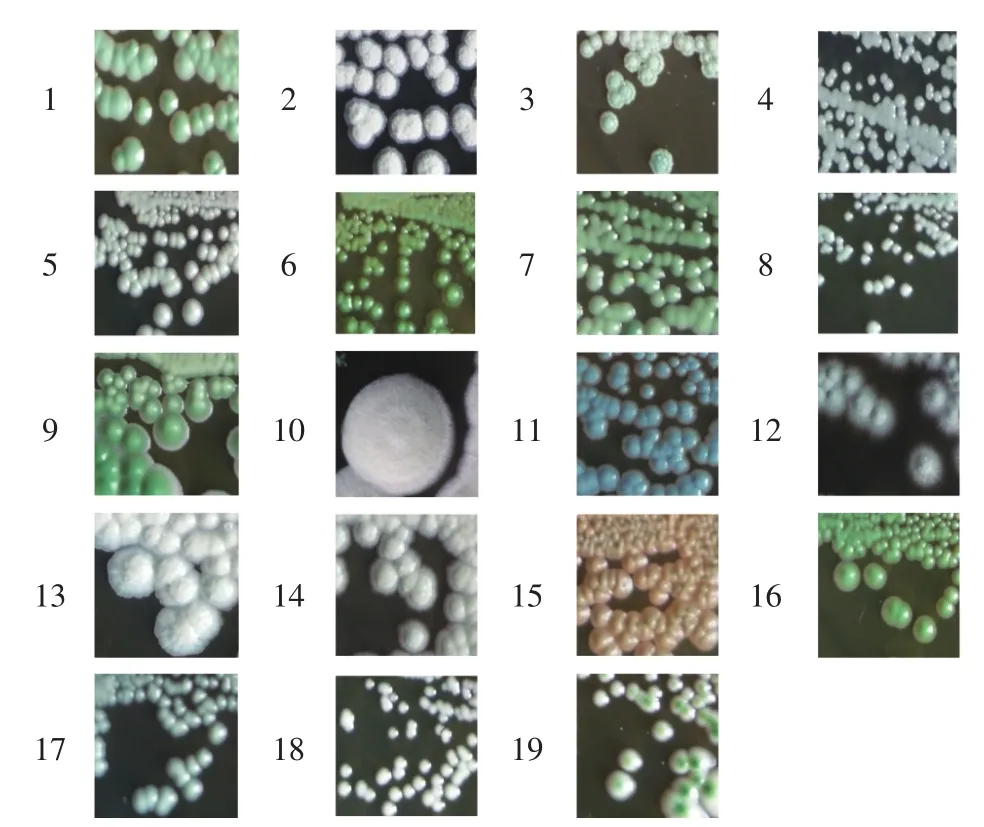
圖4 不同菌株在WL營養瓊脂培養基上的形態Fig.4 The morphology of the different strains on WL nutrient agar media
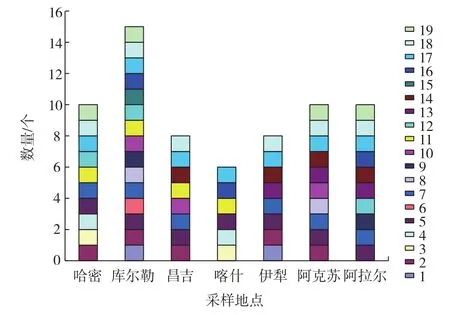
圖5 不同類型酵母菌在各地區的分布Fig.5 Distribution of different morphology of the yeasts in various regions
2.2.2 酵母菌26S rDNA D1/D2區分析
根據菌落的形狀、顏色、狀態選取不同的代表菌株33株提取其DNA,并進行26S rDNA的D1/D2PCR擴增,經電泳,條帶大約在600 bp,見圖6,符合測序要求,檢測后,將結果與NCBI進行同源性比對分析并構建系統發育樹,見圖7。

圖6 部分酵母菌26S rDNA D1/D2 PCR產物Fig.6 Amplified products of the D1/D2 sequences of several yeast strains
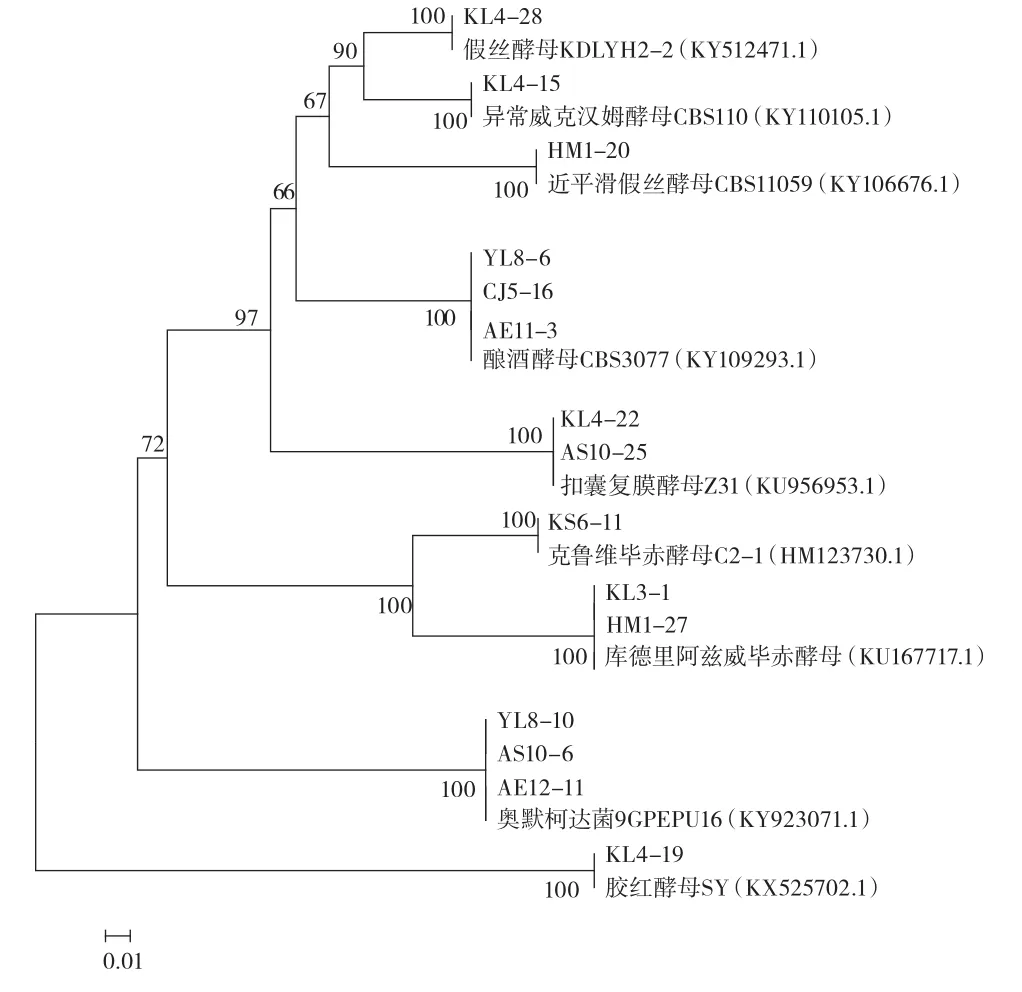
圖7 酵母菌基于26S rDNA的系統發育樹Fig.7 Phylogenetic tree of the yeast strains according to 26S rDNA
根據測序結果得出: 類型 5、6、7、8、9、16、17、18、19鑒定為釀酒酵母;類型1鑒定為假絲酵母;類型2鑒定為異常威克漢姆酵母;類型3、4鑒定為近平滑假絲酵母菌;類型10鑒定扣囊復膜酵母;類型11鑒定為克魯維畢赤酵母;類型12鑒定為庫德里阿茲威畢赤酵母;類型13、14鑒定為奧默柯達菌;類型15鑒定為膠紅酵母。鑒定的33株菌中,釀酒酵母18株,假絲酵母1株,異常威克漢姆酵母1株,近平滑假絲酵母菌7株及膠紅酵母6株;乳酸菌80株,共鑒定近平滑假絲酵母菌2株,扣囊復膜酵母2株,克魯維畢赤酵母1株,庫德里阿茲威畢赤酵母3株,奧默柯達菌4株,膠紅酵母1株,共7個屬9個種。
2.2.3 各地區酵母菌分布情況
酵母菌在各地區總的分布見表5。
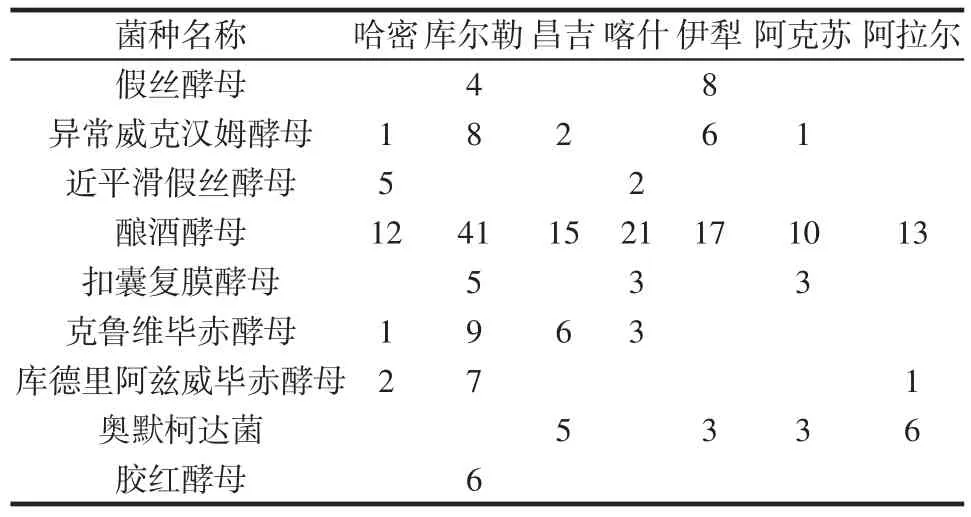
表5 酵母菌在各地區總的分布Table 5 Distribution of the yeasts in various regions
從表5可以看出,釀酒酵母是分布最廣,數量最多的菌種,并且在所有地區均有發現,這與以往的研究一致。異常威克漢姆酵母也分布較廣,研究發現,其能產生抑菌物質從而延長面食的貨架期[24],這與新疆馕有超長的食用周期可能存在一定的關系。膠紅酵母僅存在于庫爾勒地區,說明是該地區的特有菌株。
3 結論
通過對新疆7個地區12個樣品的研究發現,不同地區的酸面團菌種區別較大。主要對其中的酵母和乳酸菌進行檢測,結論如下:乳酸菌發現7種,分別為戊糖乳桿菌、副干酪乳桿菌、戊糖片球菌、類食品乳桿菌、干酪乳桿菌、食竇魏斯氏菌及嗜檸檬酸明串珠菌,從菌種占的數量和地區分布來看,戊糖乳桿菌和類食品乳桿菌為優勢菌;酵母菌發現9種,分別為假絲酵母、異常威克漢姆酵母、近平滑假絲酵母、釀酒酵母、扣囊復膜酵母、克魯維畢赤酵母、庫德里阿茲威畢赤酵母、奧默柯達菌及膠紅酵母,釀酒酵母為優勢菌;非乳酸菌3株,分別為馬洛姆西桿菌、表皮葡萄球菌及暹羅芽孢桿菌。由此可知,傳統酸面團菌種組成復雜多樣,正是其復雜的微生物菌相致使傳統發酵面食擁有獨特的風味和口感。本次研究也說明了新疆不同地區不同酸面團樣品的微生物存在明顯差異,這為以后的深入研究奠定基礎。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
